


Abstract
Multidrug resistance (MDR) is a major challenge to the successful treatment of acute myeloid leukemia (AML). Our purpose was to determine whether 111In-HuM195 anti-CD33 antibodies modified with peptides harboring nuclear localizing sequences (NLS) could kill drug-resistant AML cell lines and primary AML patient specimens expressing MDR transporters through the emission of Auger electrons. Methods: HuM195, M195, and irrelevant mouse IgG (mIgG) were conjugated to 10 ± 3 NLS peptides and then labeled with 111In by diethylenetriaminepentaacetic acid substitution to a specific activity of 1–8 MBq/μg. The binding affinity of HuM195 and M195 was determined for HL-60 and mitoxantrone-resistant HL-60-MX-1 cells. Nuclear localization of 111In-NLS-HuM195, 111In-NLS-M195, 111In-HuM195, and 111In-M195 was measured by subcellular fractionation. The antiproliferative effects of 111In-NLS-HuM195, 111In-NLS-M195, 111In-HuM195, and 111In-M195 (2.5–250 kBq/well) on HL-60 and HL-60-MX-1 were studied using the WST-1 assay. Clonogenic survival of HL-60 and HL-60-MX-1 leukemic cells and 10 primary AML specimens with MDR phenotype (assessed by flow cytometry) was determined after exposure for 3 h at 37°C to 2.5–250 mBq/cell of 111In-NLS-HuM195, 111In-HuM195, or 111In-NLS-mIgG. Clonogenic survival versus the amount of radioactivity incubated with the cells (mBq/cell) was plotted, and the mean lethal amount of radioactivity and the lower asymptote of the curve (plateau) were determined. Results: The 111In-labeled anti-CD33 monoclonal antibodies HuM195 and M195 modified with NLS were efficiently routed to the nucleus of HL-60 cells and their mitoxantrone-resistant clone after CD33-mediated internalization. The following are the principal findings of our study: 111In-NLS-HuM195 was more effective at killing HL-60 and HL-60-MX-1 cells than was 111In-HuM195, a strong correlation between the specific activity of the 111In-labeled radioimmunoconjugates and their cytotoxicity toward AML cells existed, and leukemic cells from patients were killed by 111In-NLS-M195 or 111In-M195, but the cytotoxic response among specimens was heterogeneous. Conclusion: NLS conjugation enhanced the nuclear uptake and cytotoxicity of 111In-HuM195 and 111In-M195 toward drug-resistant AML cell lines as well as patient specimens expressing a diversity of MDR phenotypes, including Pgp-170, BCRP1, or MRP1 transporters. Targeted Auger electron radioimmunotherapy using 111In-labeled anti-CD33 monoclonal antibodies modified with NLS may be able to overcome MDR and provide a means of treating chemotherapy-resistant myeloid leukemias in patients.
Acute myeloid leukemia (AML) is a clonal malignancy of the hematopoietic system characterized by accumulation of immature cell populations in the bone marrow or peripheral blood (1). AML is the most common type of leukemia in adults but has the lowest survival rate of all leukemias (2). Induction chemotherapy with doxorubicin, idarubicin, etoposide, and mitoxantrone induces complete remission in 65%–70% of adults with de novo AML (3–5). However, despite these intensive treatments, patients relapse because of the development of multidrug resistance (MDR) (1). Extrusion of drugs by membrane adenosine triphosphate (ATP)–dependant drug efflux pumps (ATP-binding cassette [ABC] transporters) is commonly implicated in MDR. The best characterized ABC transporter is Pgp-170, encoded by the MDR1 gene, which has been associated with poor treatment outcome in AML (6). Other drug efflux pumps include MRP1, which also contributes to MDR in AML, and MRP2-MRP9 and BCRP1, which are still under investigation and have not yet been proven to play a role in MDR in AML (1,5).
Immunotherapeutic strategies could potentially circumvent MDR and yield remissions in drug-resistant AML. One promising target on myeloid leukemia cells is CD33, a 67-kDa sialoadhesion glycoprotein that is expressed in more than 90% of cases of AML (7). CD33 is also displayed by normal myeloid cells but is not present on nonhematopoietic tissues or the earliest pluripotent hematopoietic progenitor cells (7,8). This restricted distribution of CD33 has been exploited for immunotherapy of AML using the humanized anti-CD33 monoclonal antibody (mAb) HuM195 (Lintuzumab; Protein Design Labs) or the anti-CD33 mAb calicheamicin immunoconjugates ([gemtuzumab ozogamicin] Mylotarg; Wyeth). Unfortunately, HuM195 was found to have limited efficacy as a single agent and did not improve survival when combined with chemotherapy in a phase III clinical trial (9). An overall 30% response rate to gemtuzumab ozogamicin has been achieved in patients with AML (10), but calicheamicin is a substrate for Pgp-170 and is unable to kill Pgp-170–expressing leukemia cells (11).
Radioimmunotherapy (RIT) of AML using 131I- and 90Y-labeled anti-CD33 mAbs M195 and HuM195 dramatically decreased leukemic cells in the blood and bone marrow of patients, but these radioimmunoconjugates (RICs) were associated with severe myelosuppression and hepatotoxicity (12). These normal-tissue toxicities are thought to be partially due to irradiation of nontargeted cells by the long-range (2–10 mm) β-particles emitted by 131I or 90Y (“cross-fire” effect) (13). The Auger electron–emitting radionuclide 111In is an attractive alternative for labeling HuM195 for RIT of myeloid leukemias. More than 99% of the Auger electrons emitted by 111In have a range of less than 1 μm, which obviates the cross-fire effect and yields high-linear-energy transfer comparable to that of highly cytotoxic α-emitters (i.e., 50–100 keV/μm). We previously reported that HuM195 labeled with 111In and modified with 13-mer peptides (CGYGPKKKRKVGG) harboring the nuclear localization sequence (NLS; underlined) of SV-40 large T-antigen was internalized and routed to the nucleus of leukemia cells, in which the Auger electrons were lethal to HL-60 myeloid leukemia cells and primary AML specimens (14). 111In-NLS-HuM195 was not toxic to normal tissues in BALB/c mice at doses up to 185 MBq/kg. Our hypothesis in the current study was that mitoxantrone-insensitive HL-60-MX-1 cells and primary AML specimens expressing ABC transporters would be sensitive to the cytotoxic effects of 111In-labeled M195 and HuM195 mAbs modified with NLS peptides. Thus, RIT using 111In-NLS-HuM195 could potentially offer an opportunity to treat patients with AML who have evidence of MDR at presentation or who have relapsed after induction chemotherapy due to the emergence of MDR.
MATERIALS AND METHODS
Human AML Cells and Primary AML Specimens
HL-60 (CCL-240) and mitoxantrone-resistant HL-60-MX-1 (CRL-2258) human leukemia cells, as well as M195 hybridoma cells (HB-10306) secreting M195 mAbs, were purchased from the American Type Culture Collection. We determined that the concentration of mitoxantrone required to inhibit the growth of HL-60-MX-1 cells by 50% (EC50) is almost 5-fold greater than that for HL-60 cells (EC50 = 1.11 vs. 0.26 nM, respectively). All cells were cultured at 37°C and in a 5% CO2 atmosphere in growth medium (RPMI 1640 medium; Sigma) supplemented with 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 4.5 g of d-glucose per liter, 1 mM sodium pyruvate, 1% (v/v) penicillin/streptomycin (Sigma), and 10% (v/v) non–heat-inactivated fetal calf serum (FCS) (Sigma). AML specimens were collected under a protocol (no. 02-0763C) approved by the Human Subjects Ethics Review Board at the Princess Margaret Hospital, University Health Network, Toronto, Ontario. Mononuclear blood cells were isolated by Ficoll-Paque (Stem Cell Technologies) density-gradient centrifugation and cryopreserved in growth medium containing 50% (v/v) FCS and 5% (v/v) dimethyl sulfoxide (Sigma) until needed.
Special cell culture conditions using complete Methocult-4800 medium (Stem Cell Technologies) with or without supplementation with recombinant growth factors were used for the clonogenic assays of HL-60 and HL-60-MX-1 cells, as well as for primary AML specimens exposed to 111In-labeled anti-CD33 mAbs. Complete Methocult medium was prepared by mixing 40 mL of Methocult-4800 with 30 mL of FCS, 10 mL of a 10% (w/v) bovine serum albumin solution (Sigma), 704 μL of 14.2 mM 2-mercaptoethanol (Sigma), 1 mL of 200 mM l-glutamine (Sigma), 1 mL of 100× antibiotics solution (product P0781; Sigma), and 18 mL of Iscove's modified Dulbecco's medium (product I3390; Sigma). For primary AML specimens, complete Methocult medium was supplemented with recombinant human growth factors obtained from Peprotech. The final concentrations were 50 ng/mL for stem cell factor, 20 ng/mL for GM-CSF, 20 ng/mL for IL-3, 20 ng/mL for IL-6, 20 ng/mL for G-CSF, and 50 ng/mL for Flt-3 ligand.
111In-NLS-CD33 mAbs
Humanized antihuman-CD33 HuM195 mAbs (IgG1) were provided by Protein Design Labs Inc. Mouse IgG (mIgG) was purchased from Sigma (product I5381). Anti-CD33 M195 murine mAbs were isolated from the culture medium of M195 hybridoma cells by affinity chromatography (batch method) using Protein A Sepharose (Sigma). Sodium dodecylsulfonate polyacrylamide gel electrophoresis on a 4%–20% Tris-HCl–gradient gel under nonreducing conditions and Western blot analysis probed with antihuman-CD33 (eBioscience) showed a single band for M195 corresponding to an Mr of 150 kDa. Conjugation of mAbs with diethylenetriaminepentaacetic acid (DTPA) dianhydride for labeling with 111In was performed before NLS modification to prevent derivatization of ε-amino groups of lysines on the NLS peptides—these residues are critical for nuclear importation (15). Briefly, HuM195, M195, or mIgG (100 μL of a 5 mg/mL solution in 100 mM sodium bicarbonate, pH 8.2) were reacted with a 10-fold molar excess of DTPA dianhydride (5 μL of a 6.7 mM solution in anhydrous dimethyl sulfoxide; Sigma) for 1 h at room temperature. Excess DTPA was removed by size-exclusion chromatography on a Sepharose G50 (Sigma) minicolumn eluted with phosphate-buffered saline (PBS), pH 7.6. Purified DTPA-conjugated antibodies were reconcentrated to 2 mg/mL on a Microcon YM-100 device (Millipore). DTPA substitution was 1.8 ± 0.5 mol of DTPA per mole of antibodies (n = 15), measured as previously reported (16).
NLS peptides were conjugated to DTPA-HuM195, M195, or mIgG mAbs (14) by introducing maleimide groups through a reaction of 150 μL of antibodies (2 mg/mL) with a 30-fold molar excess of sulfosuccinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (sulfo-SMCC) (Pierce) or 50 μL of a 1.2 mM solution in PBS (pH 7.6) for 1 h at room temperature. Excess sulfo-SMCC was removed by size-exclusion chromatography on a Sepharose G50 minicolumn eluted with PBS, pH 7.0. Maleimide-derivatized DTPA antibodies (100 μL of a 2 mg/mL solution in PBS, pH 7.0) were reacted with a 25-fold molar excess (50 μL of a 0.67 mM solution in PBS, pH 7.0) of 13-mer peptides (CGYGPKKKRKVGG) harboring the NLS of SV-40 large T-antigen (underlined) (Advanced Protein Technology Center, Hospital for Sick Children) for 18 h at 4°C. Excess NLS peptides were removed by size-exclusion chromatography on a Sepharose G50 minicolumn eluted with 100 mM sodium acetate, pH 6.0. NLS-conjugated antibodies were finally reconcentrated to 2 mg/mL. The level of NLS substitution was 10 ± 3 NLS peptides per antibody molecule measured as previously reported (14). NLS-conjugated or unmodified DTPA-HuM195, DTPA-M195, or DTPA-mIgG was radiolabeled by incubation of 10–200 μg of mAbs with 111InCl3 (MDS Nordion) for 60 min at room temperature. The radiochemical purity was 98.2% ± 4.5% (n = 75), determined by instant thin-layer silica gel chromatography and size-exclusion high-performance liquid chromatography as previously reported (14), and the final specific activity (SA) was 1–8 MBq/μg. HuM195 or M195 mAbs were labeled with 123I using the IODO-GEN method (17).
Flow Cytometry of Leukemia Cells
The immunoreactivities of HL-60, HL-60-MX-1, and AML patient specimens were assessed with mouse antibodies specific for human CD33 (anti-CD33) conjugated to fluorescein isothiocyanate or phycoerythrin-cyanine-5 (PE-cy5) and with anti-CD243-fluorescein isothiocyanate (anti-Pgp-170), anti-BCRP1-PE (anti-ABCG2), and anti-MRP1-fluorescein isothiocyanate (Becton-Dickinson Pharmingen). Nonspecific binding was also assessed with isotype-matched irrelevant antibody controls.
Radioligand Saturation Binding Assays
The CD33 binding affinities (Kd) and maximum binding sites (Bmax) for 111In-labeled NLS-HuM195 and NLS-M195 and 123I-labeled HuM195 and M195 were determined in direct radioligand saturation binding assays using HL-60 and HL-60-MX-1 cells as previously reported (14). The bound radioligand concentration (mol/L) was plotted versus total radioligand concentration (mol/L), and the resulting curve was fitted to a 1-site saturation binding model using Prism software (version 4.0; GraphPad). The dissociation constant (Kd) and Bmax were estimated. All data points and radioligand binding experiments were performed in triplicate.
Measurement of Nuclear Localization
The nuclear localization of 111In-labeled HuM195, M195, NLS-HuM195, and NLS-M195 in HL-60 and HL-60-MX-1 cells was measured by subcellular fractionation. Briefly, 111In-labeled mAbs (200 nM in 100 μL of growth medium) were incubated for 3 h at 37°C with 1 × 106 HL-60 or HL-60-MX-1 cells in microtubes. The cell suspensions were then recovered and centrifuged for 5 min at 500g, the supernatant was removed, and the cell pellet was resuspended in ice-cold PBS, pH 7.4. This procedure was repeated 3 times. Then 500 μL of cell membrane lysis buffer (containing 1.25 mL of potassium chloride [3.73 g/50 mL], 250 μL of magnesium chloride [10.83 g/50 mL], 333 μL of Tris-HCl [1 g/10 mL], and 250 μL of NP-40) was added, and the cells were incubated for 6 min at 4°C. Finally, the cell suspension was centrifuged for 5 min at 1,000g, the cytoplasmic fraction (supernatant) was removed, and the nuclear pellet was isolated and resuspended in ice-cold PBS, pH 7.4. The nuclear pellet was rinsed twice with ice-cold PBS (pH 7.4), the suspension was recentrifuged, and the supernatant was removed and combined with that previously separated. Finally, the radioactivities contained in the cytoplasmic and nuclear fractions were measured in a γ-counter. All data points and individual experiments were performed in triplicate.
Cell Proliferation Assays
The antiproliferative effects of 111In-HuM195, 111In-M195, 111In-NLS-HuM195, or 111In-NLS-M195 on HL-60 and HL-60-MX-1 cells were measured by the WST-1 cell-viability assay (Roche Diagnostics). Approximately 1 × 103 HL-60 or HL-60-MX-1 cells were suspended in 100 μL of growth medium and seeded in replicates (n = 5) into wells in a 96-well culture plate. Antiproliferative effects were tested at 3 different specific activities (1, 2, or 5 MBq/μg). Control treatments included 111In-NLS-mIgG (5 MBq/μg) and 111In-acetate as well as nonradiolabeled NLS peptides, DTPA-NLS-HuM195, or DTPA-NLS-M195. For the nonradioactive treatments, mass-equivalent concentrations identical to the highest tested for the radiolabeled mAbs (i.e., lowest specific activity) or equivalent NLS-peptide concentrations for these 111In-labeled antibodies were used. Increasing concentrations (0–250 kBq/well) of 111In-labeled mAbs in 100 μL of growth medium were added to each well, and the cells were cultured for 7 d. WST-1 reagent (20 μL) was then added, and the dishes were incubated at 37°C in 5% CO2 for 2 h. The absorbance of the wells was measured at 450 and 690 nm (reference wavelength) in a microplate reader (model Elx800; Bio-Tek Instruments Inc.). Growth inhibition was defined as the ratio of the absorbance of wells containing treated cells to the absorbance of wells containing untreated cells. This ratio was plotted versus the concentration (nmol/L) of 111In-labeled mAbs, and the resulting curve was analyzed by nonlinear top-to-bottom regression using Prism (version 4.0). The concentrations required to inhibit cell growth by 50% (EC50) were calculated.
Clonogenic Assays
The clonogenic survival of HL-60 or HL-60-MX cells exposed to 111In-M195, 111In-NLS-HuM195, 111In-NLS-M195, or 111In-NLS-mIgG (irrelevant control) was determined. Briefly, 1 × 106 cells were incubated for 3 h at 37°C in Eppendorf microtubes with 111In-labeled mAbs (0–250 mBq/cell; specific activities of 1, 3, 6, or 8 MBq/μg) contained in 60 μL of growth medium. An additional 940 μL of growth medium were then added, and the cell suspensions were centrifuged for 5 min at 500g. This was repeated twice. Finally, the cells were resuspended in growth medium to a concentration of 4–5 × 103 cells/mL, and approximately 400 μL were added to 4 mL of complete Methocult. Approximately 1 mL of this mixture was plated in triplicate into 35-mm cell culture dishes (Stem Cell Technologies). The cells were cultured at 37°C in 5% CO2 for 14 d, and the number of colonies (>100 cells) formed was counted using an inverted microscope. All experiments were performed in triplicate. Clonogenic survival was calculated by dividing the number of colonies in treated dishes by the number in untreated dishes and multiplying by 100%. Clonogenic survival versus the amount of radioactivity incubated with the cells (mBq/cell) was plotted and the resulting curve fitted to a dose-response curve, N = ([N0 − plateau] × exp[−D/D0] + plateau), where N is the number of cells surviving, N0 is the number of cells initially, D is the amount of radioactivity incubated with the cells (mBq/cell), D0 is the mean lethal amount of radioactivity (mBq/cell), and plateau is the lower asymptote of the curve (18).
The clonogenic survival of primary AML specimens exposed to 111In-labeled mAbs was determined by first thawing the specimens gradually over 10 min at 37°C and then rinsing the cells with Hanks' balanced salt solution (Stem Cell Technologies) containing 5% FCS. The cells were subsequently resuspended in growth medium, and 5 × 105 cells were incubated with 0–250 mBq/cell of 111In-M195 or 111In-NLS-M195 (8 MBq/μg) or 111In-NLS-mIgG (8 MBq/μg) in 60 μL of growth medium in microtubes for 3 h at 37°C. The cells were recovered and centrifuged at 500g. Approximately 2 × 105 cells were resuspended in 1 mL of complete Methocult medium supplemented with growth factors and plated in triplicate into 35-mm cell culture dishes. The cells were cultured at 37°C in 5% CO2 for 14 d, and the number of colonies formed (>100 cells) was counted using an inverted microscope. The clonogenic survival was calculated, and the data were analyzed by Prism (version 4.0) as described previously.
Statistical Analysis
Statistical significance was tested using the Mann–Whitney U test (P < 0.05) or ANOVA with the Bonferroni adjustment (P < 0.05).
RESULTS
Flow Cytometry of Leukemia Cells
CD33 was highly expressed by all 10 primary AML specimens, with 79%–99% of the cells immunopositive (Table 1). Similarly, almost 100% of HL-60 and HL-60-MX-1 cells displayed CD33 epitopes. No expression of the selected MDR transporters was found on HL-60 and HL-60-MX-1 cells. The resistance of HL-60-MX-1 cells to mitoxantrone is atypical and is due to the absence of the topoisomerase IIb isoform (19). In contrast, primary AML specimens exhibited immunoreactivity for Pgp-170 (8%–77%), BCRP1 (0.1%–14%), and MRP1 (1%–73%).
Expression of CD33 and MDR Transporter Epitopes on Primary AML Specimens
Radioligand Saturation Binding Assays
NLS conjugation and 111In-labeling of M195 resulted in a 3-fold increased Kd for binding to HL-60 and HL-60-MX-1 cells (13.9 ± 1.1 and 42.7 ± 3.5 nmol/L, respectively), compared with 123I-M195 (Kd, 4.1 ± 0.4 and 14.3 ± 1.6 nmol/L, respectively). Modification of HuM195 with NLS and labeling with 111In did not significantly change the Kd for binding CD33 on HL-60 or HL-60-MX-1 cells (1.2 ± 0.4 vs. 1.7 ± 0.6 nmol/L, respectively; P > 0.05) but was 2- to 3-fold higher than that for 123I-HuM195 (0.5 ± 0.2 and 0.8 ± 0.4 nmol/L, respectively). No significant differences in Bmax values for binding of 123I-M195, 123I-HuM915, 111In-NLS-M195, or 111In-HuM195 to HL-60 or HL-60-MX-1 cells (3–4 × 104 CD33 epitopes per cell) were exhibited.
Measurement of Nuclear Localization
The percentage of radioactivity imported into the nucleus for 111In-M195 and 111In-NLS-M195 was 4.4 ± 0.1 versus 22.4 ± 6.3 (P < 0.05) in HL-60 cells and 5.0 ± 1.4 versus 18.5 ± 4.4 (P < 0.05) in HL-60-MX-1 cells. The percentage of nuclear radioactivity for 111In-HuM195 and 111In-NLS-HuM195 was 5.8 ± 0.6 versus 24.1 ± 3.2 (P < 0.05) in HL-60 cells and 5.1 ± 0.8 versus 19.7 ± 3.4 (P < 0.05) in HL-60-MX-1 cells. These results were consistent with those previously reported for HL-60 cells incubated with 111In-HuM195 modified with 4–8 NLS peptides (14). No significant differences in the proportion of nuclear localization of radioactivity between HL-60 and HL-60-MX-1 cells for 111In-NLS-M195 or 111In-NLS-HuM195 were demonstrated.
Cell Proliferation Assays
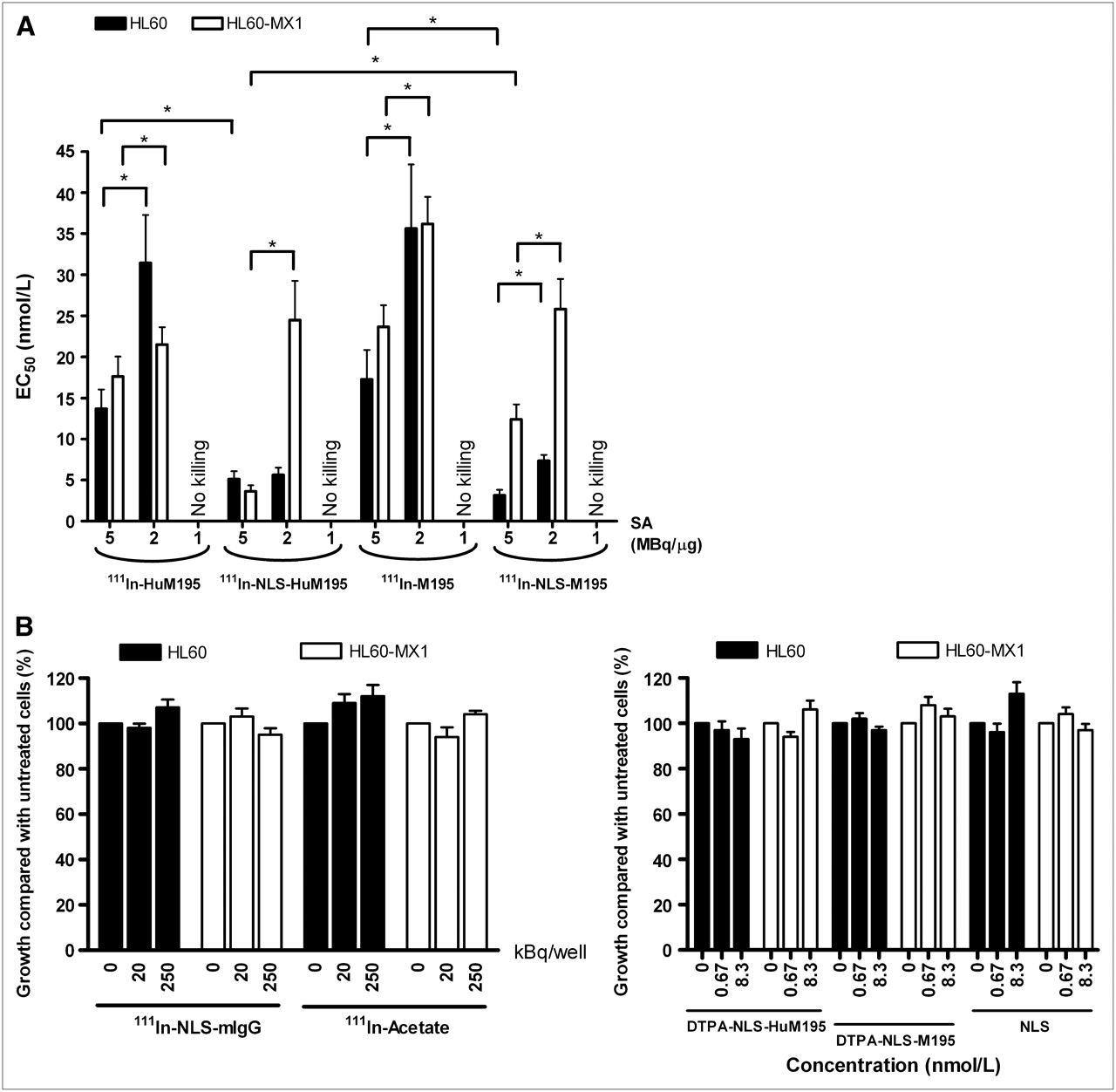
Growth inhibition of HL-60 and HL-60-MX-1 cells by the 111In-labeled mAbs with or without NLS modification was greater for an SA of 5 MBq/μg than for that of 2 MBq/μg (Fig. 1A). No growth inhibition was found at an SA of 1 MBq/μg. At the highest SA tested, NLS incorporation resulted in significantly lower EC50 values, compared with unmodified 111In-HuM195 and 111In-M195 mAbs. No significant differences between 111In-NLS-HuM195 and 111In-NLS-M195 in the ability to inhibit the growth of HL-60 cells were exhibited. HL-60-MX-1 cells, however, were as sensitive as HL-60 cells to 111In-NLS-HuM195 but were significantly less growth-inhibited by the murine antibody form 111In-NLS-M195. At the highest concentration (1.7 nmol/L) and SA tested (5 MBq/μg), the growth of HL-60 and HL-60-MX-1 cells were reduced to 3.2% ± 1.1% and 5.1% ± 2.4%, respectively, by 111In-NLS-HuM195 and to 1.8% ± 1.4% and 2.7% ± 1.6%, respectively, by 111In-NLS-M195. Neither cell line was affected by exposure to 111In-NLS-mIgG or 111In-acetate or to nonradiolabeled NLS peptides, DTPA-NLS-HuM195, and DTPA-NLS-M195 (Fig. 1B).

(A) Growth inhibition of HL-60 and HL-60-MX-1 cells by 111In-HuM195, 111In-NLS-HuM195, 111In-M195, or 111In-NLS-M195 at 3 different specific activities measured using WST-1 cell viability assay. Results are expressed as mean ± SD of EC50 values (n = 3). Significant differences (P < 0.05) are indicated by asterisks. (B) Effect of 111In-NLS-mIgG, 111In-acetate, DTPA-NLS-HuM195, DTPA-NLS-M195, or NLS peptides at increasing radioactivity or mass concentrations on growth of HL-60 and HL-60-MX-1 cells. Results are expressed as mean ± SD of percentage cell growth (n = 3) relative to untreated cells.
Clonogenic Assays
A representative clonogenic survival curve for HL-60-MX-1 cells treated with 111In-NLS-M195 is shown in Figure 2, and the results of these assays for both cell lines at an SA of 3–8 MBq/μg are summarized in Table 2. No effects on the clonogenic survival of HL-60 or HL-60-MX-1 cells were observed with 111In-labeled mAbs at 1 MBq/μg, and these results are not shown. The survival of HL-60 or HL-60-MX-1 cells was reduced by more than 60% as indicated by the asymptotic values (percentage of cells surviving at the highest concentration of radioactivity added). Moreover, these asymptotic values were lower for 111In-NLS-M195 and 111In-NLS-HuM195 than those for 111In-M195 without NLS peptides and were systematically decreased with increasing SA. The D0 values represent the mean lethal concentration (mBq/cell) added to the cells. At the highest SA tested (8 MBq/μg), D0 values were 1.5- to 3-fold significantly greater for HL-60-MX-1 than for HL-60 cells. The D0 values were 2-fold significantly lower for 111In-NLS-HuM195 than for 111In-NLS-M195. D0 and asymptotic values were not calculated for 111In-NLS-mIgG, because it had no effect on the clonogenic survival of either cell line.
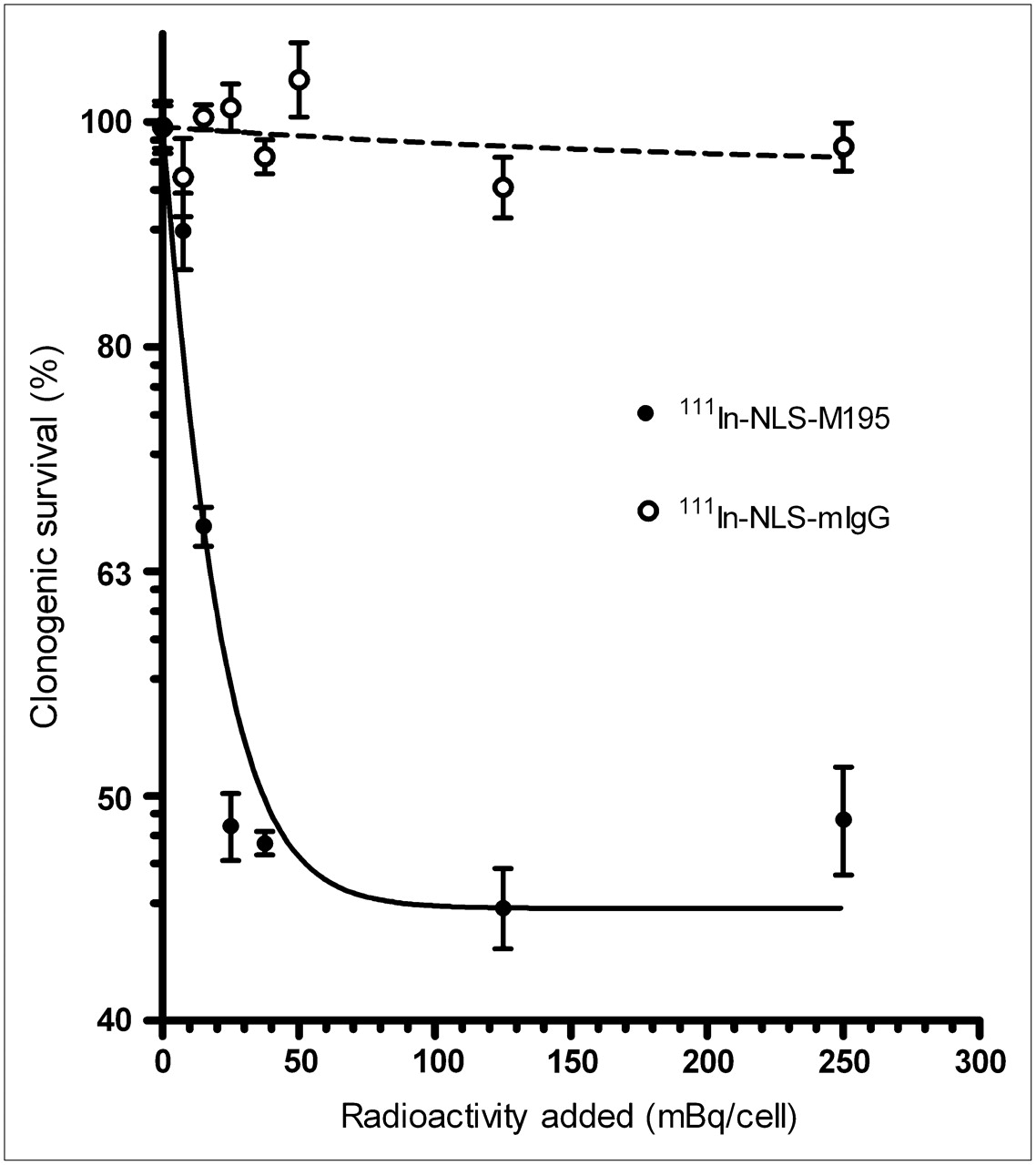
Representative clonogenic survival curve for HL-60-MX-1 cells after treatment with 111In-NLS-M195 or 111In-NLS-mIgG at SA of 8 MBq/μg. Results are expressed as mean ± SD (n = 3) of percentage cell survival relative to untreated cells.
D0 and Lower Asymptote for Clonogenic Survival of HL-60 and Mitoxantrone-Resistant HL-60-MX-1 Cells Exposed to Radioconjugates at Different Specific Activities
The results of clonogenic assays of 10 primary AML specimens exposed to 111In-NLS-M195 or 111In-M195 are shown in Table 3. 111In-NLS-M195 and 111In-M195 decreased the clonogenic survival (lower asymptote) to 23%–49% and 35%–61%, respectively. Comparisons for individual specimens revealed that no significant differences in the D0 values for 111In-NLS-M195 and 111In-M195 existed, but the asymptotic values were significantly lower in all cases for 111In-NLS-M195.
D0 and Lower Asymptote for Clonogenic Survival of Primary AML Specimens Exposed to 111In-M195 or 111In-NLS-M195 at Specific Activity of 8 MBq/μg
DISCUSSION
In this study, we showed that the Auger electron emissions from the 111In-labeled anti-CD33 mAbs HuM195 and M195 modified with NLS peptides significantly diminished the survival in vitro of mitoxantrone-resistant HL-60-MX-1 myeloid leukemia cells and primary AML specimens expressing ABC MDR transporters. This finding creates new opportunities to circumvent MDR in AML and possibly achieve remissions in patients refractory to chemotherapy, which is a major obstacle in the effective clinical management of this disease (20). Other approaches to overcoming drug resistance in AML have focused on the use of selective MDR1 inhibitors such as PSC-883 or quinine, but these have proven relatively ineffective in clinical trials (21,22). Newer inhibitors are under development that may be more effective, but they may increase toxicity from chemotherapy, because of the presence of MDR1 transporters in drug-eliminating organs (8). Auger electron therapy using 123I-labeled 4′-thio-2′-deoxyuridine, on the other hand, has proven successful in treating chemoresistant and radioresistant HL-60 cells (23). Targeted Auger electron RIT with 111In-NLS-HuM195 or 111In-NLS-M195 is not expected to be severely limited by normal-tissue toxicity, because CD33-mediated internalization and NLS-promoted nuclear importation (24) are required for manifestation of the cytotoxic effects of the Auger electrons (14,25).
Nuclear radioactivity for NLS-modified 111In-labeled M195 and HuM195 (18%–24%) was 4- to 6-fold greater than for the corresponding RICs without NLS modification. These results confirm those previously reported for 111In-NLS-HuM195 in HL-60 cells (14) but now extend these findings to drug-resistant HL-60-MX-1 cells. Nuclear localization is critical for exploiting the Auger electrons emitted by 111In for targeted radiotherapy of AML, because cellular radiation dosimetry models have revealed that the radiation-absorbed dose to the nucleus is amplified more than 34 times from 111In deposited in the nucleus, compared with on the cell surface, and is 17 times greater for nuclear than for cytoplasmic 111In (25). The efficiencies of nuclear localization of 111In-NLS-M195 or 111In-NLS-HuM195 were equivalent for HL-60 and HL-60-MX-1 cells and were not affected by expression of the drug-resistance phenotype. Assuming that a single HL-60 or HL-60-MX-1 cell was targeted to CD33 saturation (3 × 104 receptors/cell) with 111In-NLS-HuM195 or 111In-NLS-M195 at an SA of 8 MBq/μg, that 10% of the radioactivity is localized to the cell surface, that 70% is deposited into the cytoplasm, and that 20% is deposited into the nucleus with elimination only by radioactive decay, we estimate that the maximum radiation-absorbed dose to the nucleus would be about 3,067 cGy using the cellular dosimetry model of Goddu et al. (25).
Growth inhibition and decreased clonogenic survival of HL-60 and HL-60-MX-1 cells were strongly dependent on the SA of 111In-NLS-HuM195 and 111In-NLS-M195, with a minimum of 2–3 MBq/μg required for cytotoxicity (Fig. 1; Table 2). At the lowest SA (1 MBq/μg), only 1 antibody molecule in 10 is complexed to 111In, and because there are 3–4 × 104 CD33 binding sites per cell, a maximum of 3–4 × 103 111In atoms can be bound to the cell surface. Because only 20% of cell-bound and internalized 111In-NLS-HuM195 and 111In-NLS-M195 molecules will be transported into the nucleus, this further diminishes the effectiveness of these low SA RICs. However, at the higher SA examined (3, 6, or 8 MBq/μg), substantially more 111In atoms would reach the nucleus, in which they can cause lethal DNA damage. This premise is in agreement with the cytotoxic effects of HuM195 labeled with the α-emitter 213Bi reported by Nikula et al. (26), who similarly found a strong effect of SA on the clonogenic survival of HL-60 cells. In our study, at the highest SA tested (8 MBq/μg), 111In-NLS-HuM195 exhibited a 2-fold significantly lower D0 value for HL-60 cells than did 111In-NLS-M195 (5.6 ± 0.6 vs. 9.8 ± 0.8 mBq/cell, respectively; P < 0.05). This effect was not observed for lower SA RICs or for HL-60-MX-1 cells. In fact, the D0-values for HL-60-MX-1 cells were 1.5- to 3-fold significantly higher than those for HL-60 cells exposed to 111In-NLS-HuM195, 111In-NLS-M195, or 111In-M195 at an SA of 8 MBq/μg, suggesting that HL-60-MX-1 cells are modestly less sensitive to the cytotoxic effects of the Auger electrons. The reason for the apparently greater potency of high-SA 111In-NLS-HuM195 for HL-60 cells, compared with the potency of 111In-NLS-M195, is not known but may be related to its 11-fold higher binding affinity (Kd, 1.2 ± 0.4 vs. 13.9 ± 1.1 nmol/L, respectively). However, the partial resistance of HL-60-MX-1 cells, compared with that of HL-60 cells, to 111In-NLS-HuM195 cannot be explained by differences in binding affinity because the Kd value for HL-60-MX-1 cells (1.7 ± 0.6 nmol/L) was not significantly different from that for HL-60 cells. Partial resistance cannot be explained by differences in nuclear-importation efficiency either, because these were equivalent for both cell lines. It is possible both that DNA repair may account for the effects of increasing SA on the cytotoxic potency of the RICs and that variability in repair capacity or repair rates may account for the differences in responsiveness between the 2 leukemia cell lines (27).
Although the cell proliferation assays demonstrated that a high proportion (95%–98%) of the growth of HL-60 or HL-60-MX-1 cells was inhibited by exposure to high concentrations (1.7 nmol/L) and specific activities (5 MBq/μg) of 111In-NLS-HuM195 or 111In-NLS-M195 (Fig. 1A), the conditions under which the clonogenic assays were conducted revealed a lower asymptotic value of 25%–40% survival for these cells or for primary AML specimens (Tables 2 and 3). These asymptotic values were diminished for 111In-labeled RICs at increasing SAs and with NLS–peptide conjugation. A short incubation period (3 h) was used to deliver the RICs to leukemic cells in the clonogenic assays, whereas in the cell proliferation experiments, these cells were exposed continuously to the RICs for 7 d. Recycling and repopulation of CD33 due to de novo synthesis may have allowed much more 111In-NLS-HuM195 or 111In-NLS-M195 to bind, internalize, and be transported to the nucleus in these cell proliferation assays, thus enhancing the cytotoxic effect. Such receptor recycling providing high cytotoxicity from Auger electrons has been previously reported for lymphoma cells exposed to 111In-anti-CD74 mAbs (28).
A wide range of Pgp-170 and MRP1 expression in AML specimens was demonstrated, with some specimens exhibiting a high proportion of immunopositive cells (Table 1). In contrast, the prevalence of BCRP1 expression was much lower. No significant correlations between the cytotoxic response of AML specimens to 111In-NLS-M195 measured by the D0 and plateau values (Table 3) and the expression of any of these drug-resistance transporters (r2 values, −0.322–0.250; P > 0.05) were demonstrated. These results confirmed the ability of 111In-NLS-M195 to decrease the survival of mitoxantrone-resistant HL-60-MX-1 cells and further demonstrated that this effect extends to primary AML specimens with a diversity of MDR phenotypes. Taken together, these results are promising for the treatment of chemotherapy-resistant AML in patients using 111In-NLS-conjugated anti-CD33 mAbs. Humanized analogs such as 111In-NLS-HuM195 would be preferred, because of the well-recognized development of human antimouse antibodies in patients who are administered murine mAbs (29).
CONCLUSION
111In-labeled anti-CD33 mAbs HuM195 and M195 modified with NLS were routed efficiently and equivalently to the nucleus of wild-type HL-60 myeloid leukemia cells and the mitoxantrone-resistant subclone, HL-60-MX-1, after CD33-mediated internalization. Nuclear delivery enhanced the cytotoxicity of the Auger electrons emitted by 111In-NLS-HuM195 and 111In-NLS-M195. Decreased cell growth and clonogenic survival were achieved for both HL-60 and HL-60-MX-1 cells and for primary AML specimens from patients expressing MDR phenotypes. Targeted Auger electron RIT using 111In-labeled anti-CD33 mAbs modified with NLS may be able to overcome MDR and provide a means of treating chemotherapy-resistant myeloid leukemias in patients.
Acknowledgments
We gratefully acknowledge the scientific advice and technical assistance provided by Drs. Liqing Jin and John Dick (University Health Network). This research was supported by grant 166591 from the Canadian Institutes of Health Research (CIHR) to Raymond M. Reilly and by a Wyeth-CIHR fellowship (DFW-77882) to Veerle Kersemans. Parts of this study were presented at the 54th Annual Meeting of the Society of Nuclear Medicine, Washington, DC, June 2–6, 2007, and at the Annual Congress of the European Association of Nuclear Medicine, Athens, Greece, October 1–4, 2006.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication September 15, 2007.
- Accepted for publication March 19, 2008.





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- In Vitro and In Vivo Efficacy of a Novel CD33-Targeted Thorium-227 Conjugate for the Treatment of Acute Myeloid Leukemia
- Auger Electron Radioimmunotherapeutic Agent Specific for the CD123+/CD131- Phenotype of the Leukemia Stem Cell Population
- Cellular Dosimetry of 111In Using Monte Carlo N-Particle Computer Code: Comparison with Analytic Methods and Correlation with In Vitro Cytotoxicity
- Trastuzumab-Resistant Breast Cancer Cells Remain Sensitive to the Auger Electron-Emitting Radiotherapeutic Agent 111In-NLS-Trastuzumab and Are Radiosensitized by Methotrexate
- Multidrug Resistance in Acute Myeloid Leukemia: Potential New Therapeutics