


Abstract
Although the incidence of de novo neuroendocrine prostate cancer (PC) is rare, recent data suggest that low expression of prostate-specific membrane antigen (PSMA) is associated with a spectrum of neuroendocrine hallmarks and androgen receptor (AR) suppression in PC. Previous clinical reports indicate that PCs with a phenotype similar to neuroendocrine tumors can be more amenable to imaging by 18F-FDG than by PSMA-targeting radioligands. In this study, we evaluated the association between neuroendocrine gene signature and 18F-FDG uptake–associated genes including glucose transporters (GLUTs) and hexokinases, with the goal of providing a genomic signature to explain the reported 18F-FDG avidity of PSMA-suppressed tumors. Methods: Data-mining approaches, cell lines, and patient-derived xenograft models were used to study the levels of 14 members of the SLC2A family (encoding GLUT proteins), 4 members of the hexokinase family (genes HK1–HK3 and GCK), and PSMA (FOLH1 gene) after AR inhibition and in correlation with neuroendocrine hallmarks. Also, we characterize a neuroendocrine-like PC (NELPC) subset among a cohort of primary and metastatic PC samples with no neuroendocrine histopathology. We measured glucose uptake in a neuroendocrine-induced in vitro model and a zebrafish model by nonradioactive imaging of glucose uptake using a fluorescent glucose bioprobe, GB2-Cy3. Results: This work demonstrated that a neuroendocrine gene signature associates with differential expression of genes encoding GLUT and hexokinase proteins. In NELPC, elevated expression of GCK (encoding glucokinase protein) and decreased expression of SLC2A12 correlated with earlier biochemical recurrence. In tumors treated with AR inhibitors, high expression of GCK and low expression of SLC2A12 correlated with neuroendocrine histopathology and PSMA gene suppression. GLUT12 suppression and upregulation of glucokinase were observed in neuroendocrine-induced PC cell lines and patient-derived xenograft models. A higher glucose uptake was confirmed in low-PSMA tumors using a GB2-Cy3 probe in a zebrafish model. Conclusion: A neuroendocrine gene signature in neuroendocrine PC and NELPC associates with a distinct transcriptional profile of GLUTs and hexokinases. PSMA suppression correlates with GLUT12 suppression and glucokinase upregulation. Alteration of 18F-FDG uptake–associated genes correlated positively with higher glucose uptake in AR- and PSMA-suppressed tumors. Zebrafish xenograft tumor models are an accurate and efficient preclinical method for monitoring nonradioactive glucose uptake.
The androgen receptor (AR) plays a central role in regulating the transcriptional events driving prostate cancer (PC) progression and development of metastatic castration-resistant PC (mCRPC) (1). AR inhibition is an effective therapeutic approach for most patients at different stages of PC. Although the incidence of de novo neuroendocrine PC (NEPC) is considered rare, several emerging forms of PC with low levels of AR are identified. The suppression and low activity of AR in these patients is largely associated with a neuroendocrine gene signature (NEGS) and resistance to AR inhibition (2,3).
AR-directed therapy of mCRPC could promote cellular plasticity and development of an AR-suppressed phenotype similar to NEPC, which manifests the histopathology of neuroendocrine disease (4). Another emerging phenotype of mCRPC is AR-null and neuroendocrine-null, classified as double-negative PC (DNPC) (3). A recent molecular subtyping of PC patients with no history of AR-directed therapies identified a neuroendocrine-positive subtype with low chromatin binding and activity of AR. These patients have been referred to as having neuroendocrine-like PC (NELPC) since it does not represent the neuroendocrine histopathology (2,5). NEPC is associated with the loss of RE1-silencing transcription factor (REST) due to alternate splicing by the RNA splicing factor serine/arginine repetitive matrix 4 (SRRM4). SRRM4 plays a crucial role in progression to NEPC under next-generation AR inhibitors, such as abiraterone and enzalutamide (6). Hence, the elevation of SRRM4 and the loss of its target REST could be specific markers of treatment-induced NEPC.
AR regulates the expression of FOLH1 gene encoding the transmembrane protein prostate-specific membrane antigen (PSMA). PSMA-targeted molecular imaging and therapy are transforming the landscape of PC management (7,8). Despite the impactful implications of PSMA, there are clinical reports that suggest that PSMA-targeted imaging does not effectively visualize NEPC tumors (8–10). Preclinical studies confirmed that the induction of lineage plasticity by AR inhibition leads to neuroendocrine transdifferentiation and suppression of PSMA (11).
Similar to glucose, 18F-FDG is actively transported into the cell by the protein family of glucose transporters (GLUTs) and is encoded by SLC2A genes, followed by phosphorylation by hexokinase (12). In some types of tumors, a positive correlation has been reported between 18F-FDG uptake and the levels of specific GLUTs and hexokinases (12–14). 18F-FDG PET is a well-accepted approach for delineation of proliferative and poorly differentiated or dedifferentiated neuroendocrine tumors (15). Nevertheless, 18F-FDG PET has been considered ineffective in assessing metastatic tumor burden and monitoring therapy response (16). Recent case reports illustrate that NEPC delineation may be more feasible by 18F-FDG than by PSMA radioligands (17,18) and Spratt et al. (19) demonstrated the utility of 18F-FDG PET for NEPC imaging. Interestingly, Thang et al. (20) screened patients with 68Ga-PSMA-11 and 18F-FDG PET, and they identified a subset of patients with low PSMA-radioligand uptake and high 18F-FDG uptake.
Development of nonradioactive glucose analogs enabling delineation of the glucose uptake of tumors has been studied using a variety of optical approaches mostly in mouse models (21). PC xenograft studies in zebrafish are coming to the forefront as a cost-effective and time-efficient model for drug screening, and the fluorescent glucose bioprobe GB2-Cy3 has been used to monitor glucose uptake in a zebrafish model (22,23). However, the feasibility of monitoring glucose uptake in PC in a zebrafish model has not been evaluated.
In this work we used data-mining approaches, cell lines, and patient-derived xenograft (PDX) models to study the expression levels of glucose uptake–associated genes, including GLUTs and hexokinases in NEPC and NELPC, to provide a genomic rationalization for the previously reported 18F-FDG avidity of PSMA-suppressed PC tumors. We also present the feasibility of nonradioactive in vivo imaging of glucose uptake using a zebrafish model.
MATERIALS AND METHODS
Cell Culture
The LNCaP cell line was purchased from ATCC and grown in RPMI-1640 in the presence of 10% fetal bovine serum. Progression to NEPC was achieved by culturing LNCaP cells in RPMI-1640 medium with 10% charcoal-stripped serum for 4 mo. Under these conditions, cell morphology gradually changed into a mixture of a neuroendocrine-like phenotype and a nonneuroendocrine phenotype. After 4 mo, a subclone with a neuroendocrine-like phenotype was isolated (LNCaP-NEPC). LNCaP cells, maintained in RPMI-1640 plus fetal bovine serum, were used as a control and are referred to as LNCaP-adenocarcinoma PC (AdPC).
Antibodies
The immunoblotting technique used was previously described (11). Antibodies are as follows: actin (MAB150-1R; Chemicon-Millipore), PSMA (D4S1F; Cell Signaling), AR (sc-816; Santa Cruz Biotechnology), NSE (sc-271384; Santa Cruz Biotechnology), GCK (sc-17819; Santa Cruz Biotechnology), and GLUT12 (ab100993; Abcam).
Data-Mining Analysis
Using 268 PC samples from 3 different cohorts, we assessed the transcript abundance for all of the SLC2A family (SLC2A1-14) and the hexokinase family (HK1–HK4, HK4 referred to as GCK). In addition, we monitored the PSMA gene (FOLH1), SRRM4 as a positive marker of treatment-induced NEPC, and REST as a negative marker of NEPC. Patients with a lack of NEGS or neuroendocrine histopathology are referred to as AdPC.
To assess the lineage plasticity of mCRPC, samples from a tissue acquisition necropsy platform established at the University of Washington (UW) were used (3). All rapid autopsy tissues were collected from patients who gave written informed consent under the aegis of the PC Donor Program at the UW, and the Institutional Review Board of the UW approved this study. We classified our mCRPC subtypes as AdPC (AR-positive/neuroendocrine-negative), AR-suppressed AdPC (low AR/neuroendocrine-negative), NEPC (AR-negative/neuroendocrine-positive), and DNPC (AR-negative/neuroendocrine-negative). In addition, we used the Beltran cohort (4) with histologically confirmed mCRPC-AdPC and mCRPC-NEPC samples. We identified a NELPC subset among AdPC tumors from the Memorial Sloan Kettering Cancer Center cohort (24) using the meta-signature of prototypical high-grade NEPC (25). Gene set enrichment analysis was performed on the identified subsets using gene sets downloaded from the Molecular Signatures Database (26).
Mouse PDX Models
Fresh PC tissues from patients were grafted under the kidney capsules of nonobese diabetic or severe combined immunodeficient mice. The Institutional Review Board and Animal Care Committee of the University of British Columbia approved this study, and all subjects gave written informed consent. We previously characterized and validated these models (27).
GB2-Cy3 Synthesis and Cellular Uptake
Synthesis and in vitro uptake of a glucose bioprobe, GB2-Cy3, were previously reported with some modifications (22,28,29). Full experimental details are provided in the supplemental material, and Supplemental Schemes 1 and 2 summarize the GB2-Cy3 synthetic strategy (supplemental materials are available at http://jnm.snmjournals.org).
In Vivo Glucose Uptake Imaging
Wild-type zebrafish (Danio rerio) were maintained following the Canadian Council on Animal Care Guidelines. In vivo uptake of GB2-Cy3 was visualized in a zebrafish model by modifications of previous protocols (23,30). Full experimental details are provided in the supplemental material. This study was approved by the University of Windsor Animal Care Committee.
Statistical Analysis
Statistical analysis was done using GraphPad Prism. The results are expressed as the mean ± SEM. The box–whisker plots show the median (horizontal line), the interquartile range (margins of box), and the absolute range (vertical line). Differences between 2 groups were compared by unpaired Student t testing. One-way ANOVA was followed by a Benjamini–Hochberg or Tukey adjustment. Neurite length was measured by manual tracing and determined using National Institutes of Health ImageJ software as previously described (11,31). Pearson correlation was used for nearest-neighbor analysis and pairwise correlation of the studied genes. Kaplan–Meier plots and heat maps were generated using camcAPP (32) and Broad Institute Morpheus software.
RESULTS
Differential Expression of FOLH1, SLC2A, and Hexokinase in mCRPC
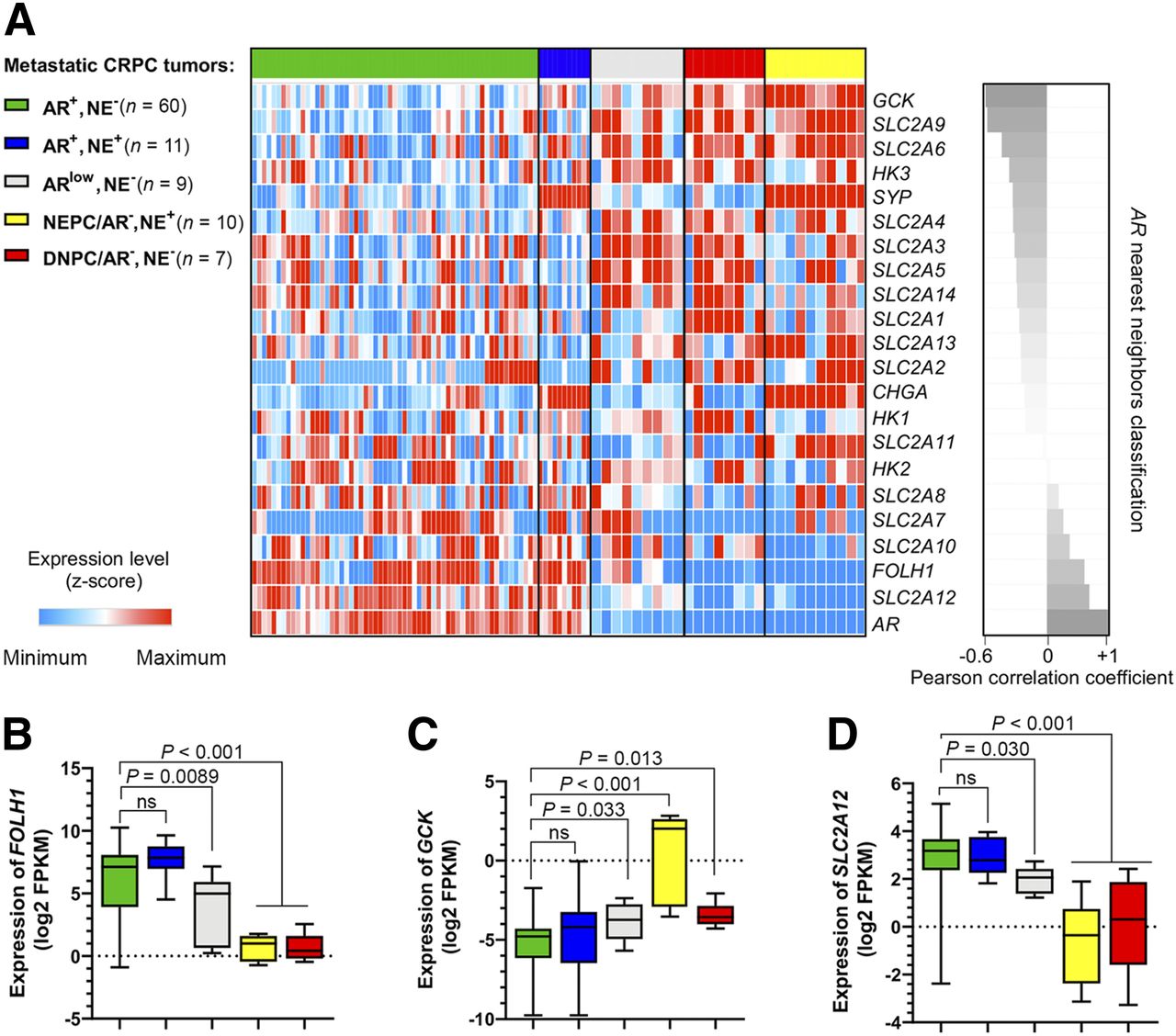
Figure 1A shows that expression of SLC2A12 and FOLH1 are the nearest neighbors to AR (r > 0.6, P < 0.01) and GCK is the farthest neighbor (r = −0.6, P < 0.01) in the UW cohort (3). We observed a significant suppression of FOLH1 in low-AR mCRPC phenotypes, including NEPC and DNPC samples (Fig. 1B). Figure 1C shows that NEPC tumors have a 5-fold elevation of GCK (P < 0.0001) when compared with AR-positive samples. Alternatively, Figure 1D demonstrates that NEPC and DNPC samples suppress expression of SLC2A12. Supplemental Figure 1 verifies that in the Beltran cohort (4) FOLH1-suppressed NEPC samples have similar profiles of GLUT gene expression. In summary, GCK gene expression is elevated and the SLC2A12 gene is suppressed in NEPC.
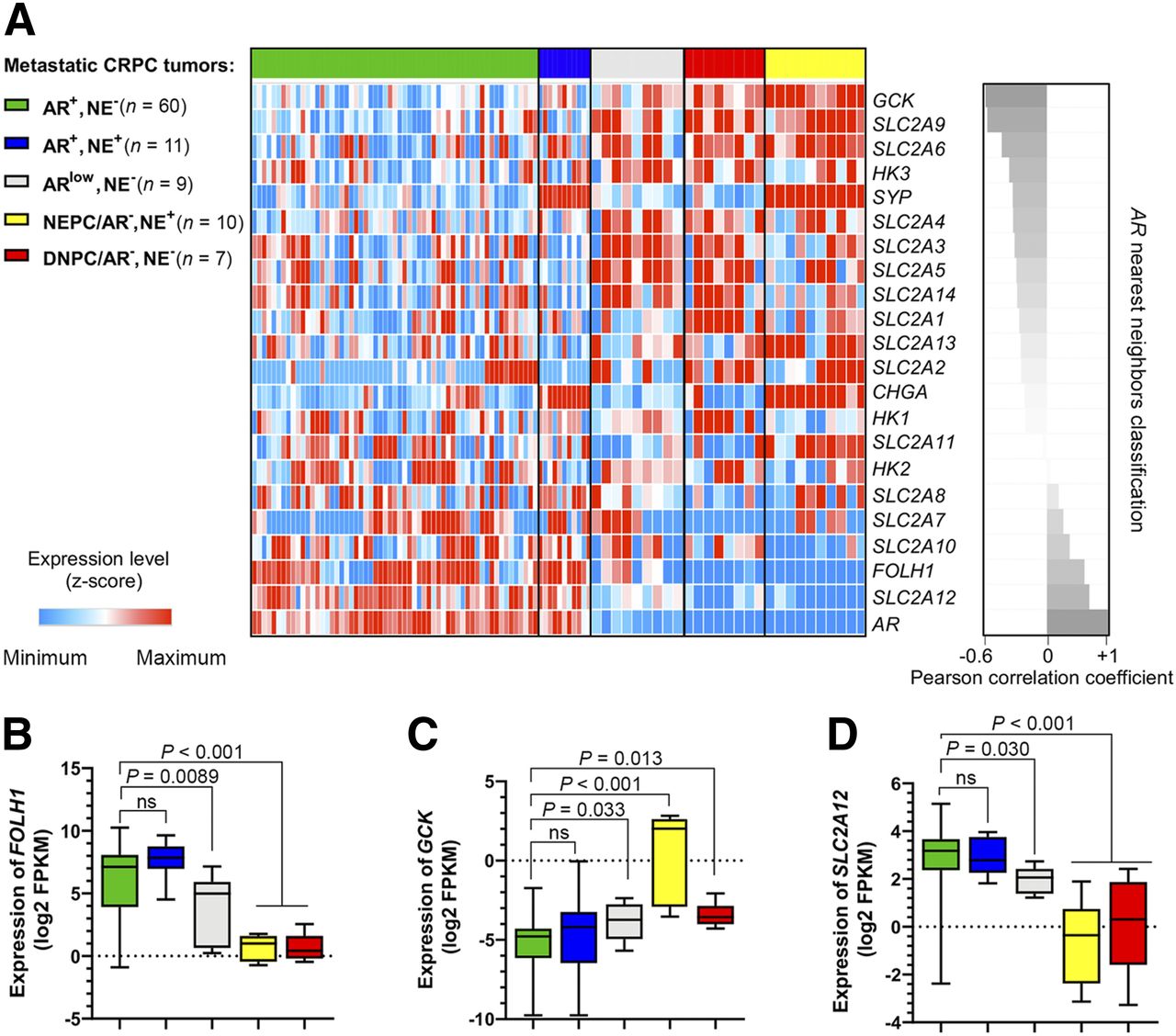
AR-negative subsets of mCRPC in UW cohort (3) have suppressed expression of FOLH1 and differential expression of SLC2A and hexokinase genes. (A) Heat-map plot of expression levels of SLC2A family members, hexokinase genes, AR, FOLH1, and neuroendocrine markers sorted on the basis of nearest neighbors clustering to AR. (B, C, and D) Box–whisker plots showing expression of FOLH1, GCK, and SLC2A12, respectively.
Differential Expression of SLC2A and Hexokinase in NELPC
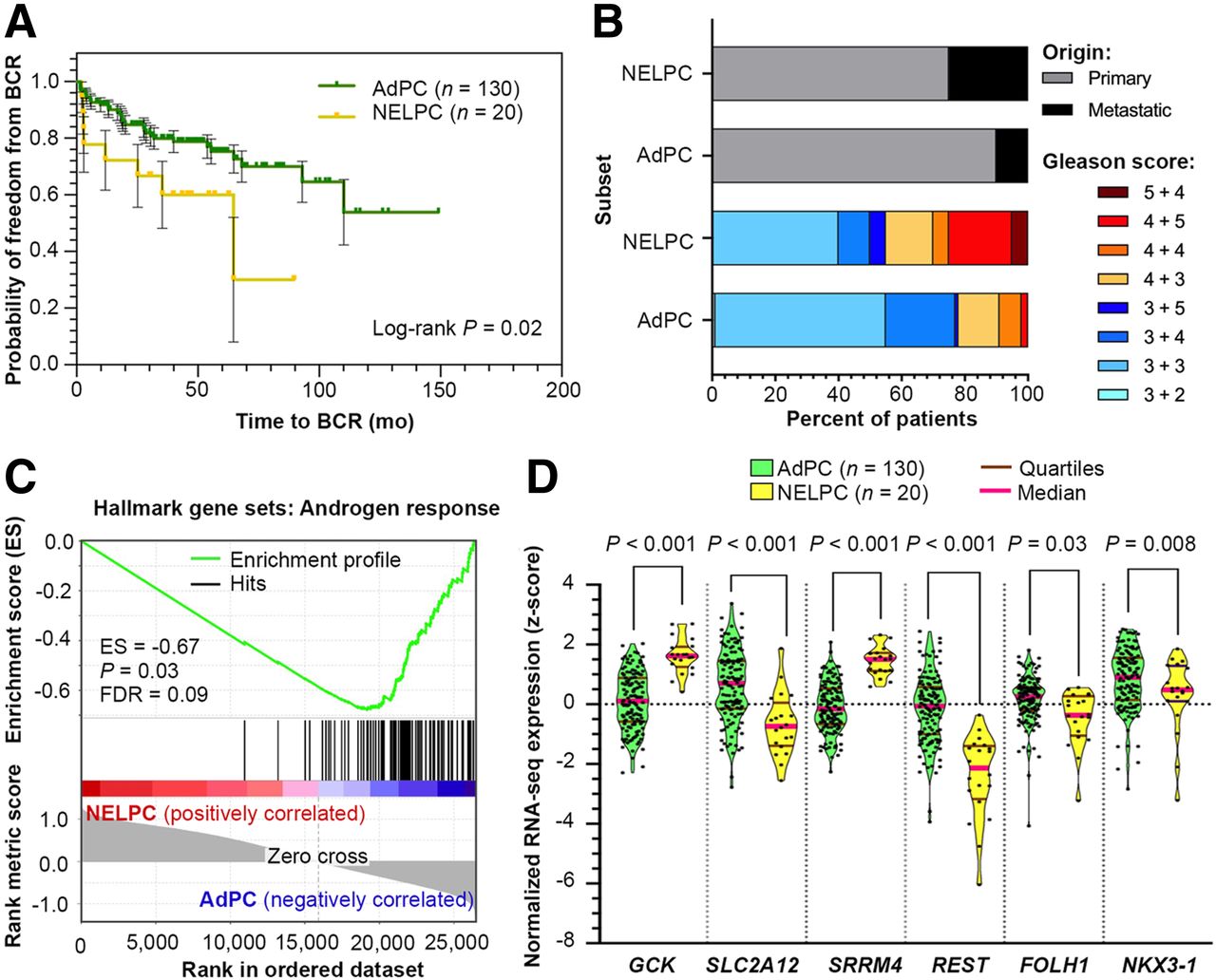
The meta signature of prototypical high-grade NEPC (25) was used to isolate a potential NELPC group among a population of metastatic and primary AdPC samples lacking NEPC histopathology (Supplemental Figs. 2 and 3). Figure 2A shows that the transcriptionally identified NELPC subset had a shorter time to biochemical recurrence in NELPC (log-rank P = 0.02). Figure 2B shows a NELPC hallmark in both primary and metastatic samples, with the more prevalent signature seen in samples that are metastatic and have a high Gleason score. Figure 2C shows a lack of hallmarks of AR response in NELPC. Supplemental Figure 4 shows that SLC2A1, SLC2A3–5, SLC2A9, SLC2A10, SLC2A12–14, HK1, and HK2 genes cluster with REST; these are herein referred to as REST-clustered genes. On the other hand, SLC2A2, SLC2A6–8, SLC2A11, HK3, and CGK cluster with SRRM4 and other neuroendocrine-markers; these are herein referred to as SRRM4-clustered genes. Pairwise correlation with SRRM4 expression is presented in Supplemental Figures 5–7. Similar to NEPC, SLC2A12 and FOLH1 expression is decreased in NELPC relative to AdPC (Fig. 2D). GCK expression is significantly higher in NELPC.
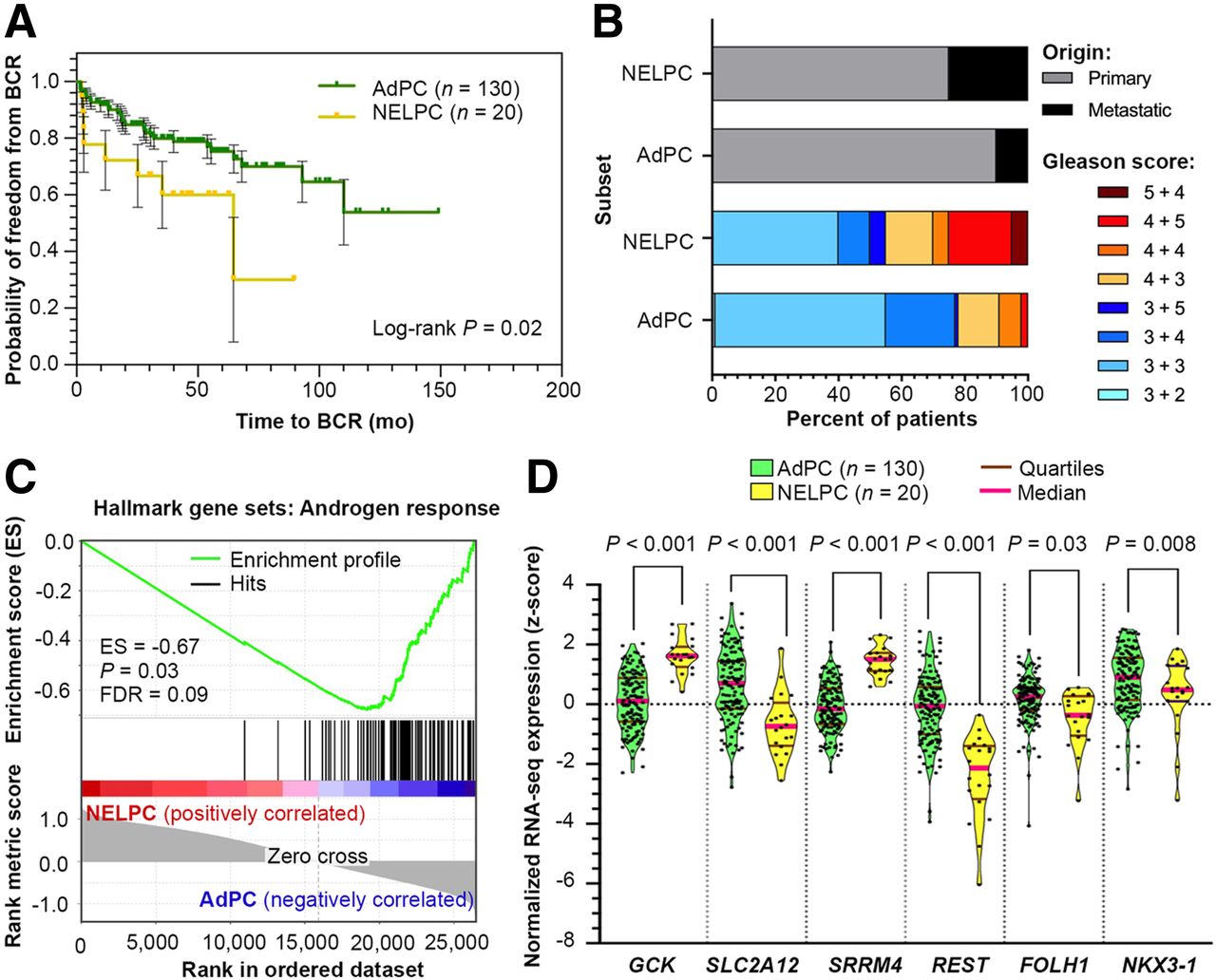
NELPC subset of Memorial Sloan Kettering Cancer Center cohort (24) displays shorter time to BCR (A), greater prevalence of metastatic and high Gleason score specimens (B), and lack of AR response (C). (D) Violin plots compare distribution of SLC2A12, GCK, and FOLH1 expressions in NELPC and AdPC subsets. BCR = biochemical recurrence; ES = enrichment score; FDR = false discovery rate.
The Association of SLC2A and Hexokinase with Gleason Score and Biochemical Recurrence in NELPC
Supplemental Figures 8–10 depict expression levels of the studied genes during progression of AdPC based on Gleason score. Most REST-clustered SLC2A genes and hexokinases are either unchanged or suppressed at high Gleason scores, whereas SRRM4-clustered genes are significantly increased in samples with high Gleason scores. Kaplan–Meier survival curves studying high and low expression levels of the studied genes are represented in Supplemental Figures 11–13. The high expression of SRRM4-clustered genes such as GCK and REST-clustered gene SLC2A1 (as an exception) is significantly associated with decreased biochemical recurrence (log-rank P = 0.015 for GCK). Interestingly, high levels of REST-clustered genes, including SLC2A12, are associated with a shorter time to biochemical recurrence (log-rank P = 0.012 for SLC2A12). Supplemental Table 1 summarizes the performed analysis on NELPC.
Sharing of SLC2A12 Suppression and GCK Overexpression Among NEPC and NELPC
RNA-sequencing data from 268 PC samples from the Memorial Sloan Kettering Cancer Center (24), Beltran (4), and UW (3) cohorts were used to stratify SLC2A1–SLC2A14 and HK1–HK4 genes into neuroendocrine-clustered and AdPC-clustered groups (Supplemental Fig. 14). The intersection between the clustered genes in different cohorts and inclusion of the most differentially expressed genes showed that GCK is the most highly expressed gene and SLC2A12 is the most suppressed gene in samples with an NEGS.
The Distinct GCK-Amplified and SLC2A12-Suppressed Signature of NEPC in PDX Models
The LTL331 PDX is a model of PC progression from AdPC to NEPC. LTL331 tumors regress after castration but relapse within 24–32 mo with tumors harboring NEPC phenotypes (27). Figure 3A demonstrates that GCK expression is minimal before progression to CRPC but maximizes after cellular plasticity to CRPC and NEPC. Conversely, SLC2A12 expression maximizes in hormone sensitive AdPC and, with a slight fluctuation, gradually decreases after castration. The expression of SLC2A12 and FOLH1 is the nearest neighbor to AR (r > 0.6, P < 0.01), and GCK is the farthest neighbor (r = −0.87, P < 0.01) in the UW cohort (3). Figure 3B shows—in our other well-characterized PDX models consisting of 20 AdPC and 3 NEPC models—that we observed significant elevation of GCK and suppression of SLC2A12 gene expression in the NEPC models. Overall, NEPC models have a SLC2A12-low and GCK-high signature.
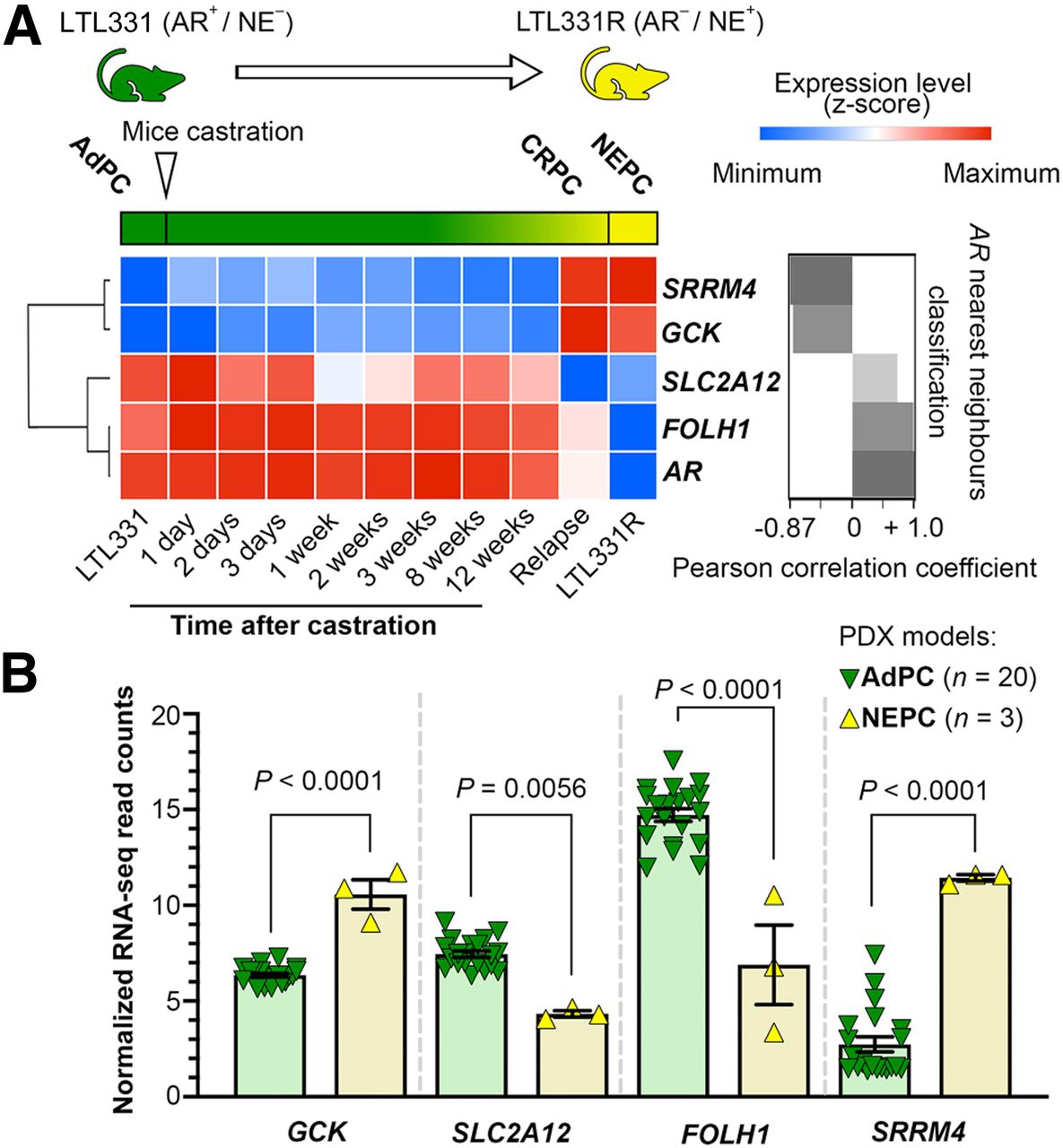
NEPC PDX models have suppressed expression of FOLH1 and differential expressions of SLC2A12 and GCK. (A) Transcription of studied genes during progression to NEPC and correlation with AR. (B) Expression of studied genes in PDX models.
Higher In Vitro and In Vivo Glucose Uptake in Neuroendocrine-Induced Cell Lines
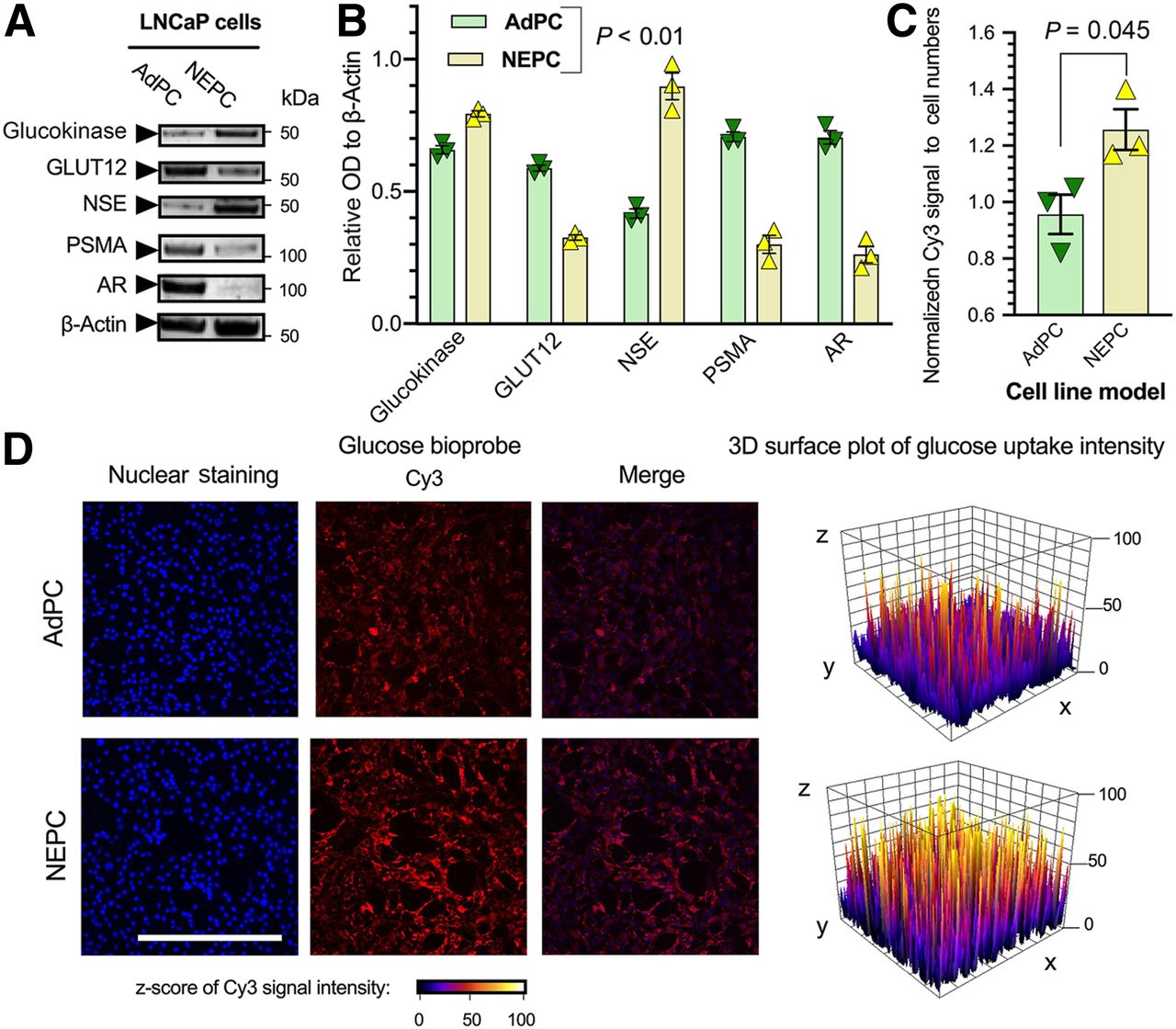
To investigate the role of progression to a neuroendocrine phenotype on glucose uptake, we used the well-characterized neuroendocrine subclone cell line (LNCaP-NEPC) in which NEPC cells are derived from LNCaP cells (LNCaP-AdPC) by culturing in an androgen-depleted environment to mimic clinical androgen-deprivation therapy (Supplemental Figs. 15A, 15B, and 15D) (33). Figure 4A shows that protein levels of the neuroendocrine marker NSE are increased in the LNCaP-NEPC, whereas PSMA and AR levels are significantly decreased. The LNCaP-NEPC cell line has a significantly higher level of glucokinase (GCK) protein and a significantly lower level of GLUT12 (Fig. 4B).

Neuroendocrine-induced LNCaP cell line represents higher glucose uptake and differential protein levels of glucokinase and GLUT12. (A and B) Western blot analyses of protein levels. (C and D) Quantification of GB2-Cy3 uptake and representative images of LNCaP cells. Scale bar = 200 μm.
Supplemental Figures 16–23 chemically characterize GB2-Cy3. Supplemental Figure 24 illustrates GB2-Cy3 uptake and its localization in LNCaP cells. Figure 4C shows that the LNCaP-NEPC cell line has a higher in vitro uptake of GB2-Cy3. Similarly, a zebrafish model was used for nonradioactive in vivo imaging of glucose uptake and displayed higher GB2-Cy3 in engrafted LNCaP-NEPC cells (Fig. 5). These observations indicate that suppression of PSMA and AR and elevation of neuroendocrine markers in LNCaP cell lines are associated with a differential level of glucose uptake, suppression of GLUT12, and elevation of glucokinase proteins.
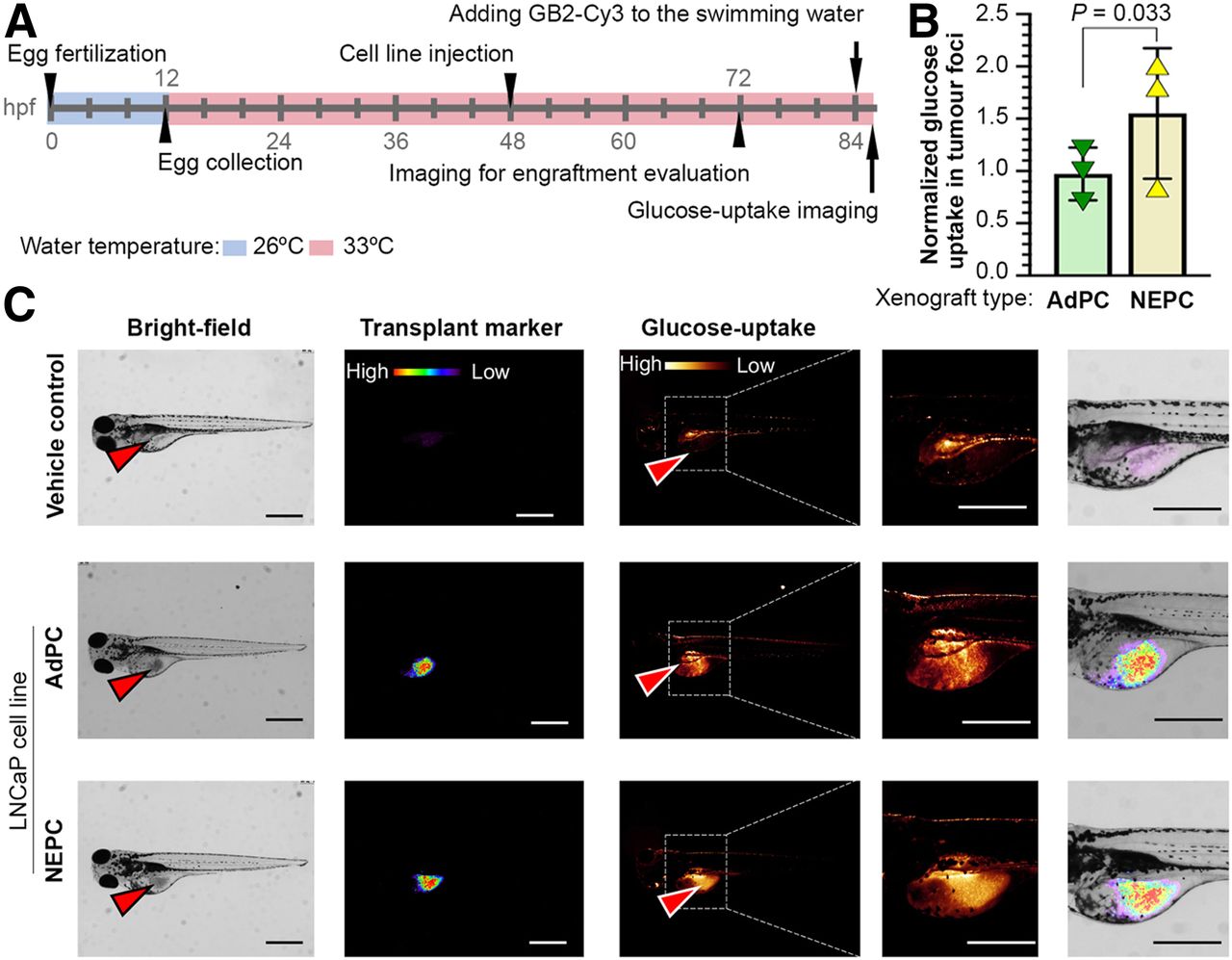
Neuroendocrine-induced LNCaP cell line xenografts represent higher glucose uptake in zebrafish model. (A) Schematic of experiment. (B and C) Quantification of GB2-Cy3 uptake and representative images of embryos injected with different LNCaP cells. Arrowheads show injection sites. Scale bar = 200 μm.
DISCUSSION
The development of AR-indifferent and neuroendocrine-positive tumor phenotypes through divergent clonal evolution as a mechanism of resistance to AR inhibition in mCRPC is a well-characterized concept (3,4). However, Stelloo et al. (2) identified an NELPC in a treatment-naïve and primary cohort. Our work identified an NELPC subset among primary and metastatic samples with no history of exposure to next-generation AR inhibitors. Our data also show that the incidence of NELPC is more prevalent in metastatic specimens. These data support that either AR-indifferent subsets of cells can exist among AdPC that possess a greater susceptibility for neuroendocrine transdifferentiation or AR-indifferent, neuroendocrinelike cells exist from an early time point and are gradually selected for during treatment pressures.
Clinically, there are data to support a correlation between the detection rate of 68Ga-PSMA-11 and the prostate-specific antigen level (7). Also, patients with low levels of prostate-specific antigen show lower PSMA-radioligand uptake and higher 18F-FDG uptake (18,20). This relationship supports the goals of this study to investigate the mechanistic basis for 18F-FDG avidity in NEPC and DNPC. Here, we show that the isoforms of GLUTs are structurally and functionally related proteins with different affinities to glucose. They are expressed in different cells on the basis of the metabolic necessity for glucose uptake (34). The elevation of glycolysis in NEPC has been previously reported (35,36). Irrespective of the overall contribution of GLUT to glucose metabolism, GLUT and hexokinase family members could be associated with 18F-FDG uptake (13,14). Like glucose, 18F-FDG is phosphorylated by hexokinases whereas their products, glucose-6-phosphate and FDG-6-phosphate, can have different levels of inhibition on hexokinases depending on their structure (37). Supplemental Figure 25 represents structural domains of the isoforms of human hexokinase proteins (38). Glucokinase lacks the N terminus domain and cannot be inhibited by either glucose-6-phosphate or FDG-6-phosphate.
GLUT11 is considered a high-affinity GLUT and can be effectual in elevation of glucose uptake while it is amplified (34). McBrayer et al. (39) evaluated the association of GLUT11 expression and 18F-FDG uptake in multiple myeloma. We have observed a significant elevation of GLUT11 expression in both NEPC and NELPC. Similarly, GLUT7 and GLUT8 have a high affinity to glucose (40), and we observed their upregulation in samples with NEGS. Contrary to HK1–HK3, glucokinase, which is known as a glucose sensor in pancreatic β-cells, is not inhibited by its product glucose-6-phosphate but remains active while glucose is abundant (37,38). We can speculate that the apparent elevation of glucose uptake in NEPC or NELPC could be due to elevation of the expression of the aforementioned high affinity of GLUTs (Supplemental Fig. 26). Although our study provides functional support for this conclusion, it is also important to remember the complications associated with a focus on gene expression of this vast family of glucose transport regulators. Avril’s commentary (13) elaborates on the complex number of molecule-, cell-, tissue-, and organ-related variables regulating the resulting 18F-FDG signal, all of which may provide inconsistencies between GLUT expression and the resulting 18F-FDG signal.
GLUT1 is a high-affinity and basal GLUT expressed ubiquitously in human tissues. In hormone-sensitive PC, GLUT1 gene expression positively correlates with androgen levels (41). Our work implies that GLUT1 expression has limited prognostic potential since it cannot estimate the development or existence of low-AR phenotypes such as NEPC or DNPC phenotype. GLUT12 has recently been introduced as a downstream target of AR, and its expression is tightly regulated by androgens (42). Our data demonstrated that the SLC2A12 suppression is also highly associated with the development of a PSMA-low and neuroendocrine-high gene signature in mCRPC. This work shows that GCK upregulation and SLC2A12 suppression correlate with the PSMA suppression, higher Gleason score, and shorter time to biochemical recurrence in NELPC.
For preclinical studies, the use of 18F-FDG imaging in mouse xenografts can be limited by several factors, such as the operating cost and short half-life of the radioactive substance and nonradioactive glucose probes, which are of particular interest (21). Also, the engraftment of tumors in mice can be challenging to establish and is time consuming. This work demonstrates the feasibility of nonradioactive imaging of glucose uptake in PC xenografts using a zebrafish model as a rapid and cost-effective model.
CONCLUSION
NEPC, DNPC, and NELPC have distinct differential expression of GLUT and hexokinase genes. In accordance with this expression, the suppression of PSMA in NEPC is associated with elevated glucose uptake.
DISCLOSURE
This study was supported by the Canadian Institutes of Health Research (142189 [Lisa Porter] and PJT156150 [Xuesen Dong]), the Natural Sciences and Engineering Research Council of Canada (2018-06338 [John Trant]), and a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health and Welfare, Republic of Korea (H18C1916 [Gi Jeong Cheon]). Financial support was also provided by TELUS Ride for Dad, the PC Fight Foundation (Lisa Porter), and the Ontario Trillium Scholarship Program (Martin Bakht). The gene expression analysis was supported in part by a Department of Defense Idea Development Award-Partnering-PI (W81XWH-17-1-0414;W81XWH-17-1-0415 [Colm Morrissey]) and the Pacific Northwest PC SPORE (P50CA97186 [Colm Morrissey]). No other potential conflict of interest relevant to this article was reported.
KEY POINTS
QUESTION: Do the expression levels of glucose uptake–associated genes correlate with the development of NEGS or suppression of the PSMA gene?
PERTINENT FINDINGS: Data-mining approaches, cell lines, mouse, and zebrafish PDX models were used to demonstrate that GLUT and hexokinase expression, specially GCK and SCL2A12, is associated with NEGS, PSMA suppression, and higher glucose uptake.
IMPLICATIONS FOR PATIENT CARE: This study supports the use of 18F-FDG PET for imaging of low-PSMA PC tumors with NEGS.
Footnotes
Published online Dec. 5, 2019.
- © 2020 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication June 2, 2019.
- Accepted for publication October 22, 2019.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A Compound that Inhibits Glycolysis in Prostate Cancer Controls Growth of Advanced Prostate Cancer
- The VISION Forward: Recognition and Implication of PSMA-/18F-FDG+ mCRPC
- Identification of alternative protein targets of glutamate-ureido-lysine associated with PSMA tracer uptake in prostate cancer cells
- The Added Value of 18F-FDG PET/CT Compared with 68Ga-PSMA PET/CT in Patients with Castration-Resistant Prostate Cancer
- Identification of PCWG3 Target Populations Is More Accurate and Reproducible with PSMA PET Than with Conventional Imaging: A Multicenter Retrospective Study