


Abstract
We previously found that 67Ga was more potent and more specific in single-cell kill than other Auger electron emitters and β-particle emitters, using an anti-CD74 antibody (Ab) (major histocompatibility complex [MHC] class II invariant chain). Because anti-CD74 Abs follow an unusual processing pathway, with rapid delivery in very large amounts to lysosomes, it was important to determine if similar results would be obtained with other, more typical Abs. Methods: Target cells were Raji B-lymphoma cells, and the Abs tested were antimature MHC class II antigen (lacking the invariant chain) and anti-CD20, both of which react with high-density antigens. Labeling was with 125I or 131I, by conventional iodination; with 111In using the chelator benzyl-diethylenetriaminepentaacetic acid; or with 67Ga using the chelator 1,4,7-triazacyclononane-1,4,7-triacetic acid. Abs were incubated with the cells for 2 d, uptake of radioactivity was assayed at various times, and toxicity was assayed primarily by a clonogenic assay. The fraction surviving was plotted versus cumulative disintegrations per cell to determine relative potency. Results: The ranking of the radionuclides for potency was 131I > 67Ga > 125I > 111In. 67Ga was approximately 2- to 3-fold more potent than 111In. This was very similar to previous results with anti-CD74. Dosimetry calculations were generally consistent with the level of toxicity observed. In previous studies of nonspecific toxicity, the order of ranking was the same. Conclusion: The subcellular location of the bound Ab (whether on the cell surface or the cytoplasm) does not appear to be an important variable in the choice of radionuclide for single-cell kill. 67Ga is a promising radionuclide for killing micrometastases, for high-density target antigens, but methods for achieving higher specific activity are required to fully exploit this approach. Each of the 4 radionuclides tested has certain advantages, and further studies are required to select the optimal radionuclide for a particular purpose.
We recently described the efficient and specific killing of tumor cells in vitro by antibodies (Abs) conjugated to radionuclides emitting electrons of varying energies (1,2). This killing was due to the radioactivity of Abs binding to each individual cell, and there was no significant role of either cross-fire from Abs bound to nearby cells or radioactivity in the medium. Killing was demonstrated only with Abs reacting with high-density antigens: The 3 Abs that were effective bound at a level of >106 molecules per cell. Thus, this therapeutic approach is currently limited to very high-density antigens, but it could be effective with lower density antigens if the approach were optimized. One obvious variable is the radionuclide used, which affects both the potency of specific toxicity and the level of nonspecific toxicity. The previous experiments used 125I, 111In, 67Ga, 99mTc, 131I, and 90Y. The first 4 of these emit Auger and conversion electrons, whereas the last 2 emit β-particles, which are much higher energy electrons. The relative effectiveness of these 6 radionuclides was investigated most thoroughly with Ab LL1, which reacts with CD74, the invariant chain of the major histocompatibility complex (MHC) class II antigen. Although all killed effectively and specifically, it was found, unexpectedly, that 67Ga was significantly more potent than the other Auger electron emitters, meaning that fewer decays per cell were required to obtain a particular level of cell kill. In comparison with β-particle emitters, the Auger emitters produced less nonspecific toxicity (due to unbound radioactivity, linked to nonreactive Abs, in the medium). Therefore, the specificity index for 67Ga was substantially higher than for any of the other radionuclides tested. The potency of 67Ga is likely to be due to its abundant Auger electrons of 5–10 keV. In contrast, the other Auger electron emitters have primarily lower energy electrons of 1–5 keV. Although these experiments were performed in vitro, there is considerable evidence that similar considerations would be important in vivo. Using a human B-lymphoma xenograft model in SCID mice, effective therapy of disseminated tumor was demonstrated with 111In and 67Ga conjugates of LL1, whereas 90Y conjugates were ineffective (3). The lack of activity of 90Y can be attributed to the >10-fold lower maximum tolerated dose, relative to the Auger electron emitters, which is due to greater nonspecific toxicity, the cause of which is the range of the β-particles.
It is uncertain, however, to what extent the results obtained with LL1 are applicable to other Abs, because of the unusual processing pathway taken by this Ab. LL1 is present at a relatively low density on the cell surface but is continuously internalized and replaced by newly synthesized molecules, so it is delivered to lysosomes in very large amounts, approximately 107 Ab molecules per cell per day (4). It is rapidly degraded in lysosomes, but if residualizing labels are used (as was done in the previous studies) a large amount of activity remains trapped within lysosomes. It seems reasonable to suggest that other Abs, following more typical processing pathways, may display differences in the relative potency of various radionuclides. Low-energy electrons, including Auger and conversion electrons, would be expected to be more potent from an intracellular location, especially if close to the nucleus, than from the cell surface. It was therefore important to perform similar experiments with other, more typical Abs reacting with the cell surface, and this was the purpose of the present investigation. The 2 Abs used, L243 and 1F5, were previously found to efficiently kill single cells (2). L243 reacts with a monomorphic determinant of MHC class II αβ and binds in large amounts to the cell surface. Although there is some slow catabolism of the bound Ab, primarily due to the slow turnover of membrane constituents, the large majority of the bound Ab remains on the cell surface for many days after Ab binding (2). 1F5 reacts with CD20 and has generally been considered to be noninternalizing. However, we recently demonstrated that it is gradually transported in large amounts to the juxtanuclear endocytic recycling compartment, which is a noncatabolic pathway (5). For this reason, cellular uptake over 1–2 d is much higher than the number of sites per cell and reaches approximately 2 × 106 Ab molecules per cell for Raji B-lymphoma cells. Because this uptake is distinct from the lysosomal uptake of LL1, and because a considerable fraction of the total bound Ab remains on the cell surface, it was of interest to also include 1F5 in this study. Abs to both CD20 and MHC class II, radiolabeled with β-particle emitters, are currently in clinical use for the therapy of B-cell lymphoma (6–8), so the results described are of immediate clinical relevance.
MATERIALS AND METHODS
Abs and Radiolabeling
The hybridomas producing Ab L243, anti-MHC class II, and 1F5, anti-CD20, were obtained from the American Type Culture Collection (Rockville, MD). The 1F5 cells were recloned as described (2). Both are IgG2as. The Abs were produced and purified as described (2). Radiolabeling methods have been described in detail (1). Briefly, 125I and 131I were conjugated using a conventional chloramine-T procedure. The radioiodine was from Perkin-Elmer (Boston, MA), and the specific activities were 370–740 MBq/mg (10–20 mCi/mg). Labeling with 111In used the chelator benzyl-diethylenetriaminepentaacetic acid (benzyl-DTPA). The 111In was from ISO-TEX (Friendswood, TX), and the specific activities were 370–1,850 MBq/mg (10–50 mCi/mg). Labeling with 67Ga used the chelator 1,4,7-triazacyclononane-1,4,7-triacetic acid (C-NOTA). The 67Ga was from MDS Nordion (Vancouver, Canada), and the specific activity was approximately 370 MBq/mg (10 mCi/mg). After labeling, all Abs were purified over a PD-10 gel-filtration column equilibrated with tissue culture medium. This was done partially for buffer exchange because removal of unbound radionuclide was not generally required for the radiometals. All labeled Abs were analyzed either by gel-filtration high-performance liquid chromatography on a Bio-Sil SEC-250 column (BIO-RAD, Hercules, CA) or by instant thin-layer chromatography on silica gel strips (Gelman Sciences, Ann Arbor, MI), or both, and >95% of the counts were associated with Ab.
Immunoreactivity was assayed with representative labels using a cell-binding assay. Binding under conditions of antigen excess (using a large number of cells, serially diluted) demonstrated the maximum bindable counts per minute (cpm). Control tubes had excess unlabeled Ab added to block specific binding and, therefore, to indicate the level of nonspecific binding: Specific binding was calculated by subtraction. Raji cells were used for L243, and RL cells were used for 1F5. For both Abs, the DTPA conjugates were prepared with an earlier batch of purified Ab that was different from the batch used for the other labels, accounting for some difference in immunoreactivity. The percentage specifically bound was 51.5% for 131I-1F5, 35.7% for 125I-1F5, 44.3% for 67Ga-1F5, and 74.8% for 111In-1F5. For L243, the values were 21.6% for 131I-L243, 19.0% for 125I-L243, 24.9% for 67Ga-L243, and 56.3% for 111In-L243. The batch of L243 used for 125I, 131I, and 67Ga labeling was prepared with ascites fluid that had a low Ab concentration, which probably accounts for the low immunoreactivity. The potential impact of this variation in immunoreactivity is discussed below.
Cell Lines
The cell lines used were the B-cell lymphomas Raji and RL. The origin of these lines and the culture conditions have been described (2). Cell lines were tested routinely for mycoplasma contamination using the Mycotect assay (Life Technologies, Grand Island, NY) and were negative.
Cytoxicity Assays
The assay using cell counts, until cells multiplied 16-fold, was described previously in detail, as was the cloning assay (1,9). Briefly, in both assays cells were incubated for 2 d with serial dilutions of Ab in 24-well plates, in a volume of 1.5 mL. In some experiments, the Ab concentrations used were selected from preliminary studies and were intended to include the range of cell killing from 99% to 99.9%. After 2 d, the cells were handled in various ways. One well at each Ab concentration, as well as a control well, was used for the clonogenic assay, as described below. The remaining wells were diluted into 11.5–20 mL of medium, depending on the particular assay, and maintained for an additional 3–19 d. At 4 h, 24 h, and daily intervals thereafter, out to 5–6 d, aliquots of cells were washed and counted for viable cells and for radioactivity. The cpm per viable cell was calculated. Although cells counts were obtained at every time point, most of the data is expressed in terms of cpm per initial cell number, rather than the actual cell number, for reasons that are discussed under Results. Other aliquots were used for cell counts for up to 21 total days. In the cloning assay, 8 serial dilutions of cells were plated in 96-well plates, with feeder cells of mitomycin C-treated Raji cells. Each dilution of cells was plated in 48 wells. Cells with growing clones were scored 14 d after plating, and the cell concentration was calculated as described (1). The cloning efficiencies were always >50%.
Dosimetry
The cpm was converted to disintegrations per minute (dpm) from the γ-counter efficiencies (1). From a graph of dpm per cell versus time, the total cumulative disintegrations per cell were determined from the area under the curve. The logarithm of the fraction surviving, at each Ab concentration (determined by the cloning assay), was plotted versus the cumulative disintegrations per cell over 5–6 d. A straight line was fitted to the points by linear regression, and the slope m of the line was used to calculate D0 (the disintegrations required for 63% kill in the linear part of the curve), as −0.4318/m. The estimated cGy dose to the nucleus was calculated from the disintegrations using the published S factors for radioactivity in either the cytoplasm or on the cell surface (10), as indicated, for a cell the size of Raji (RC = 8 μm; RN = 6 μm). Other details have been described (1,9).
RESULTS
Cytotoxicity of Raji Cells with 67Ga Conjugates of L243 and 1F5
Figure 1 shows the killing of Raji cells with these conjugates. Killing with conjugates of 125I, 131I, and 111In, in similar experiments, has been described elsewhere (2). Results with a nonreactive control Ab, determined previously (1), are also shown, for comparison. Although the negative control Ab was an IgG1, and L243 and 1F5 are IgG2as, this difference is not significant, because we have shown that there is virtually no nonspecific uptake of either IgG1 or IgG2a Abs by Raji cells (M.J. Mattes, unpublished data, 2000). Figure 1 cannot be used for a precise quantitative comparison of the Abs because it does not consider variations in specific activity and immunoreactivity, but it does provide a crude estimate of relative efficacy of the Abs. Both Abs killed the cells efficiently and specifically, but 1F5 was substantially less active than L243, with a maximum kill of slightly <100% (fraction surviving, 4.69 × 10−5). The difference between the Abs is not due to differences in immunoreactivity because the immunoreactivity of 67Ga-1F5 was higher than that of 67Ga-L243, The shape of the 1F5 curve, with a bend upward at approximately 0.407 MBq/mL (11 μCi/mL), is discussed below.

Cytotoxicity of Raji cells with 67Ga conjugates of L243 (▪) and 1F5 (•) compared with negative control Ab (□). Level of toxicity was calculated from growth curves. Highest concentration of L243, 1.11 MBq/mL (30 μCi/mL), killed 100% of cells; because fraction surviving of 0 cannot be plotted on logarithmic scale, it was plotted as 10−6, a reasonable estimate considering that each sample contained 5 × 105 cells. Results shown are representative of at least 2 experiments with each Ab, each done in duplicate.
Comparison of 4 Radionuclides in Cytotoxicity of Raji Cells
Four radionuclides were tested for their potency in killing Raji cells—namely, 131I, 125I, 111In, and 67Ga. In this article, the word “potency” is used only to refer to the relative effectiveness of various radionuclides and is quantified as the number of disintegrations per cell required for a particular level of cell kill. Because catabolism of these Abs after binding to the cell surface is slow (although not insignificant) (11,12), it was not necessary to use a residualizing form of iodine to deliver sufficient radioactivity, as was required in the previous studies with LL1.
The cloning assay was used to determine the level of killing, rather than the assay based on growth curves, because this was found to provide a more precise measure of cell kill. However, the results of the 2 assays were consistent with each other. Accordingly, the preliminary experiments, to select the optimal Ab concentrations, were done with the assay based on growth curves, because this is much simpler to perform. In the cloning assay, three 2-fold dilutions of each Ab were tested, with the concentrations selected to include a range of fraction surviving from 0.01 to 0.001. The highest concentration tested was 0.148 MBq/mL (4 μCi/mL) for 131I-L243, 0.444 MBq/mL (12 μCi/mL) for 125I-L243, 0.592 MBq/mL (16 μCi/mL) for 111In-L243, 0.37 MBq/mL (10 μCi/mL) for 67Ga-L243, 0.74 MBq/mL (20 μCi/mL) for 131I-1F5, 1.85 MBq/mL (50 μCi/mL) for 125I-1F5, 4.44 MBq/mL (120 μCi/mL) for 111In-1F5, and 2.59 MBq/mL (70 μCi/mL) for 67Ga-1F5. These concentrations are given because they provide some indication of the efficacy of the 2 Abs: Note that much higher concentrations of 1F5 than of L243 were required to obtain comparable levels of cell kill. The uptake per cell at various time points, in representative experiments, is shown in Figure 2. For improved clarity of this comparison, we have plotted the values obtained for a single Ab concentration, using the concentration at which the fraction surviving was closest to 0.0032 (2.5 logarithms of kill). These values provide a rough indication of the relative potency of the radiolabels, but they cannot be used for a precise comparison, because there was some variation in the exact level of cell kill, the specific activities of the Abs, and in the immunoreactivities of the Abs. Still, it can be inferred that radionuclides with relatively high uptake per cell are less potent than the others, because more activity was required to obtain the same level of kill. For both Abs, the order of potency was 131I > 67Ga > 125I > 111In.
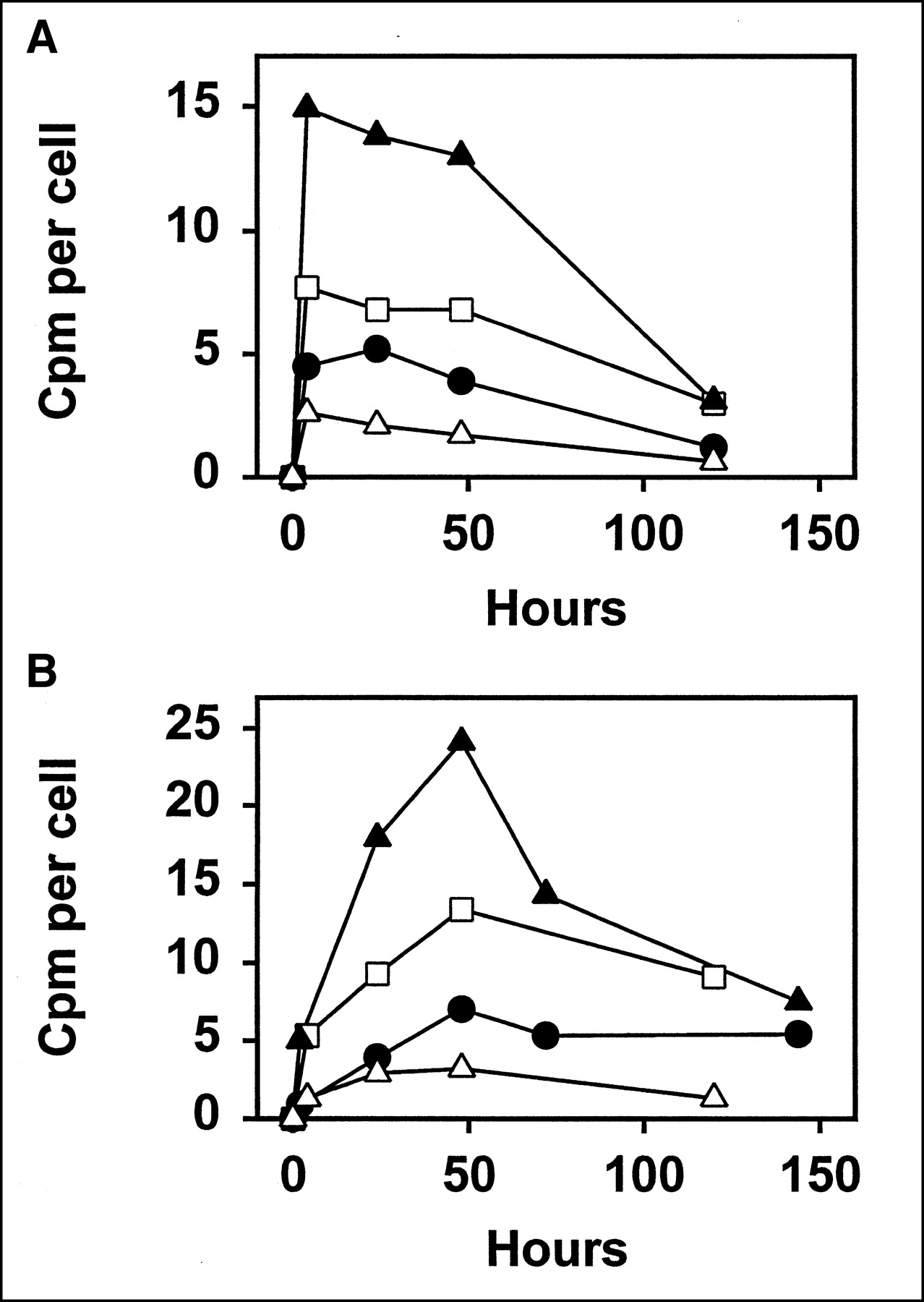
Uptake of radioactivity by Raji cells incubated with Ab L243 (A) or 1F5 (B). Data shown are for Ab concentration at which cytotoxicity was closest to 2.5 logarithms (fraction surviving, 0.00316, as determined by cloning assay), and was selected from 2 or 3 experiments with each radiolabel, each of which had consistent results and was done in duplicate. Radionuclides used were 125I (□), 111In (▴), 67Ga (•), and 131I (▵). Note that more potent radionuclides will have lower uptake in this comparison (because they produce same level of kill with fewer disintegrations).
Some differences between the different Abs and radionuclides, as shown in Figure 2, should be noted. The kinetics of uptake was clearly different for 1F5 than for L243, in that L243 binding reached its peak in only 2–4 h and did not increase significantly after that time. In contrast, 1F5 binding was relatively low at 2–4 h and required 1–2 d to reach its peak. This reflects the different processing pathways followed by the Abs, as described above. The difference in half-life of the radionuclides also may affect the shape of the binding curve. With the radionuclides having shorter half-lives of approximately 3 d (111In and 67Ga), the cpm per cell is expected to decreased markedly after day 2 (when the cells were diluted 10-fold with medium, which will greatly reduce subsequent binding) as a result of radioactive decay. In contrast, cpm from 125I are expected to remain at a high level for the duration of the experiments, 5–6 d, because of the 60-d half-life. However, this effect is not prominent in Figure 2, probably because it is obscured by other factors.
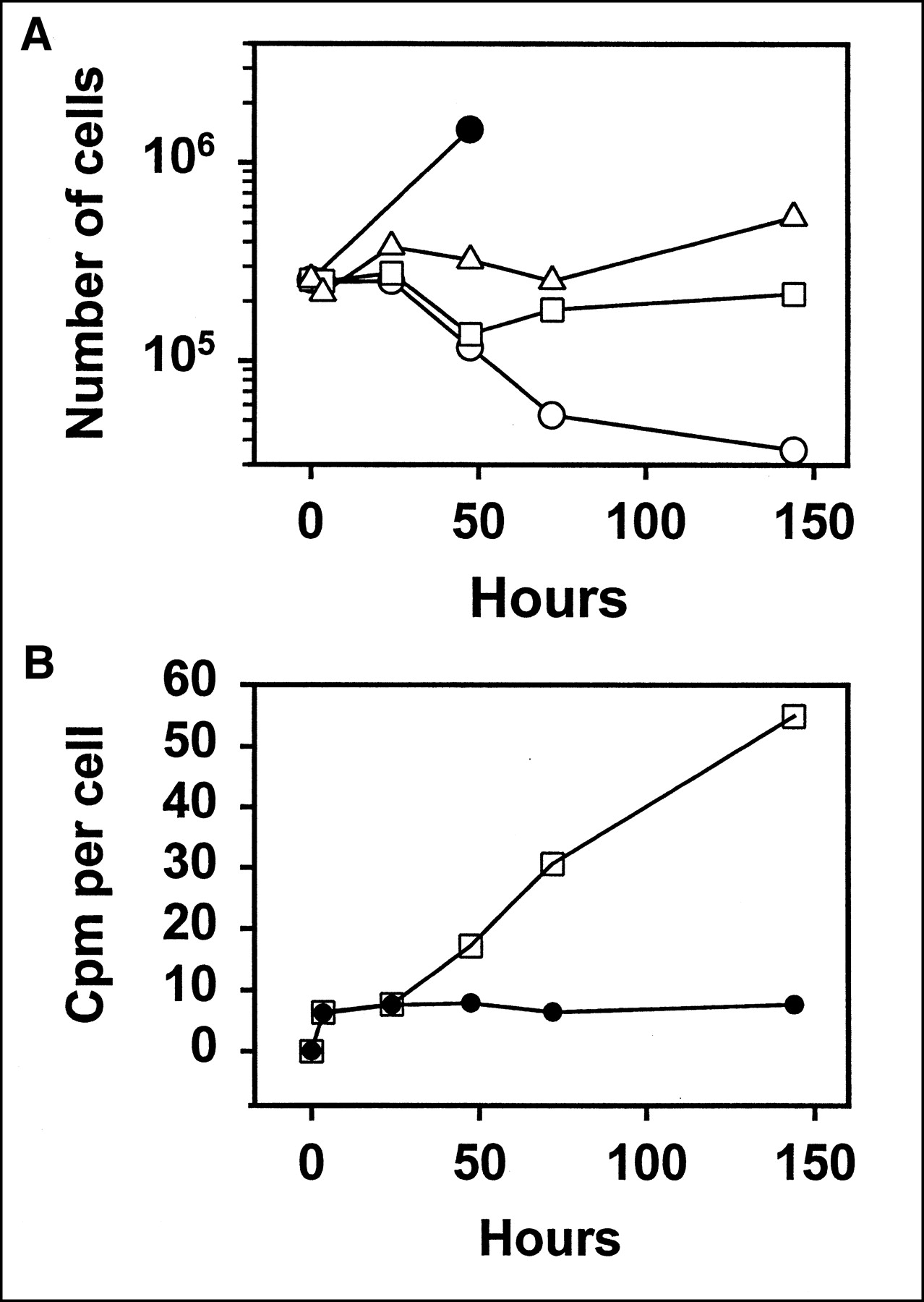
The method of calculating cpm per cell must be discussed. This can be expressed using either the actual viable cell number at each time point or the initial viable cell number. Both were determined in every experiment. However, use of the initial cell number is more useful, because the cells begin to die as early as day 2, and this results in misleading high values of the cpm per viable cell. This occurs because the dead cells retain their radioactivity and are pelleted together with the viable cells. This problem has been discussed in more detail elsewhere (1). Because the cells generally divide very little over the 2 d of Ab exposure, as a result of the quite high dose of radiation, the cpm per initial cell number provides a good estimate, and is probably the best possible estimate, of the true value. To illustrate this point, Figure 3A shows growth curves in a typical experiment. This figure demonstrates that at relatively high radiation doses, at which the cell kill is 99%–99.9%, cell division very rapidly halts, and there is little or no increase over the starting cell number. Figure 3B demonstrates that calculation of the cpm per actual cell number (as opposed to the initial cell number) gives misleading high values because of cell death.

(A) Typical growth curve in toxicity assay. Raji cells were incubated with 125I-1F5 at concentrations of 1.85 MBq/mL (50 μCi/mL) (○), 0.925 MBq/mL (25 μCi/mL) (□), or 0.463 MBq/mL (12.5 μCi/mL) (▵). At relatively high levels of radiolabeled Ab used, cell division stops very rapidly. Significant cell death is first evident at 48 h. Growth rate of untreated cells is also shown (•). Fraction surviving in this experiment, calculated from cloning assay, was 1.07 × 10−4, 2.79 × 10−3, and 0.0207, respectively. (B) Difference between cpm per actual cell number (□) and cpm per initial cell number (•), in same experiment, with Ab concentration of 1.85 MBq/mL (50 μCi/mL). Due to cell death, cpm per actual cell number becomes artifactually high at times ≥48 h.
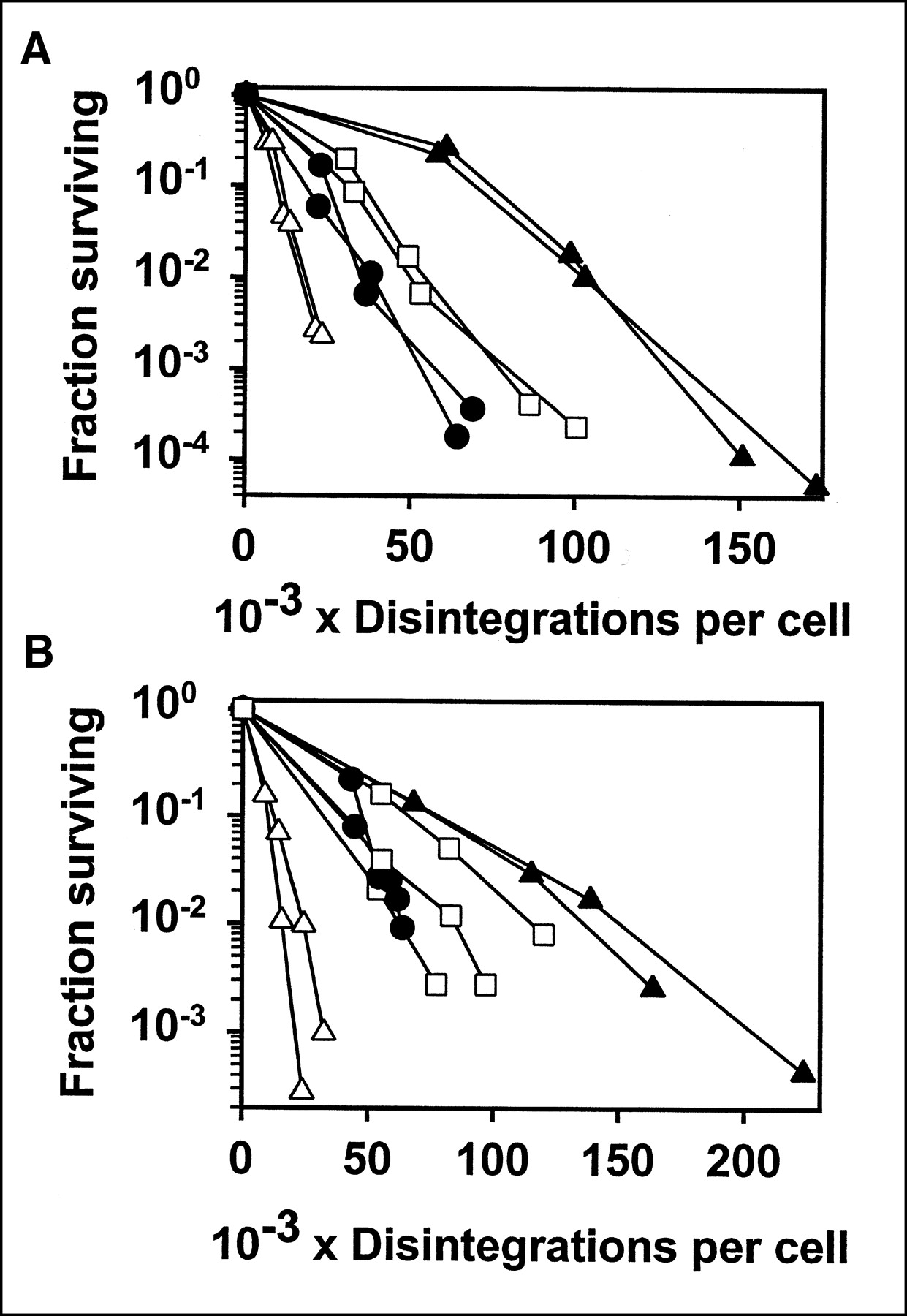
Figure 4 shows the fraction surviving versus cumulative disintegrations, for both L243 and 1F5 labeled with 4 radionuclides. 67Ga was substantially more potent than 111In and was slightly more potent than 125I. The β-particle emitter, 131I, was more potent than 67Ga and, thus, was the most potent of the radionuclides tested. The results were similar for both of the Abs tested. Although the difference between the radionuclides is shown clearly in Figure 4, we have also analyzed the data in other ways. From the straight line fitted to the data points shown, the D0 was calculated. This is the dose, in cumulative disintegrations per cell, required for 63% kill in the linear part of the dose-response curve. These values are presented in Table 1. We also include the dose required for 2.5 logarithms of cell kill (fraction surviving = 0.00316) because this range was included in virtually all of the experiments. This value was calculated from the 2 data points on either side, so it might be considered more reliable than the values calculated from the slope of the fitted line. In some cases with 1F5, this level of killing was not quite reached (as shown in Fig. 4), and in that case the dose was calculated from the 2 points with highest killing, by extrapolation. In fact, the 3 types of calculations are very consistent with each other. For both of the Abs tested, the relative potency of the radionuclides was 131I > 67Ga > 125I > 111In. The differences between radionuclides in the dose required for 2.5 logarithms of cell kill, for both L243 and 1F5, were all statistically significant, by the Student t test (P < 0.05), except for the comparison between 125I-1F5 and 67Ga-1F5, for which there was not a significant difference, and for the comparison between 125I-1F5 and 111In-1F5, for which P < 0.1.
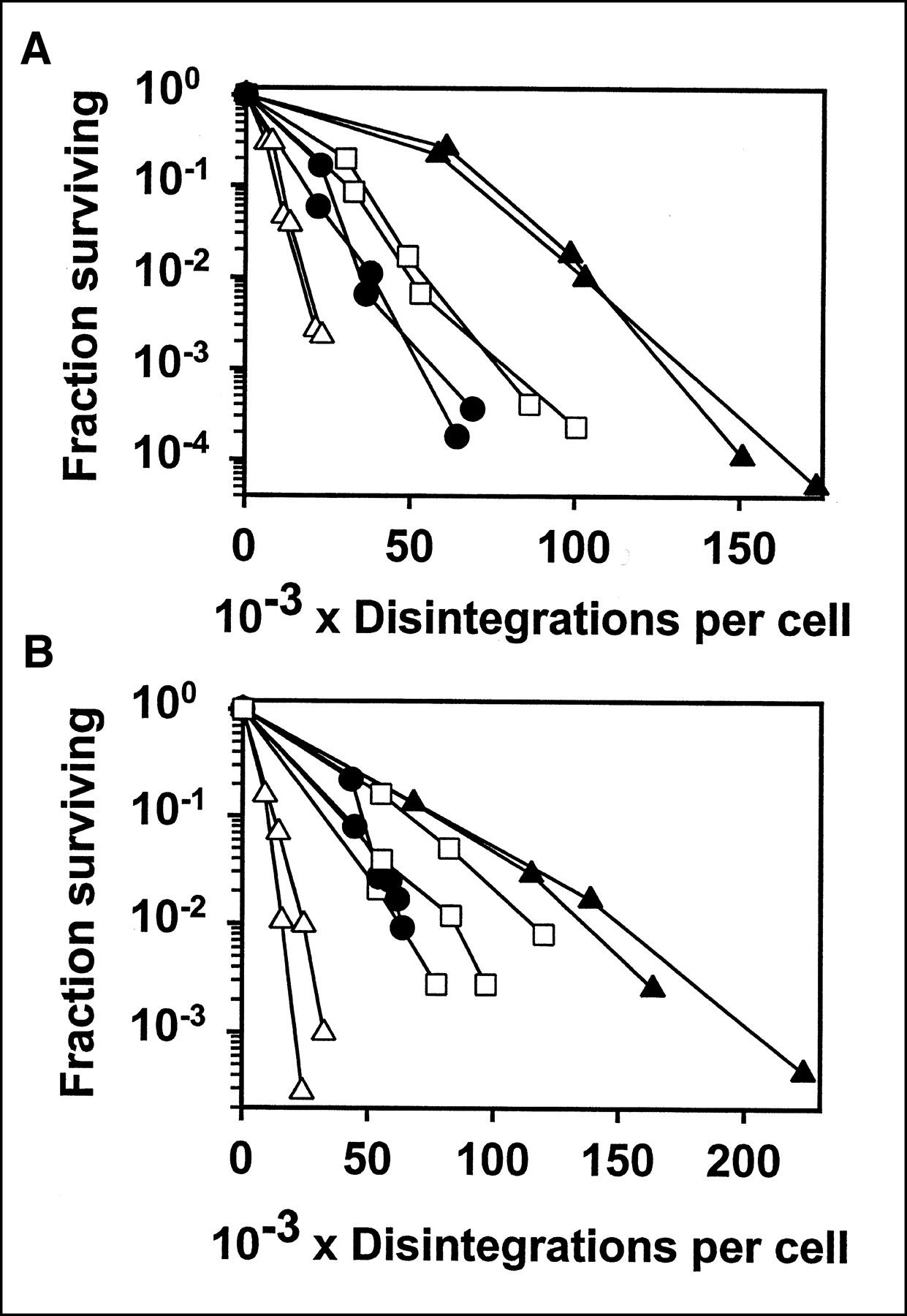
Cytotoxicity of Raji cells with Abs conjugated to 125I (□), 111In (▴), 67Ga (•), or 131I (▵). Results are shown for L243 (A) and 1F5 (B). Fraction surviving, determined by cloning assay, is plotted vs. cumulative disintegrations per cell over 5–6 d. Results of 2 or 3 separate experiments for each radiolabeled Ab are shown, with cpm determined in duplicate.
Comparison of Radionuclides, Conjugated to Abs, in Killing of Raji Cells
Although this work is primarily focused on the relative potency of the various radionuclides, per disintegration, another factor that is important in practice is the Ab concentration required to achieve the desired level of uptake. As indicated earlier (Results), this concentration, in MBq/mL (μCi/mL), was much higher for 1F5 than for L243. This factor depends on the number of antigenic sites per cell (which is much higher for L243 than for 1F5) and is also affected by the antigen accessibility, Ab avidity, and other factors. As an indication of this factor, Table 1 also presents that Ab concentration, in MBq/mL (μCi/mL), at which the cell kill was closest to 2.5 logarithms (fraction surviving, 0.00316), in the experiments that have been performed. As shown, these values were approximately 8-fold higher for 1F5 than for L243. This implies that 8-fold higher Ab concentrations are required to obtain similar levels of Ab uptake.
Dosimetry
Figure 5 shows the fraction surviving as a function of the calculated cGy dose delivered per cell. The purpose of this calculation was to determine if the calculated radiation dose is consistent with the effects observed and whether this comparison differs for the various radionuclides. For L243, because most of the Ab is on the cell surface, at all times, it seems most appropriate to select the cell surface S value, even though some fraction of the radioactivity is intracellular. For 1F5, the choice is not clear, because most of the Ab appears to localize to an intracellular site, but a considerable fraction remains on the cell surface. Therefore, we have performed the calculations in both ways, as shown in Figures 5B and 5C, respectively. The most accurate estimate would thus be some intermediate value, but many of the conclusions are the same in either case. Three observations can be made: (a) The calculated doses for 125I and 111In are considerably higher than those for 67Ga and 131I. Similar results were obtained previously for Ab LL1 (1). This suggests that the effects of 125I and 111In are overestimated by the dose calculation (or that the effects of 131I and 67Ga are underestimated). (b) When the dose from 67Ga is calculated for cell surface localization, for both Abs, the effect of this radionuclide is overestimated (or the effect of 131I is underestimated), as shown clearly by the fact that 67Ga and 131I switch relative positions between Figure 5A and Figure 4A. (c) The dose-response curve with external 137Cs irradiation is included in Figure 5. Although this provides an interesting comparison, we discuss below several factors that complicate the comparison, together with possible explanations for the other effects observed.

Fraction surviving vs. calculated radiation dose, for L243 and 1F5, using data from Figures 4B and 4D. Dose for L243 is calculated assuming cell surface localization (A). Because 1F5 is localized to intracellular site and cell surface, in a proportion that is not precisely known, dose is calculated for both cytoplasmic localization (B) and cell surface localization (C). Cumulative dose was calculated over 5–6 d. Radionuclides used were 125I (□), 111In (▴), 67Ga (•), and 131I (▵). Dashed line shows sensitivity of Raji cells to external 137Cs γ-irradiation as determined (9).
DISCUSSION
These results further support our previous conclusion that efficient and specific single-cell kill by radiolabeled Abs is achievable with Abs to high-density cell surface antigens. More specifically, they demonstrate the following: (a) 67Ga was considerably more potent than the other Auger electron emitters tested, 111In and 125I, at single-cell kill. The advantage over 111In was approximately 2- to 3-fold, meaning that fewer disintegrations produced the same level of cell kill. The advantage over 125I was less, approximately 1.4-fold, but still significant. (b) 131I was somewhat more potent than 67Ga. Whereas the disadvantage of β-particle emitters such as 131I, in comparison with Auger electron emitters, is greater nonspecific toxicity, 131I still has a high level of specificity, and so may potentially be more useful than Auger emitters in certain circumstances. (c) In terms of the specificity index, 125I appears to be the best of the radionuclides tested, although the difference from 67Ga and 131I is not large. This is due to the very low nonspecific toxicity of 125I. (d) The difference between radionuclides does not appear to be dependent on either the particular Ab used or the subcellular localization of that Ab, because the results presented here with L243 and 1F5 are generally very similar to the previous results with LL1. Because 2 batches of both L243 and 1F5 were used in this study, differing significantly in immunoreactivity (Materials and Methods), the possible impact of this difference should be discussed. Because, in these experiments, the uptake of radioactivity per cell was determined and was used in most of the calculations and graphs (rather than the initial concentration of radioactivity), and because, in these experiments, nonspecific toxicity is insignificant compared with specific toxicity (1), such variation in immunoreactivity is not expected to affect the results. Moreover, the 111In conjugates had higher immunoreactivity than the other 3 conjugates, for both Abs, so this cannot explain the lower potency of 111In conjugates.
Even when saturation of antigen can be readily obtained, as in in vitro experiments, specific activity is a critical factor for Abs to antigens having a moderate cell density. For example, 67Ga-1F5 killed Raji cells effectively, but high protein concentrations (10 μg/mL) were required to obtain a cell kill of approximately 4 logarithms. At up to 0.407 MBq/mL (11.1 μCi/mL), with an Ab concentration of 1.1 μg/mL, the slope of the curve was steep, but at higher concentrations it became flatter. This is because antigen on the cell surface was nearly saturated, and much higher Ab concentrations resulted in only slightly greater uptake. This interpretation was verified in the uptake experiments, in which increasing Ab concentrations resulted in only slightly greater uptake. Such a near-plateau in Ab binding did not occur with L243 at the concentrations used, because 100% kill was obtained at concentrations that were nonsaturating. The higher level of Ab uptake with L243 is due to higher antigen expression on the cell surface. The important implication is that, with a higher specific activity, 1F5 could deliver more 67Ga to the cell, and the dose-response curve would probably remain steep out to higher levels of cell kill. Assuming a linear dose-response in the semilogarithmic plot, which is usually seen in experiments of this type (1,2), we can predict that a 4-fold increase in specific activity, with an Ab concentration of 1.1 μg/mL, would result in a decrease in the fraction surviving from 0.045 to <10−6. The advantage of 67Ga can be exploited only if labeling methods are improved so that higher specific activities can be attained, which seems feasible. Currently, the 10-fold higher specific activities of 111In conjugates compared with 67Ga conjugates more than compensate for the greater potency of 67Ga. There are 2 major factors involved in the production of 67Ga conjugates with higher specific activity: the chelator used and the purity of the gallium.
Although this study has analyzed only the potency of the various radiolabels, it should be emphasized that selection of the optimal radionuclide for single-cell kill depends on its specificity as well as its potency. The level of nonspecific toxicity, due to the presence of the radionuclides in the medium, was investigated previously, using the same cell line and conditions (1). The mean initial concentration of a nonreactive labeled Ab required for 99% kill was 25.41 MBq/mL (686.8 μCi/mL) for 125I, 12.28 MBq/mL (332.3 μCi/mL) for 111In, 10.18 MBq/mL (275 μCi/mL) for 67Ga, and 2.65 MBq/mL (71.6 μCi/mL) for 131I. To allow a better comparison with the data presented in this study, we can calculate the cumulative disintegrations per well (containing 1.5 mL) in 2 d to be 6.46 × 1012 for 125I, 2.51 × 1012 for 111In, 2.13 × 1012 for 67Ga, and 6.22 × 1011 for 131I. Thus, 131I produced the greatest nonspecific toxicity, but it was also the most potent of the isotopes for specific cell kill, as shown in Table 1. Table 2 is an attempt to quantify the relative specificity of the 4 radionuclides tested. Because 131I was most potent in both tests, it is convenient to express the potency of the other radionuclides relative to 131I. Therefore, increased specificity, relative to 131I, is indicated by a specific toxicity ratio that is higher than the nonspecific toxicity ratio. As shown in Table 2, this advantage is greatest for 125I (2.4- to 3.5-fold), less for 67Ga (1.1- to 1.6-fold), and nonexistent for 111In (0.6- to 0.7-fold) (meaning that 111In has a slightly lower level of specificity than 131I). The advantage of 125I is due to its very low nonspecific toxicity, considerably lower than that of 111In or 67Ga, whereas its specific toxicity is only slightly less than that of 67Ga. The disadvantage of 111In is due to its very low specific toxicty. Although 90Y was not included in this study, it was included in previous experiments with LL1 (1). 90Y had the highest nonspecific toxicity, without a particularly high level of specific toxicity, and therefore had the lowest specificity index of the 5 radionuclides tested. Therefore, 90Y is not favorable for single-cell kill, and this is the reason that it was not included in the current study.
Advantage of Auger Electron Emitters in Cytotoxicity of Raji Cells Relative to 131I
The basic question, of course, is which radionuclide would be best for clinical use, in a particular setting. This question cannot be definitively answered, in part because the differences in the level of specificity between some of the radionuclides is not large and also because the level of nonspecific toxicty that will occur in humans cannot be precisely predicted from the level occurring in vitro, or even in mouse models. Furthermore, the level of nonspecific toxicity will depend to some extent on the particular Ab used. The long, 60-d half-life of 125I appears to be a disadvantage for clinical use. The fact that 131I can kill larger, heterogeneous tumor masses as well as single cells must be considered an advantage, even though the same property causes greater nonspecific toxicity. An attractive strategy for therapy is to combine 2 radionuclides, attacking microscopic tumor with Auger electrons and larger tumor masses with β-particles.
Although this study has focused on the comparison of radionuclides, it is also of interest to compare the Abs tested. From Figure 1 and Table 1, it is evident that L243 is an effective toxic agent at a much lower concentration than 1F5, whereas, on the basis of previous data (1), L243 and LL1 are similar in the concentration required for effective toxicity. This difference can be attributed to the differences in the level of Ab binding. The advantage of 1F5, however, is its greater specificity for B-cells, because it does not react with macrophage-lineage cells, as do the other 2 Abs. Although conjugates of LL1 and L243 appear to be very similar as toxic agents, the effective therapy obtained with 111In-LL1, reported previously (3), has not been matched in recent experiments with 111In-L243, used at the same specific activity (M.J. Mattes, unpublished data, 2002). The reason for this difference is not known, although it may be due to the different subclasses of the Abs (IgG1 vs. IgG2a, respectively), because IgG2as are cleared from the blood abnormally fast in SCID mice (14).
The data, together with earlier studies (1), strongly suggest that the subcellular location of the isotope has relatively little effect on its potency and that Ab internalization is not required for cytotoxicity by Auger electron emitters. This conclusion is based on the fact that the 3 Abs tested have very different subcellular localizations. Because it is frequently assumed that Auger electron emitters must be internalized to be effective toxic agents (15–18), it is useful to discuss this point. First, the published S values for radionuclides in different subcellular compartments (10) in fact show a relatively small difference between the cytoplasm and the cell surface (for radiation delivered to the nucleus): This difference is <2-fold for 125I and 111In, although it is 3.8-fold for 67Ga. Second, the assumptions necessary to perform the dose calculations tend to exaggerate the difference between the cell surface and the cytoplasm. More specifically, it was assumed that the cell membrane and the nucleus are concentric spheres. In such a model, the annulus of cytoplasm acts as an efficient shield of the nucleus for low-energy electrons emitted from the cell surface. For Raji cells the annulus is 1.55 μm. The lower energy Auger electrons of <5 keV cannot cross this barrier, and the abundant 7.4-keV electrons of 67Ga, with a range of 1.63 μm (19), will only reach the nucleus if emitted straight downward. But deviations from this model are considerable: The nucleus of Raji cells is generally indented and located very close to one side of the cell, with the other side containing most of the organelles (M.J. Mattes, unpublished data, 2001). For this reason, the fraction of decays from the cell surface that will reach the nucleus will be greatly increased, and it is probably more appropriate to use the cytoplasmic S value rather than the cell surface S value for dose calculation.
Although catabolism of L243 and 1F5 by Raji cells is quite low, it is not insignificant (11,12). For this reason, the residualizing radiolabels used, 111In and 67Ga, might be expected to have an advantage relative to the nonresidualizing iodine labels. Any iodine that is on Abs that are catabolized will be rapidly released from the cells, probably as iodotyrosine (20), whereas the residualizing labels are trapped in lysosomes, which are generally close to the nucleus. The percentage of initially bound Ab that is catabolized by Raji cells in 2 d is 21% ± 1% for L243 (11) and 36% ± 9% for 1F5 (M.J. Mattes, unpublished data, 1999; but similar to the value obtained with Ramos cells (12)). Because these values were obtained from experiments in which Ab binding was brief (1 h) followed by washing and incubation in tissue culture medium, they represent an overestimate of catabolism in a prolonged Ab incubation experiment, as used herein to determine cytotoxicity. Therefore, this effect will be small and, in any case, does not affect the comparisons between the 2 radiometals and the 2 iodine labels. If the presence of the radionuclide in lysosomes is a significant factor in toxicity, then the potency of the iodine labels should be relatively diminished. This difference might be seen by comparison with the previous LL1 results (1), because in those experiments a residualizing form of iodine was used. In fact, 131I and 125I were at least as potent on L243 and 1F5 as on LL1, suggesting that this factor is not significant.
The dosimetry calculations shown in Figure 5 should be discussed. First, it is clear that higher calculated cGy doses of 125I and 111In, in comparison with 67Ga and 131I, are required to have the same effect. We suggest that this is due to the fact that 125I and 111In have abundant Auger electrons of energies <5 keV and that the effect of these electrons may be overestimated, because few of them are close enough to the nucleus to reach it. Second, for 67Ga localized to the cell surface (Figs. 5A and 5C), the calculated dose required for a particular level of cell kill is lower than for the other Auger electron emitters and comparable with that for 131I; 67Ga appears to be relatively less effective (in terms of cGy required for a particular level of cell kill) when calculations use the cytoplasmic S value (Fig. 5B). This is probably due to the annulus effect that was described above. Third, Figure 5 includes results of external 137Cs irradiation for comparison, but the data must be interpreted cautiously because there are basic differences in the methods used. The continuous radiation exposure from the accreted radionuclides, at a rate that varies with time, should be considerably less effective than the brief 137Cs irradiation, by a factor of approximately 3, due to repair of sublethal damage (21). The relative biologic effectiveness of Auger electron emitters or β-particle emitters in the cytoplasm is not expected to be significantly greater than 1 (22,23). Taking these factors into consideration, the effect of the Ab conjugates seems generally consistent with the calculated dose.
Finally, although we are limited in practice to the use of radionuclides that are available, it should not be assumed that any of the radionuclides tested here are the best choice for single-cell kill. We previously suggested several other radionuclides, not currently available, that would be expected to be considerably more potent (2,24). As better radionuclides and better chelators become available, the goal of single-cell kill with Auger electron emitters will become increasingly feasible.
CONCLUSION
With both Abs tested, anti-CD20 and anti-MHC class II αβ, 67Ga was considerably more potent in single-cell kill of B-lymphoma cells than the 2 other Auger electron emitters tested, 111In and 125I, meaning that fewer disintegrations were required for a particular level of cell kill. The ranking of the 4 radionuclides tested was 131I > 67Ga > 125I > 111In. Results with anti-CD74, in a previous study, were very similar. Inasmuch as the subcellular distribution of these 3 Abs is very different, the data suggest that selection of the optimal radionuclide for single-cell kill is largely independent of the subcellular distribution, meaning that internalization of the Ab is not a significant factor. The ranking of these 4 radionuclides in regard to nonspecific toxicity is the same, and the radionuclides that appear to have the highest specificity indexes are 67Ga and 125I. Considering the long half-life of 125I, 67Ga appears to be a promising radionuclide for treatment of micrometastases, using antibodies to high-density antigens.
Acknowledgments
We are grateful to Phillip Andrews, Nicholas Kumburis, Thomas Jackson, and Dr. Habibe Karacay for assistance with radiolabeling. This work was supported in part by grant CA87059 from the National Institutes of Health and grant ER63191 from the Department of Energy.
Footnotes
Received Jun. 11, 2002; revision accepted Oct. 24, 2002.
For correspondence or reprints contact: M. Jules Mattes, PhD, Center for Molecular Medicine and Immunology, 520 Belleville Ave., Belleville, NJ 07109.
E-mail: mjmattes{at}gscancer.org
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Noninternalizing Monoclonal Antibodies Are Suitable Candidates for 125I Radioimmunotherapy of Small-Volume Peritoneal Carcinomatosis
- Nuclear Localizing Sequences Promote Nuclear Translocation and Enhance the Radiotoxicity of the Anti-CD33 Monoclonal Antibody HuM195 Labeled with 111In in Human Myeloid Leukemia Cells
- Trifunctional Somatostatin-Based Derivatives Designed for Targeted Radiotherapy Using Auger Electron Emitters
- In vitro cytotoxicity of carcinoma cells with 111In-labeled antibodies to HER-2
- Therapy of Small Subcutaneous B-Lymphoma Xenografts with Antibodies Conjugated to Radionuclides Emitting Low-Energy Electrons
- In vitro Toxicity of A-431 Carcinoma Cells with Antibodies to Epidermal Growth Factor Receptor and Epithelial Glycoprotein-1 Conjugated to Radionuclides Emitting Low-Energy Electrons