


Abstract
Type 1 diabetes mellitus (T1DM) is characterized by a loss of β-cells in the islets of Langerhans of the pancreas and subsequent deficient insulin secretion in response to hyperglycemia. Development of an in vivo test to measure β-cell mass (BCM) would greatly enhance the ability to track diabetes therapies. β-cells and neurologic tissues have common cellular receptors and transporters, therefore, we screened brain radioligands for their ability to identify β-cells. Methods: We examined a β-cell gene atlas for endocrine pancreas receptor targets and cross-referenced these targets with brain radioligands that were available at our institution. Twelve healthy control subjects and 2 T1DM subjects underwent dynamic PET/CT scans with 6 tracers. Results: The D2/D3 receptor agonist radioligand 11C-(+)-4-propyl-9-hydroxynaphthoxazine (PHNO) was the only radioligand to demonstrate sustained uptake in the pancreas with high contrast versus abdominal organs such as the kidneys, liver, and spleen, based on the first 30 min of data. Mean SUV from 20 to 30 min demonstrated high uptake of 11C-(+)-PHNO in healthy controls (SUV, 13.8) with a 71% reduction in a T1DM subject with undetectable levels of C-peptide (SUV, 4.0) and a 20% reduction in a T1DM subject with fasting C-peptide level of 0.38 ng/mL (SUV, 11.0). SUV in abdominal organs outside the pancreas did not show measurable differences between the control and T1DM subjects, suggesting that the changes in SUV of 11C-(+)-PHNO may be specific to changes in the pancreas between healthy controls and T1DM subjects. When D3 and D2 antagonists were used in nonhuman primates, specific pancreatic binding (SUVR-1) of 11C-PHNO was reduced by 57% and 38%, respectively. Conclusion: 11C-(+)-PHNO is a potential marker of BCM, with 2:1 binding of D3 receptors over D2 receptors. Further in vitro and in vivo studies to establish D2/D3 receptor specificity to β-cells is warranted to characterize 11C-(+)-PHNO as a candidate for clinical measurement of BCM in healthy control and diabetic subjects.
Type 1 diabetes mellitus (T1DM) is characterized by a loss of β-cells in the islets of Langerhans of the pancreas and the subsequent inability to secrete insulin in response to hyperglycemia (1). Current understanding of how the loss of β-cell mass (BCM) contributes to loss of function has been largely determined from postmortem analysis. In the absence of clinically validated methods to measure BCM in vivo, functional studies have been used as a surrogate. However, functional studies, such as C-peptide levels, a measure of endogenous insulin production by β-cells, can be affected by factors including fatty acids, insulin resistance, and even glucose itself. Current tests, such as the oral glucose tolerance test, may not accurately correlate with BCM due to β-cells that are not responsive to stimuli but may still be viable and responsive to treatment. In addition, the sensitivity of β-cells to provocative stimuli may change during different stages of diabetes (2–4). Therefore, development of techniques to measure BCM in vivo would greatly enhance the ability to simultaneously track changes in BCM and function, and to evaluate the efficacy and mechanisms of therapies to preserve or restore insulin secretion (5). Several modalities have been proposed for imaging BCM such as PET, SPECT, MRI, and optical methods (6).
Previous PET studies have attempted to measure BCM by targeting receptors specific to the endocrine pancreas (7–16). Two radioligands with promising results were 18F-FP-(+)-dihydrotetrabenazine (DTBZ), which targets vesicular monoamine transporter 2 (VMAT2) and colocalizes with insulin secretory vesicles (7–9,11), and 11C-5-hydroxytryptophan (5HTP), a precursor of serotonin present in β-cells that may be involved in insulin secretion (10,12,14). For 18F-FP-(+)-DTBZ, binding may not be entirely specific to β-cells as there is some evidence of binding in polypeptide cells (17). For 11C-5HTP, despite demonstrating binding in the islets of Langerhans, it is not clear that binding was specific to β-cells and not due to other endocrine cells (10,12,14). Dopamine, cosecreted with insulin, may act as an autocrine signal via its binding to dopamine receptors (DRs) on the surface of β-cells and thus may be a useful target (18,19).
β-cells and neurologic tissues have common cellular receptors and transporters, and therefore, we screened other brain radioligands for their ability to identify β-cells. We used a β-cell gene atlas to identify possible endocrine pancreas receptor targets and cross-referenced these targets with PET brain radioligands that were available at our institution. Existing radioligands were then examined for high pancreatic uptake and high contrast to neighboring organs to determine which radioligands warranted further study. Those radioligands that exhibited such features were tested in T1DM subjects and nonhuman primates (NHPs).
MATERIALS AND METHODS
Subjects
Twelve healthy control (HC) subjects (6 male/6 female) and 2 male T1DM subjects were included in the study. The average age and weight were 33 ± 9 y and 76 ± 12 kg, respectively. Diabetes duration, at the time of scanning, for T1DM subjects was 29 y (fasting C-peptide undetectable) and 14 y (fasting C-peptide, 0.38 ng/mL); all HCs were screened to exclude existing diabetes (normal fasting C-peptide range, 0.8–3.1 ng/mL). The study was approved by the Yale University Institutional Review Board and Radiation Safety Committees, and all subjects signed a written informed consent form.
β-Cell Gene Atlas Screening
We cross-referenced the targets of available radioligands at the Yale PET Center with a β-cell gene atlas to find radioligands that may be useful BCM biomarkers (20). Relative Affymetrix β-cell gene expression is defined on a 0–100 scale (0–25, no expression; 25–50, low; 50–75, moderate; and 75–100, enriched). All radioligands with a relative expression of 50 or more (moderate or enriched expression) were considered potentially useful.
Human Imaging
On the basis of the gene atlas, we chose 6 radioligands for evaluation. Human PET/CT imaging was performed on a Biograph mCT-X PET/CT system (Siemens Healthcare). A 30-min dynamic scan (6 × 30, 3 × 60, 2 × 120, and 4 × 300 s) centered on the pancreas was performed followed by a 30-min whole-body acquisition (2 passes, 120 s per bed position) followed by an additional 60-min dynamic scan (12 × 300 s) at the level of the pancreas. Injected doses and mass for each radioligand are provided in Table 1. Dynamic scans were reconstructed using an ordered-subset expectation maximization–based algorithm with point-spread function and time-of-flight correction. Attenuation correction was performed using the CT acquisition. Regions of interest (ROIs) were drawn manually on a summed image (0–10 min), then eroded to avoid partial-volume effects, for the pancreas, liver, kidneys, and spleen, and time–activity curves were produced in SUV units. SUV and SUV ratio (SUVR, spleen as reference) (13) were calculated on summed images (20–30 min). No respiratory motion correction was performed; a previous study demonstrated an underestimation of only 15% ± 6% in noncorrected scans for 18F-FP-(+)-DTBZ (21).
Injected Doses and Mass (Minimum–Maximum) for Radioligands and Respective Targets in Human Studies
NHP Imaging
NHP studies were performed to evaluate D2- and D3-specific binding components in the pancreas. A NHP (age, 13 y; weight, 20.9 kg) underwent scans on 2 separate days, with 2 scans per day: baseline followed by a blocking scan with tracer injections separated by 4 h. Mean injected doses were 182 ± 1 MBq, and mass was 0.64 ± 0.10 μg. The blocking scans began 30 min after an infusion of D3-selective antagonist SB277011A (0.3 mg/kg) and D2-selective antagonist L-741,626 (1 mg/kg). Acquisitions on the mCT-X included a 30-min dynamic scan (6 × 30, 3 × 60, 2 × 120, and 4 × 300 s) centered on the pancreas followed by a 4-bed (3 min/bed) whole-body acquisition with 3 passes from head to thigh (36 min total). ROIs were drawn manually on a summed image from 0 to 10 min for the pancreas and spleen. Mean pancreatic SUV and SUVR (spleen as reference) from 20 to 30 min were calculated. Binding potential (BPND) was calculated in the pancreas using simplified reference tissue model (SRTM). In the brain, substantia nigra (SN), caudate, and putamen ROIs were defined from a brain atlas (22) coregistered to the image (30–60 min), and SUV and SUVR (cerebellum as reference) were determined. Using SUVR-1 as a measure of specific binding, we determined percentage reduction in specific pancreatic and brain binding due to the blocking drugs.
To confirm the use of SUVR-1 in blocking studies, we used brain modeling methods in a second NHP (age, 17 y; weight, 14.5 kg) with baseline and blocking brain scans on the Focus 220 (Siemens) with only L-741,626, using the same timing and doses as above. As in previous studies, time–activity curves were generated for brain ROIs, BPND was calculated using SRTM2 (22–24), and regional occupancy was calculated.
RESULTS
We identified 6 potentially useful radioligands targeting 4 different receptor sites: 11C-raclopride, 11C-FLB457, and 11C-(+)-4-propyl-9-hydroxynaphthoxazine (PHNO), and 11C-(+)-PHNO (D2/D3 receptors); 11C-DASB (serotonin transporter); 11C-AS247190 (11β-hydroxysteroid dehydrogenase type 1 (HSD1) enzyme inhibitor) (25); and 11C-UCB-J (synaptic vesicle protein 2A) (26,27). Relative gene expressions of these targets are provided in Table 2. Initially, for β-cell gene expression screening, only the D2 receptor gene was used to identify potential dopaminergic ligands because all D2/D3 radioligands (11C-raclopride, 11C-FLB457, and 11C-(+)-PHNO) target D2 receptors to some extent, although with varying affinities for each receptor. 11C-raclopride and 11C-FLB457 are both receptor antagonists; however, 11C-raclopride (28) has lower affinity than 11C-FLB457 (29). 11C-(+)-PHNO is an agonist with 25- to 50-fold-higher affinity for D3 receptors (22). Retrospective analysis after completion of PET scans to provide insight into differences between the dopaminergic radioligands included the D3 receptor gene and the addition of the Human Primary β-Cells column (Table 2).
Radioligand Target Relative Gene Expression with Rankings of 0–100 Gathered from β-Cell Gene Atlas (20)
11C-(+)-PHNO was the only dopaminergic radioligand to demonstrate sustained uptake in the pancreas, with high contrast to the kidneys, liver, and spleen, based on SUV images (20–30 min) and time–activity curves (0–30 min). Representative PET images (Fig. 1) and time–activity curves (Fig. 2) of the 3 D2/D3 radioligands demonstrated the apparent superiority of 11C-(+)-PHNO.

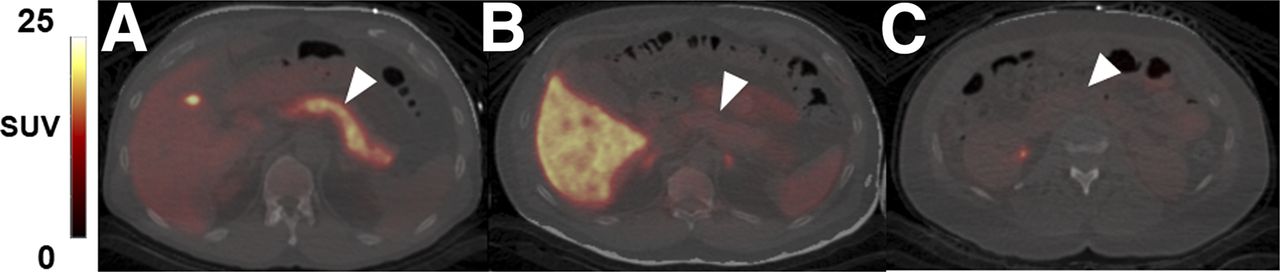
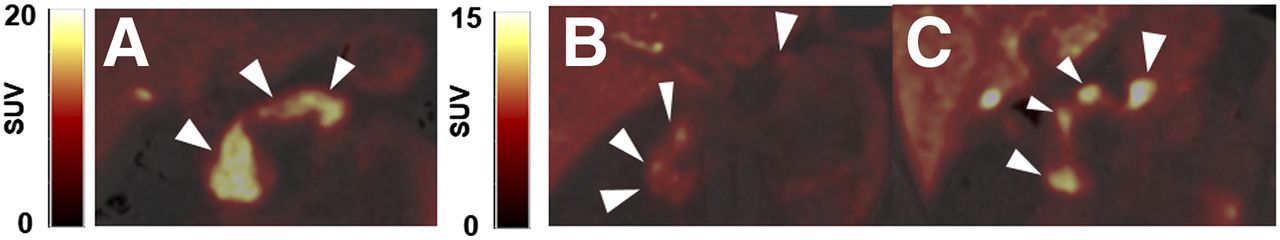
Representative axial slices of PET/CT overlay of pancreas uptake (arrowheads) for each dopaminergic radioligand 11C-(+)-PHNO (A), 11C-FLB457 (B), and 11C-raclopride (C). All SUV images summed from 20 to 30 min.

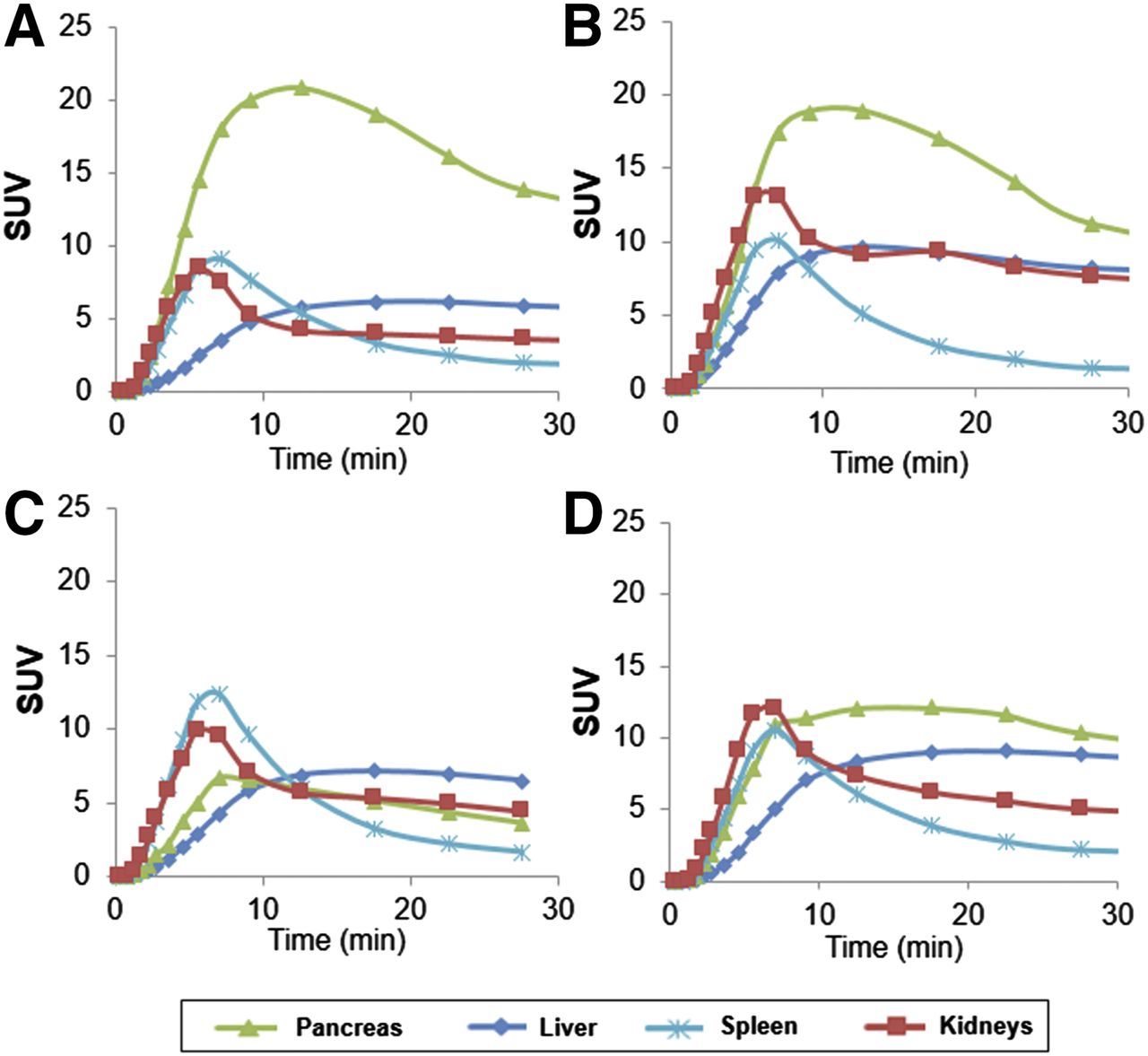
Mean time–activity curves of pancreas, liver, spleen, and kidneys of 2 HCs for each dopaminergic radioligand 11C-(+)-PHNO (A), 11C-FLB457 (B), and 11C-raclopride (C).
Low pancreas SUV and SUVR were seen in all other radioligands compared with 11C-(+)-PHNO (Table 3) as seen in representative PET images (Supplemental Fig. 1; supplemental materials are available at http://jnm.snmjournals.org) and time–activity curves (Supplemental Fig. 2). Time–activity curves (0–90 min) showed no contrast in the pancreas compared with reference organ (spleen) beyond 30 min for any radioligand.
Pancreatic SUV (20–30 Minutes) of Each Radioligand in HCs (SUVR, Spleen Reference)
11C-(+)-PHNO pancreas SUV was 13.8 ± 1.7 in HCs. A 71% reduction (SUV, 4.0) was seen in the T1DM subject with undetectable C-peptide levels. In a T1DM subject with a fasting C-peptide level of 0.38 ng/mL, a 20% reduction (SUV, 11.0) was seen (Fig. 3; Table 4). Comparisons between time–activity curves of the C-peptide–deficient T1DM subject demonstrated substantially reduced uptake and contrast to background organs compared with the HCs (Fig. 4). SUV in other abdominal organs did not show differences between HC and T1DM (Table 5), suggesting that the 11C-(+)-PHNO SUV differences are specific to changes in the pancreas.
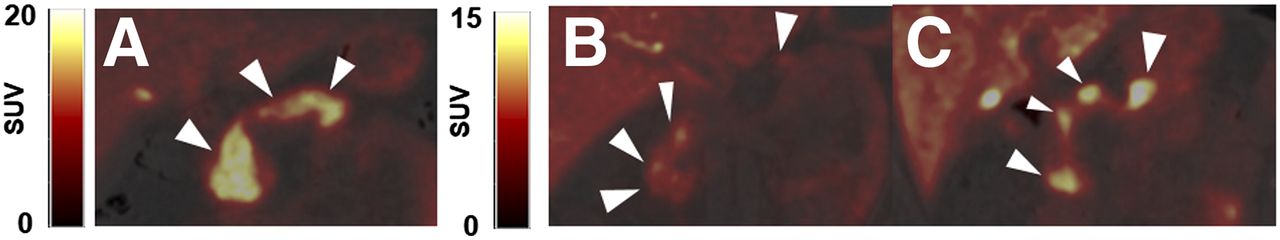
Representative coronal PET/CT images of 11C-(+)-PHNO in pancreas (arrowheads) for HC (A), C-peptide–deficient T1DM subject (B), and T1DM subject with detectable C-peptide (C). All SUV images are summed from 20 to 30 min. (A) SUV scale 0–20. (B and C) SUV scale 0–15.
SUV (20–30 Minutes) of 11C-(+)-PHNO in Pancreas, Spleen, Kidneys, and Liver
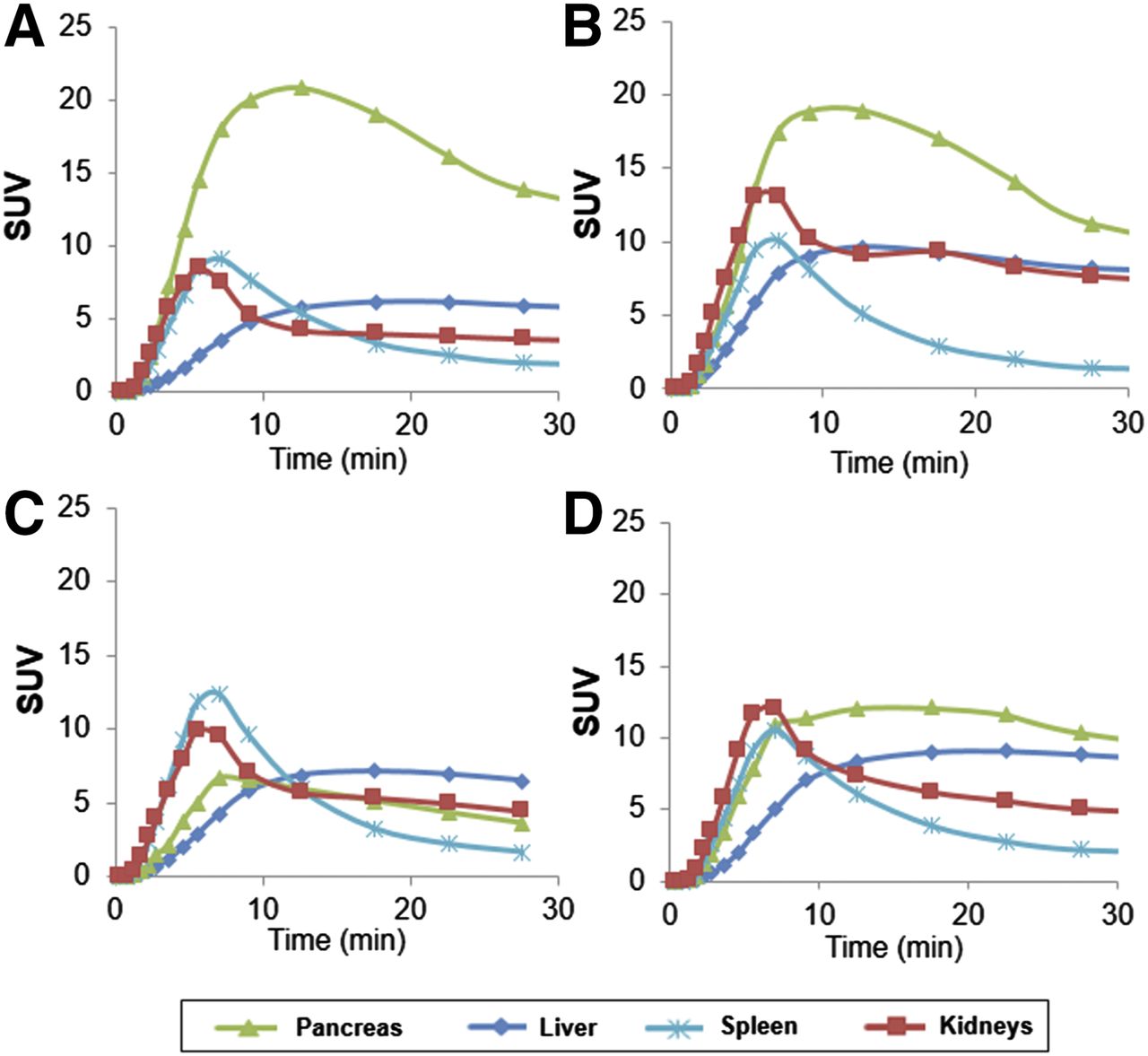
11C-(+)-PHNO time–activity curves of pancreas, liver, spleen, and kidneys for HCs (A and B), C-peptide–deficient T1DM (C), and T1DM subject with detectable C-peptide (D).
SUVR-1 and BPND in Baseline and Blocking Scans, and Percentage Reduction from Baseline in Pancreas and Brain Regions in NHPs with 11C-(+)-PHNO
Assessing the D2 and D3 components of 11C-(+)-PHNO–specific binding, SUVR-1 and BPND of 11C-(+)-PHNO in NHP was reduced by a D3 receptor antagonist in the pancreas, SN, putamen, and caudate (Table 5; Supplemental Fig. 3). SUVR-1 and BPND for the D2 receptor antagonist demonstrated a smaller reduction than the D3 receptor antagonist in the pancreas and SN and larger reductions in the putamen and caudate (Table 5). When a brain-only study was performed, BPND demonstrated reductions in the SN, putamen, and caudate similar to SUVR-1 in the previous D2 antagonist multiorgan scan (Table 5; Supplemental Fig. 4), suggesting that SUVR-1 is a valid measure for assessing blockade.
The SN can be considered approximately 100% D3-selective, and the putamen and caudate about 100% D2-selective. Because the pancreas blocking percentages fell between the values for these regions, this suggests that 11C-PHNO binds to both D2 and D3 in the pancreas.
DISCUSSION
We identified 6 radioligands through use of a β-cell gene atlas that were subsequently screened using PET/CT in HCs for high pancreas uptake and high contrast-to-background abdominal organs, similar to a previous study (12). The D3-preferring radioligand 11C-(+)-PHNO showed the best potential as a BCM-imaging agent, based on high pancreatic uptake in HCs, lower uptake in T1DM subjects, and demonstrable specific binding with NHP blocking studies.
Of the 6 radioligands studied, 3, 11C-UCB-J (synaptic vesicle protein 2A), 11C-DASB (serotonin transporter), and 11C-AS2471907 (11β-HSD1 enzyme inhibitor), did not demonstrate high pancreas signal or contrast-to -background abdominal organs (Table 3). Synaptic vesicle proteins are associated with insulin-containing granules in neuroendocrine cells (30,31); however, evoked basal hormone release was attenuated more by silencing SV2C than SV2A in neuroendocrine cells, possibly explaining low SV2A signal (31). The main mechanism of 11β-HSD1 inhibition is lowering intracellular cortisol concentrations in the liver and adipose tissue, and liver uptake was more than 25 times that of the pancreas, making 11C-AS2471907 undesirable for BCM imaging (32). Serotonin mechanisms have been implicated in the endocrine pancreas (10,33); however, given low uptake and contrast in the pancreas, 11C-DASB may not be a suitable BCM imaging agent.
In agreement with studies suggesting the feasibility of targeting DRs for imaging BCM (15,16,19), promising results were obtained for 1 of the 3 DR radioligands. 11C-(+)-PHNO demonstrated the greatest uptake and contrast compared with background organs in HCs. In contrast, a large reduction in SUV (71%) in the pancreas of a subject with T1DM without detectable C-peptide (Table 4) suggests possible specific binding of 11C-(+)-PHNO to islet β-cells. Intriguingly, a T1DM subject with detectable levels of insulin production had higher pancreas SUV than the first T1DM subject (Table 3), suggestive of intermediate BCM levels.
Using the 3 dopaminergic radioligands in our study, we can indirectly infer mechanisms of both D2 and D3 receptors. On the basis of preliminary success with D2 receptor radioligands to differentiate healthy and diabetic rats (15,16), and the β-cell gene atlas data (Table 2), D2 receptors may exist throughout the endocrine and exocrine pancreas. D2 receptors, as measured by 11C-(+)-PHNO, may not be directly or linearly correlated with β-cells; therefore, information obtained by this imaging technique should be viewed with caution. Further study is required to assess whether reduced D3 receptors in T1DM are specific to human β-cells.
11C-raclopride and 11C-FLB457 are both antagonists, and thus have equal affinity for high- and low-affinity receptor sites; 11C-FLB457 has higher affinity (29,34). 11C-(+)-PHNO is an agonist that binds to D2 and D3 receptors with a 25- to 50-fold-higher affinity for D3 receptors. Quelch et al., reported higher specific binding with [3H](+)PHNO versus [3H]raclopride in microsomal but not extracellular or cytosolic tissue in the pig striatum (35). Similar D2/3 receptor internalization may occur in the pancreas, which may explain the higher binding of 11C-(+)-PHNO than 11C-raclopride or 11C-FLB457.
To assess the D2 and D3 components of pancreatic 11C-(+)-PHNO binding, we performed blocking experiments in NHP. The brain occupancy data for the D3 antagonist agreed well with previous 11C-(+)-PHNO data (36). For each blocking study (Table 5), using the receptor occupancy values in pancreas (rpanc), putamen (rput), and SN (rSN), and assuming that SN and putamen are 100% D3- and D2-selective, respectively, we can estimate the fraction of D3 ( and D2 (1-
and D2 (1- binding in the pancreas (the supplemental materials provide a derivation of the equation):
binding in the pancreas (the supplemental materials provide a derivation of the equation): Eq. 1In the D2 antagonist–blocking study, fD3 is 0.77 using SUVR-1 or BPND and in the D3-blocking study, fD3 is 0.58 (SUVR-1) or 0.43 (BPND). Averaging suggests that specific binding in the pancreas is about two-thirds D3 and one-third D2. Note that because specific binding measurements at tracer levels represent receptor density (Bmax)/affinity (Kd), not Bmax, this does not imply that two-thirds of the D2/3 receptors are D3. Rather, if the relative D2/D3 affinities for 11C-(+)-PHNO in the pancreas are the same as those in the brain, that is, a 25- to 50-fold-higher affinity for D3 (22), then the Bmax for D2 is in fact much higher than that of D3.
Eq. 1In the D2 antagonist–blocking study, fD3 is 0.77 using SUVR-1 or BPND and in the D3-blocking study, fD3 is 0.58 (SUVR-1) or 0.43 (BPND). Averaging suggests that specific binding in the pancreas is about two-thirds D3 and one-third D2. Note that because specific binding measurements at tracer levels represent receptor density (Bmax)/affinity (Kd), not Bmax, this does not imply that two-thirds of the D2/3 receptors are D3. Rather, if the relative D2/D3 affinities for 11C-(+)-PHNO in the pancreas are the same as those in the brain, that is, a 25- to 50-fold-higher affinity for D3 (22), then the Bmax for D2 is in fact much higher than that of D3.
The connection between dopaminergic activity and the endocrine pancreas has been known for 40 y (37,38). It is believed that circulating dopamine is generally below levels needed to activate peripheral DRs (39). One current theory proposes that circulating L-DOPA (dopamine precursor) is taken up by β-cells, converted to dopamine, and cosecreted with insulin as an autocrine signal by binding to DRs on the surface of β-cells (19). Dopamine antagonists have been shown to cause hyperinsulinemia in healthy subjects and are associated with diabetes in psychiatric patients, versus agonists such as bromocriptine, which improved glycemic control (40). Interestingly, L-DOPA, a common treatment in Parkinson disease, has demonstrated reduced insulin secretion in such patients and mice (40). Combined, this evidence suggests dopamine may be an integral part of the endocrine pancreas.
Several studies have attempted to elucidate more specific mechanisms of DRs in the endocrine pancreas. In vitro studies using mouse islet cells have indicated that a D3 receptor antagonist increases insulin secretion, suggesting that dopamine binding to the D3 receptor inhibits insulin secretion (18). In contrast, Chen et al. found no evidence of pancreatic D3 receptors in rats using Western blot and immunofluorescence techniques, and concluded D1 receptors were colocalized with insulin staining in rat Islet cells (41). Additional studies have indicated the D2 receptor may be involved in insulin secretion inhibition (42–45). Several other studies have demonstrated inhibitory actions of dopamine on insulin release in preclinical models but did not assess whether specific receptors were responsible for such actions (37,46–49). It is important to note that the structure and function of proteins may differ between species such as mouse, rat, and in vitro cultures of insulinoma cell lines, such as INS-1 cells. The human β-cell gene expression atlas we used demonstrated a specificity of the D3 receptor gene to β-cells and lack of expression in the exocrine and other endocrine cells in the islets of Langerhans (Table 2) (20). Further studies are warranted to determine the specificity of different DR subtypes in the endocrine pancreas and β-cells. However, from this evidence and the large reduction seen in the T1DM subject with undetectable C-peptide, we hypothesize that high-affinity D3 receptors may be specific to the endocrine pancreas and perhaps to β-cells.
Several potential limitations in the current study must be assessed. The use of spleen as a reference region has been examined for the VMAT2 tracer 18F-FP-(+)-DTBZ (13) and we used SUVR in the current study. Several issues must be evaluated to validate this choice. First, the presence of D2/D3 receptors in the spleen must be assessed; a preliminary study has demonstrated that all 5 DR subtypes, along with VMAT1 and VMAT2, are present to some extent in the spleen (50). In NHP blocking studies, we saw 37% and 13% reductions in spleen SUV during D2 and D3 antagonist studies, respectively; however, no differences were seen between HC and T1DM subjects in spleen SUV. Second, it remains to be validated if nondisplaceable binding in the spleen is equal to the pancreas, or if a scale-factor correction is needed, as with the VMAT2 tracer (13). Further, radiolabeled metabolites may accumulate in the spleen; therefore, we focused on early data (20–30 min) for the patient comparisons, to maximize specific binding while simultaneously minimizing potential biases due to radiolabeled metabolites. Arterial sampling of HC and T1DM subjects should be performed to characterize 11C-(+)-PHNO metabolites and assess whether there are between-group differences that may affect interpretation of results, similar to a previous study with enantiomers of 18F-FP-DTBZ (13).
CONCLUSION
We have examined 6 PET brain radioligands in the pancreas in both HC and T1DM subjects and have demonstrated that 11C-(+)-PHNO, a D3-preferring receptor agonist, may be a potential marker of BCM. NHP blocking studies suggest that about two-thirds of pancreatic binding originates from D3 receptors. Further in vitro and in vivo studies to establish D3 receptor specificity to β-cells are warranted to develop 11C-(+)-PHNO as a candidate for clinical measurement of BCM.
DISCLOSURE
This study was supported by the NIH grant 1DP3DK104092-01 and was also made possible by 1S10OD010322-01 and by CTSA grant number UL1 TR000142 from the National Center for Advancing Translational Sciences (NCATS), a component of the NIH. Its contents are solely the responsibility of the authors and do not necessarily represent the official view of the NIH. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
The authors appreciate the excellent technical assistance of the Yale PET Center staff.
Footnotes
Published online Jan. 25, 2018.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication June 9, 2017.
- Accepted for publication December 7, 2017.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A metabolically stable PET tracer for imaging synaptic vesicle protein 2A: Synthesis and preclinical characterization of [18F]SDM-16
- Optoacoustic Imaging of Glucagon-like Peptide-1 Receptor with a Near-Infrared Exendin-4 Analog
- PET Imaging of Pancreatic Dopamine D2 and D3 Receptor Density with 11C-(+)-PHNO in Type 1 Diabetes
- Relative Pancreas Volume Is Reduced in First-Degree Relatives of Patients With Type 1 Diabetes