


Abstract
Clinical radioimmunotherapies with anti-CD20 monoclonal antibodies involve administering a predose of unlabeled anti-CD20 antibodies to favorably alter the biodistribution profile of the subsequently administered radiolabeled antibodies and mediate antitumor effects. Prior in vitro data suggested that unlabeled anti-CD20 monoclonal antibodies radiosensitize lymphoma cells as well. We assessed the antiproliferative and possible radiosensitizing capabilities of an anti-CD20 monoclonal antibody, rituximab. Methods: Luciferase-transfected (via a lentivirus vector) CD20+ human Raji lymphoma cells in log-phase growth were incubated with or without rituximab (20 μg/mL) for either 1 or 24 h before external-beam radiation exposure. Cell counts were measured with a luciferase assay at 24-h intervals. Subsets of these cells were also analyzed for cell cycle status by flow cytometry. Results: Rituximab pretreatment and irradiation were found to significantly inhibit tumor cell growth compared with irradiation alone (by a factor of 0.40 at 1 Gy [P < 0.01]). One hour of rituximab pretreatment modestly radiosensitized tumor cells at a radiation dose of 1 Gy (by a factor of 1.03 compared with the results for nonirradiated cells). At higher radiation doses (2 and 12 Gy), 1 h of rituximab pretreatment paradoxically radioprotected tumor cells by factors of 0.25 (P < 0.01) and 0.54 (P < 0.05), respectively. Rituximab predosing for 24 h was found to be radiosensitizing at a radiation dose of 4 Gy (by a factor of 2.84 [P < 0.01]) but radioprotective at radiation doses of 1, 8, and 12 Gy (by factors of 0.10 [P < 0.01], 2.50 [P < 0.01], and 2.07 [P < 0.05], respectively). These results correlated with retardation of the cell cycle at 6 d after rituximab administration, as determined by flow cytometry. Conclusion: Rituximab demonstrated a direct tumor antiproliferative effect in the absence of radiation. At lower levels of radiation exposure, rituximab radiosensitized Raji lymphoma cells. At higher doses of radiation, rituximab paradoxically protected tumor cells against ionizing radiation, possibly through effects on the cell cycle. These radiobiologic effects of rituximab should be carefully considered in the design of radioimmunotherapeutic trials.
Unlabeled anti-CD20 antibodies have been shown to mediate antitumor effects in animal model systems for several decades (1). Rituximab, an anti-CD20 chimeric, recombinant monoclonal antibody, is one such antibody. First described in 1994 (2), its use and efficacy in treating non-Hodgkin's lymphoma have been established such that it was granted U.S. Food and Drug Administration approval for non-Hodgkin's lymphoma therapy 3 y later. Although the precise in vivo mechanism of anti-CD20 antibodies is not completely understood, in vitro data suggest that they exert their effects through antibody-dependent cellular cytotoxicity (3), direct activation of apoptosis (4,5), cell-mediated cytotoxicity (6,7), or cell cycle arrest (8). In addition to their use in various chemotherapeutic approaches for lymphoma, anti-CD20 antibodies are also the main elements of the only 2 U.S. Food and Drug Administration–approved radioimmunotherapeutic regimens (9).
Radioimmunotherapy (RIT) involves the targeting of a radioactive monoclonal antibody to selectively deliver ionizing radiation directly to tumors. At present, 2 approved agents are available: tositumomab and 131I-tositumomab (Bexxar; GlaxoSmithKline) and 90Y-ibritumomab tiuxetan (Zevalin; Cell Therapeutics, Inc.). Protocols for both agents take into consideration the need to decrease the specific sequestration of labeled antibodies by highly accessible “antigen sinks,” including the spleen and circulating CD20+ lymphocytes. Predosing with unlabeled antibodies prevents unwanted normal-tissue toxicity by making the biodistribution profile of radiolabeled antibodies more predictable, decreasing their clearance rates (10) and increasing their percentage injected dose (11,12). However, an important radiobiologic question arises: Can such an anti-CD20 monoclonal antibody predose affect the radiosensitivity of targeted tumor cells?
Although rituximab has been reported to be a radiosensitizing agent, to date there are limited data supporting this report (13). As mentioned earlier, a purported mechanism of anti-CD20 antibody action is arrest of the cell cycle (8). We hypothesized that if rituximab arrests lymphoma cells in the radioresistant G0/G1 phase of the cell cycle (14), then it may be radioprotective.
The aim of the present study was to determine whether there were any clear radioprotective or radiosensitizing effects of rituximab at expected concentrations in serum before exposure to various single-fraction external-source radiation doses. Cell growth assays with selective cell cycle analyses were performed to determine whether and by what mechanism rituximab predosing might affect the radiosensitivity of Raji lymphoma cells.
MATERIALS AND METHODS
Cells and Cell Culture
Epstein-Barr virus–infected Raji lymphocyte tumor cells transfected via a lentivirus vector with green fluorescent protein (GFP) and luciferase were established by Dr. Zhaohui Ye in collaboration with the laboratory of Dr. Linzhao Cheng at The Johns Hopkins University (Figs. 1A and 1B) (15). A total of 105 cells in the exponential phase of growth were incubated in 10 mL of CellGro RPMI medium (Mediatech) containing 10% fetal bovine serum (Hyclone) and 1% penicillin–streptomycin (Invitrogen Corp.) over a total of 6 d and maintained at 37°C with 5% CO2 (in a Forma Series II Water-Jacketed Incubator; Thermo Fisher Scientific Inc.). Cells were plated in triplicate.

(A) Lentivirus transfection cassette. (B) Fluorescence microscopy of transfected cells, demonstrating stable GFP expression. EF1α = elongation factor 1α; IRES = internal ribosome entry site; 5′LTR = 5′ long terminal repeat; 3′LTR = 3′ long terminal repeat; LUC = luciferase.
Cell Viability Assay
Cells were initially counted with a BrightLine hemocytometer (Reichert), and cell counts obtained in this manner were then correlated with luciferase activity. In brief, luciferin (MIP) was added to a known quantity of cells, and the mixture was incubated in the dark for 2 min before 30 s of photon counting with a Monolight 3010 luminometer (Pharmingen/Becton-Dickinson) in accordance with the manufacturer's instructions. Standard curves were generated in this manner to ensure that a linear measurement was available over the ranges of the cells to be studied (3 × 101–3 × 106 cells per milliliter). After verification of the standard curves, cells were counted with the luciferase assay at 24-h intervals for 6 d. All experiments were performed in triplicate.
Cell Growth Rate
The cell growth rate was determined by averaging the slopes of cell growth data on the most linear portion of the plot (with a logarithmic vertical axis). This plot corresponded to sections of the growth curve between days 1 and 5, with R2 values generally in the range of 0.90–0.99.
Antibody
Pharmaceutical-grade rituximab (Genentech/Biogen IDEC Corp.) was kindly provided by the Sidney J. Kimmel Cancer Center at The Johns Hopkins University and stored at 4°C. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis was used to confirm the presence of intact, 150-kDa antibody molecules. Raji lymphoma cell surface expression of CD20 and antibody immunoreactivity were verified with a quantitative CD20 assay followed by a competitive inhibition assay (antibody source: Quantibrite CD20 PE; BD Biosciences) performed by use of a FACS-Caliber quantitative flow cytometer (Becton-Dickinson Immunocytometry Systems). A rituximab dose of 20 μg/mL was administered to cells either 1 or 24 h before irradiation in accordance with expected peak levels in serum given current dosing strategies.
Radiation
Transfected Raji lymphoma cells that were either not treated or treated with rituximab were either not exposed or exposed to various amounts of external-source cesium-based γ-radiation (none, 1 Gy, 2 Gy, 4 Gy, 8 Gy, or 12 Gy) on a Gammacell 3000 Elon apparatus (MDS-Nordian). Ionizing radiation was delivered at a rate of 5 Gy/min. Control cells that were not irradiated were removed from the incubator and transported to the radiation site for the period of irradiation of the other cells.
Cell Cycle Analysis
Cell cycle status was assayed at 5 d after irradiation with a Vybrant Violet DyeCycle apparatus (Invitrogen Probes) in accordance with the manufacturer's instructions. An LSRII flow cytometer (Becton-Dickinson Immunocytometry Systems) was used to quantitate cellular DNA; the resulting data were used to establish the cell cycle status. Gating was set in accordance with the manufacturer's instructions, with the single largest peak among the nonirradiated (control) cells being calibrated to cells with 2N DNA (normally occurring diploid DNA seen in nondividing cells) and the next largest peak being calibrated to cells with 4N DNA (tetraploid DNA seen in cells after DNA replication but before cell division). Interval areas under the curve were determined by an individual who was experienced in flow cytometry and were kept constant between samples.
RESULTS
Antiproliferative Effect
A clear dose–response effect was observed across samples over the 5-d observation period (Figs. 2 and 3). At levels of 8 and 12 Gy, an overall death of cells to below starting cell numbers was observed by day 5. In a comparison of samples receiving rituximab and control samples, there was a statistically significant decrease in cell survival at most levels of radiation when rituximab was added (Table 1 and Figs. 2 and 3). However, at the highest doses of radiation, the addition of rituximab proffered no additional benefit in terms of overall cell killing.
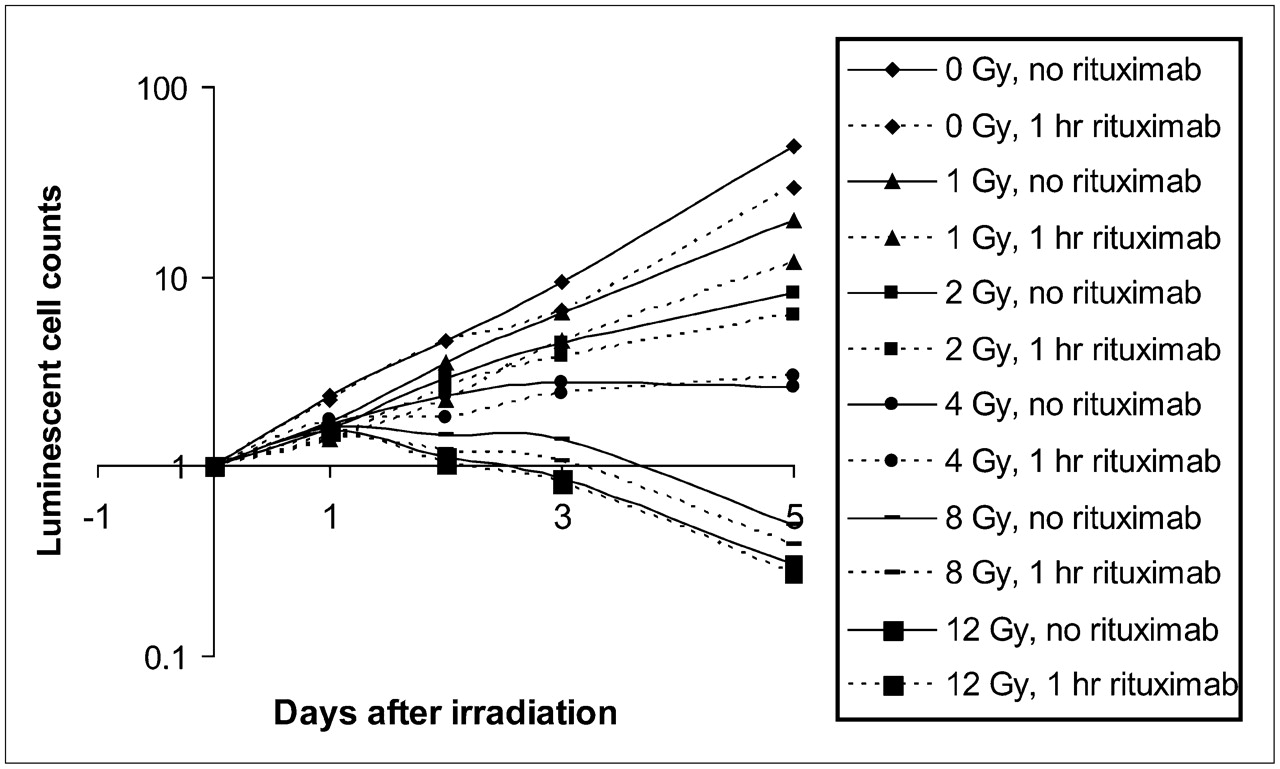
Effect of 1-h rituximab predose (20 μg/mL) on cell survival. Dose–response relationship was observed with regard to cell growth rate in response to various amounts of radiation exposure. Each data point is normalized to starting cell count and represents mean of 3 determinations.
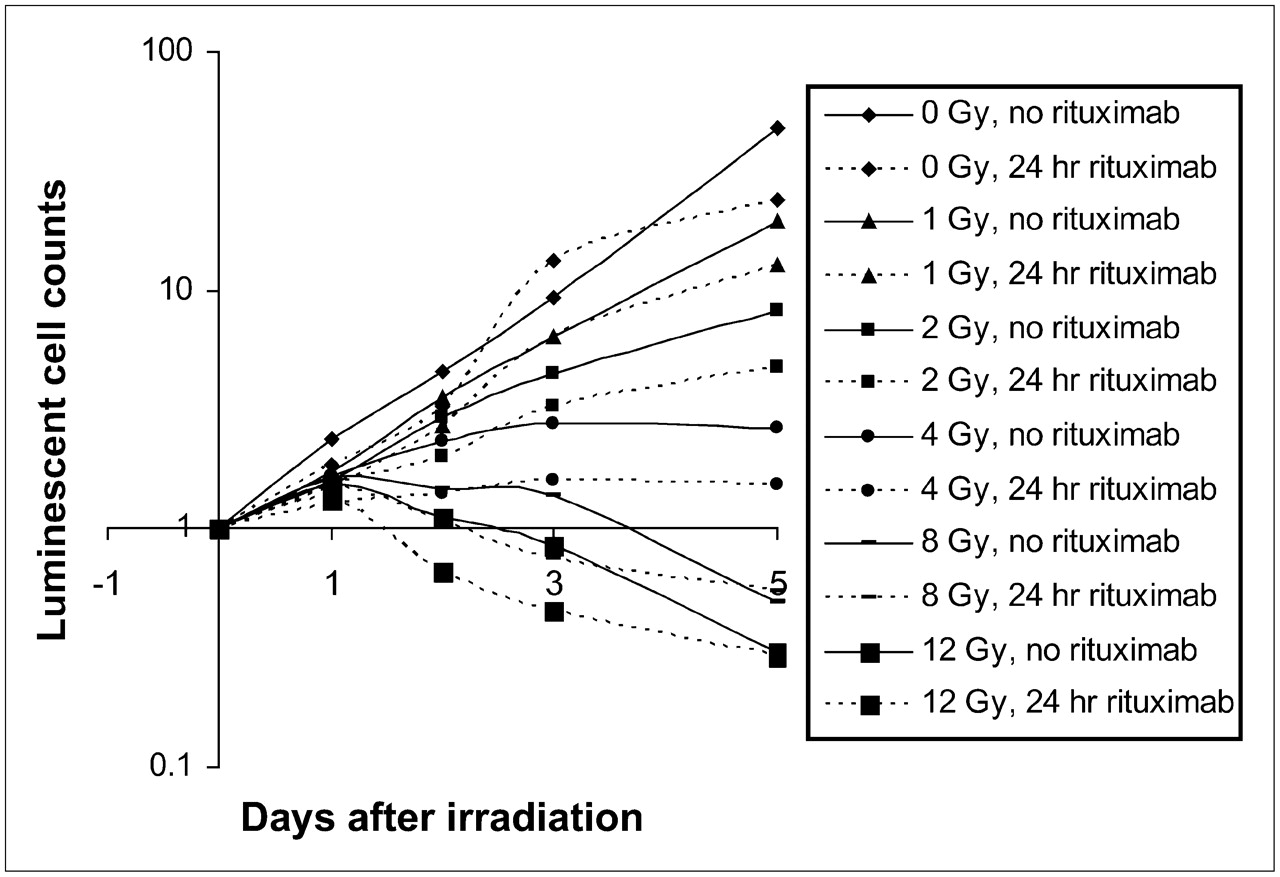
Effect of 24-h rituximab predose on cell survival. Dose–response relationship was observed with various amounts of radiation exposure. Each data point is normalized to cell count at day of irradiation and represents mean of 3 determinations.
Absolute Cell Growth over 5 Days for Cells Treated with Rituximab Predose or No Predose (Control Cells)*
Cell Growth Rate
A clear dose–response relationship was also evident with regard to the rate of cell growth and increasing levels of radiation (Table 2). With no radiation, 1 Gy, 2 Gy, and 4 Gy of radiation, both 1- and 24-h rituximab predosing significantly decreased the rate of cell growth compared with the results for cells that were exposed to similar radiation doses in the absence of rituximab. At 8 and 12 Gy, however, 24-h rituximab predosing significantly decreased the overall rate of cell death compared with the results for cells that were irradiated at similar radiation doses but were not exposed to rituximab (P < 0.01 and P < 0.05, respectively).
Mean Cell Growth Rates over 5 Days*
Cell Cycle Analysis
At 5 d after irradiation, nonirradiated cells demonstrated a modest but statistically significant increase in the percentage of cells in the G0/G1 phase when rituximab was administered 24 h before irradiation (Table 3). This finding correlated with a statistically significant decrease in the percentage of cells in the S and G2/M phases. Among cells exposed to 1 Gy of radiation, a modest trend toward a decreased percentage of cells in the S phase was observed, and at 8 Gy of radiation, there was again a modest but statistically significant increase in the percentage of cells in the G0/G1 phase. There was also a statistically significant decrease in polyploidy among cells receiving 8 Gy of radiation and rituximab compared with the results for control cells. Among cells receiving 12 Gy of radiation, no differences in the cell cycle were observed between cells receiving rituximab and control cells. However, a large number of hyperdiploid cells were seen.
Results of Cell Cycle Analyses of Various Groups of Cells Receiving Radiation, Predose of Rituximab, or Both*
DISCUSSION
The purpose of these in vitro experiments was to determine whether rituximab was protective, sensitizing, or noncontributive to the net effect of single-fraction external-beam radiation therapy given at low and high levels. In our first set of experiments, cells were administered rituximab at 20 μg/mL in combination with increasing doses of radiation and were compared with control cells, which received radiation only. With regard to the experimental design, we were reassured to find that the baseline radiosensitivity of our GFP- and luciferase-transfected (via a lentivirus vector) Raji cells was unchanged with respect to prior published data on the radiosensitivity of nontransfected Raji cells, albeit measured through different methods (16). The dose of rituximab administered was calculated from the known pharmacokinetic parameters of rituximab (17) and fell in the midrange of doses used in various inpatient clinical pharmacokinetic trials (18). The 1-h predose was chosen on the basis of the current predosing strategies for both 131I-tositumomab and 90Y-ibritumomab tiuxetan (19,20). We elected to examine the effect of a longer, 24-h, predose because it would more closely model the circumstance of patients receiving rituximab in addition to external-beam therapy or patients experiencing late dose effects from RIT.
Among cells that received no radiation, there was an observed antiproliferative effect of approximately 50% when rituximab was added. Interestingly, this was the highest observed fractional antitumor effect of rituximab in our experiments. When any amount of radiation was added, only a 34%–42% incremental decrease in cell viability was observed. Furthermore, at 4, 8, and 12 Gy (under various predose conditions), there were no statistically significant antiproliferative effects of rituximab administration above those achieved with radiation alone. Although one might expect synergy between radiation and rituximab, the converse occurred as higher doses of radiation were administered.
With regard to the rates of cell growth and death in the presence of rituximab, the net effect of any rituximab predose in the absence of radiation was to decrease the rate of cell growth by 14%. In terms of radiosensitization versus radioprotection, a radiosensitizer could then be described as an agent that decreases the rate of cell growth beyond 14% in the presence of radiation, and a radioprotectant could be described as an agent that exerts the reverse effect. In this regard, a 13% decrease in the rate of cell growth was observed at 1 Gy (P < 0.01 in a comparison with nonpretreated [control] cells; very modest radioprotection), a 19% decrease was observed at 2 Gy (P was not significant; modest radiosensitization), and a 56% decrease was observed at 4 Gy (P < 0.01; significant radiosensitization). At levels of up to 4 Gy, then, radiosensitizing effects appear to predominate (Table 2).
When the effect of rituximab at 8 and 12 Gy was examined, however, the trend appeared to reverse. The relative rates of cell death decreased by 22% with the addition of rituximab at 8 Gy and by 16% with the addition of rituximab at 12 Gy in comparisons with nonirradiated (control) cells. At these higher levels of radiation, it would appear that rituximab exposure somehow abrogated the cytotoxic effects of radiation (or vice versa). To seek a possible mechanistic correlate, and with an understanding of the antiproliferative effect of rituximab, we undertook an experiment to determine cell cycle status. Our hypothesis was that cell cycle inhibition potentially translated to radioprotection by stalling the cells in a less radiosensitive phase of the cell growth curve, allowing time for DNA repair to occur before replication.
The data from the cell cycle experiments provided definitive evidence that cells incubated with rituximab in vitro demonstrated a modest but nonetheless real and significant alteration in the cell cycle. This alteration resulted in a greater fraction of the cell population being present in the G0/G1 phase, resulting in fewer cells with actively replicating DNA. The prime tenet of therapeutic radiobiology involves the use of ionizing radiation to induce double-strand DNA breakage. This breakage, above all else, leads to errors during DNA replication that cause immediate cell death, or so-called “delayed induced genomic instability,” which is observed as decreased viability for up to several generations after the initial radiologic insult (21). However, if cells are somehow induced to slow or even stop replication, then radiation-induced cell death would be, at the very least, negatively affected. Again, flow cytometric data demonstrated a modest but statistically significant decrease in the percentage of actively dividing cells at 6 d after rituximab administration. This evidence supports our hypothesis that a reduction in DNA replication radioprotects cells exposed to otherwise radioablative doses. Given the high doses experienced by cells exposed to RIT, the data discussed here suggest that a rituximab predose may result in unintended and untoward consequences with regard to the efficacy of tumor cell killing.
CONCLUSION
Although no endogenous CD20 ligand has been identified, prior data suggested that blocking the antigen with an agent such as rituximab can prevent B-cell differentiation and proliferation. It is with this idea in mind that the current sets of experiments were undertaken. For radiation to be effective, it typically requires a cell to be actively dividing before it can be killed. In the present study, we demonstrated that a decrease in the rate of cell division confers a unique advantage against the cytotoxicity of radiation. As a clinical correlate, this finding may translate into modification of the use of rituximab during immunotherapy or even exploration of the use of other (non–CD20-targeting) delivery vehicles. It is important to note, however, that the effect observed was both modest and in vitro. The effect of rituximab and radiation in vivo will undoubtedly prove more complex and thus requires further investigation.
Acknowledgments
Financial support was provided by SPORE-Lymphoma grant P50CA096888 from the National Cancer Institute.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication June 19, 2007.
- Accepted for publication December 13, 2007.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- 177Lu-Lilotomab Satetraxetan Has the Potential to Counteract Resistance to Rituximab in Non-Hodgkin Lymphoma
- A re-examination of radioimmunotherapy in the treatment of non-Hodgkin lymphoma: prospects for dual-targeted antibody/radioantibody therapy
- Pretargeted Versus Directly Targeted Radioimmunotherapy Combined with Anti-CD20 Antibody Consolidation Therapy of Non-Hodgkin Lymphoma