


Abstract
The Auger electron–emitting radiopharmaceutical 111In-diethylenetriaminepentaacetic acid human epidermal growth factor (111In-DTPA-hEGF) binds the epidermal growth factor receptor (EGFR), is internalized, and translocates to the nucleus. The purpose of this study was to investigate the relationship between EGFR expression, DNA damage, and cytotoxicity in cells exposed to 111In-DTPA-hEGF. Methods: Breast cancer cell lines with a range of EGFR expression levels were exposed to 111In-DTPA-hEGF or γ-radiation. The cell lines (followed by number of EGFR per cell in parentheses) were MDA-MB-468 (1.3 × 106), MDA-MB-231 (1.3 × 105), and MCF-7 (1.5 × 104). The proportion of radioactivity partitioning into the nucleus was measured by cell fractionation. DNA double-strand breaks were evaluated using the γ-H2AX assay. Clonogenic survival assays were used to measure cytotoxicity. Results: All data are presented as mean ± SD. The amount of 111In-DTPA-hEGF that translocated to the nucleus (in mBq/nucleus) in MDA-MB-468, MDA-MB-231, and MCF-7 cells incubated with 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h was 131 ± 6, 8.1 ± 0.1, and 1.1 ± 0.9, respectively. The number of γ-H2AX foci per nucleus was 35 ± 15, 19 ± 10, and 1.7 ± 0.3, respectively. A reduction in the surviving fraction (SF) in MDA-MB-468 (0.013 ± 0.001) and MDA-MB-231 (0.5 ± 0.1) but not in MCF-7 cells after exposure to 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h has been demonstrated. The SF of MDA-MB-468 cells after exposure to DTPA-EGF (43 nM) and 111In-acetate (5.2 MBq/mL) for 20 h was 0.5 ± 0.1 and 0.53 ± 0.05, respectively. MDA-MB-468 was the most sensitive of the cell lines to γ-irradiation, with an SF after 2 Gy of 0.45 ± 0.04, compared with 0.7 ± 0.1 and 0.8 ± 0.1 for MCF-7 and MDA-MB-231, respectively. The number of γ-H2AX foci per nucleus in MDA-MB-468 cells correlated with the concentration, specific activity, and incubation time of 111In-DTPA-hEGF. Conclusion: DNA damage caused by 111In-DTPA-hEGF correlates with the EGFR expression level of the exposed cells and with concentration, specific activity, and incubation time of 111In-DTPA-hEGF. The γ-H2AX assay may be a useful biomarker to predict and monitor the outcome of treatment with 111In-DTPA-hEGF.
Targeted Auger electron radiation therapy is a novel approach to the treatment of disseminated cancer. The aim of the therapy is to insert lethal Auger electron–emitting radioisotopes selectively into the nuclei of cancer cells while sparing normal cells. This may be achieved by chemical conjugation of the isotopes to molecules that are able to selectively bind tumor cells, internalize, and then translocate to the nucleus. The range of most Auger electrons is comparable with the diameter of the DNA double helix (2 nm) and much less than the diameter of a cell (1). It has been shown that Auger electron–emitting isotopes such as 125I and 111In cause clustered DNA damage, including DNA double-strand breaks (DNA-dsbs) and base lesions, within 10 nucleotides from the binding site of an atom of the radionuclide (2). Given the short range of Auger electrons and the highly localized DNA damage that they cause, the success of treatment depends in part on the number of atoms of the radioisotope delivered to the nuclei of cancer cells rather than on the average dose absorbed by whole cells or organs.
Most estrogen receptor–negative, advanced breast cancers overexpress the epidermal growth factor receptor (EGFR) (3). We have shown that human epidermal growth factor (hEGF) labeled with 111In-diethylenetriaminepentaacetic acid hEGF (111In-DTPA-hEGF) selectively binds EGFR, is internalized, and translocates to the nucleus in EGFR-positive cells (4). On a molar concentration basis, 111In-DTPA-hEGF was found to be 85-, 200-, and 300-fold more effective than paclitaxel, methotrexate, and doxorubicin, respectively, at inhibiting the growth of the EGFR-overexpressing human breast cancer cell line MDA-MB-468 (5). 111In-DTPA-hEGF was also found to exert a strong antitumor effect on MDA-MB-468 xenografts in athymic mice (6). Pharmacokinetic studies of 111In-DTPA-hEGF in mice have shown that 111In-DTPA-hEGF is rapidly eliminated from the blood after intravenous administration (7). To our knowledge, no major normal tissue toxicity has been observed with doses of 111In-DTPA-hEGF as high as 44 and 85 MBq in mice and rabbits, respectively (7).
The purpose of the current study was to investigate to what extent EGFR expression level, nuclear translocation, and intrinsic cellular radiosensitivity determine the level of DNA damage and cytotoxicity caused by 111In-DTPA-hEGF. We used the γ-H2AX fluorescence assay, which is a sensitive measure of DNA-dsbs, to evaluate DNA damage (8,9). Sedelnikova et al. have reported a correlation between the number of γ-H2AX foci and survival in cells transfected with 125I-labeled triplex-forming oligonucleotides (10), suggesting that the γ-H2AX assay is useful for the evaluation of the cytotoxic effects of Auger electron–emitting DNA-targeting agents. Detailed information about the intracellular distribution and the nature and magnitude of DNA damage caused by 111In-DTPA-hEGF will be necessary for the development of a microdosimetry system for targeted Auger electron radiotherapy with this agent (11).
MATERIALS AND METHODS
Cell Culture
The human breast cancer cell lines MDA-MB-468, MDA-MB-231, and MCF-7 were purchased from the American Type Culture Collection. The cell lines were cultured at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium (DMEM) containing penicillin (100 U/mL), streptomycin (100 μg/mL), and l-glutamine (2 mmol/L) and supplemented with 10% fetal calf serum (HyClone). Subconfluent cells were harvested by trypsinization and seeded into 6-well tissue culture plates (5 × 105 cells per well). DMEM (1 mL) was added to each well, and the cells were cultured for 24 h before exposure to DTPA-hEGF, 111In-acetate, 111In-DTPA-hEGF, or γ-irradiation. A sterilized premium coverglass (22 × 22 mm) (Fisherfinest; Fisher Scientific) was placed in the wells containing cells for use in the γ-H2AX assay.
Radiopharmaceutical
111In-DTPA-hEGF (37–120 kBq/pmol) was prepared by adding 111In-chloride (MDS-Nordion, Inc.) to a kit containing DTPA-conjugated hEGF1–51 (Radiopharmacy, University Health Network) in sodium acetate buffer (1 M, pH 6) and incubating for 1–2 h at room temperature (12). The radiochemical purity of 111In-DTPA-hEGF was determined to be greater than 90% by instant thin-layer chromatography silica gel in sodium citrate buffer (100 mM, pH 5). The radioactivity of 111In-DTPA-hEGF was measured using an automatic γ-counter (Wallac 1480; Perkin Elmer). Specific radioactivity was calculated by dividing radioactivity by the mass of DTPA-hEGF used in the radiolabeling step (12).
Treatment with DTPA-hEGF, 111In-Acetate, 111In-DTPA-hEGF, and γ-Irradiation
Cells were incubated with 1 mL of DMEM with or without DTPA-hEGF (21 nM, 1 h; 43 nM, 20 h), 111In-acetate (3.2 MBq/mL, 1 h; 5.2 MBq/mL, 20 h), or 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM, 1 h; 5.2 MBq/mL, 43 nM, 20 h) before being fixed for the γ-H2AX assay or used in cell fractionation or clonogenic assays. The incubation time of 1 h was chosen to allow evaluation of the effect of initial internalization and nuclear uptake of 111In-DTPA-hEGF on DNA damage. The 20-h time point was chosen to allow evaluation of the combined effects of continuing DNA damage and DNA repair after protracted exposure to 111In-DTPA-hEGF. Cells were γ-irradiated on ice at a dose rate of 0.9 Gy/min to a total dose of 1 or 10 Gy using a 137Cs source (Gammacell 40 Exactor; MDS Nordion Inc.). After γ-irradiation, cells were cultured at 37°C for 30 min or 20 h before being fixed for the γ-H2AX assay. We used doses of 1 and 10 Gy for the 30-min and 20-h time points, respectively, as the number of γ-H2AX foci generated by these doses of γ-irradiation was comparable with that obtained after the selected exposures to 111In-DTPA-hEGF.
γ-H2AX Assay
Cells were fixed with 2% paraformaldehyde and 0.5% Triton X-100 (Sigma-Aldrich) in phosphate-buffered saline (PBS) (pH 8.2) for 15 min, permeabilized for 15 min with PBS containing 0.5% Nonidet P-40 (Sigma-Aldrich), and blocked for at least 1 h at room temperature in 2% bovine serum albumin (BSA) plus 1% donkey serum. Cells were then incubated with antiphospho-histone H2AX (Ser139, clone JBW301) mouse monoclonal IgG1 (Upstate Biotechnology) at a 1:800 dilution in 3% BSA-PBS overnight at 4°C and then with donkey antimouse IgG (H+L) (Alexa Fluor 488; Invitrogen Molecular Probes) at a 1:500 dilution for 45 min at room temperature. After each step, cells were washed with PBS for 2 × 5 min; however, after incubation with antibodies cells were washed with 0.5% BSA plus 0.175% polysorbate-20-PBS for 3 × 10 min. Coverglasses were mounted on slides (25 × 75 × 1 mm, Fisherbrand) using Vectashield mounting medium with 4′,6-diamidino-2-phenylindole (Vector Laboratories). The edges of the coverglasses were sealed with clear nail polish. From the step using the secondary antibody onward, all procedures were performed in the dark. Images were acquired with a confocal microscope (LSM 510; Zeiss) using a Plan-Apochromat 63×/1.2 water objective (Zeiss) and laser scanning set to z-stack mode with the Enterprise laser (Coherent Inc.) at 364 nm and an argon laser at 488 nm. Optical sections (1.2 μm) through the cells were imaged and combined in an extended in-depth projection. The laser intensity, signal amplification, and offset and image resolution were kept constant for all image acquisitions. The projected images were exported from an image browser (LSM; Zeiss) as tagged image file format files and processed using publicly available software (ImageJ; National Institutes of Health) with a customized macro to obtain the number of γ-H2AX foci. The parameters used in the macro, such as the threshold gray value of 20 and a minimum particle size of 8 pixels (0.1 μm), were chosen to give the best fit between automatic and manual counting. At least 30 cells were imaged per slide.
Intracellular Localization of 111In-DTPA-hEGF
The surface-bound 111In-DTPA-hEGF was obtained by applying an acid wash to cells using a solution of 200 mM of sodium acetate and 500 mM of sodium chloride (pH 2.5). Cytoplasmic and nuclear fractions were isolated using a nuclei isolation kit (Nuclei EZ Prep Nuclei Isolation Kit, NUC-101; Sigma-Aldrich). Briefly, the cells were harvested by trypsinization and the cell pellets were resuspended in 500 μL of lysis buffer on ice for 5 min. The lysed cells were centrifuged at 1,000g for 5 min to separate the nuclei (pellet) from the cytoplasm (supernatant). The pellet was resuspended in 1 mL of cold PBS. The radioactivity in the surface-bound, cytoplasmic, and nuclear fractions was measured using a Wallac 1480 γ-counter (Perkin Elmer). Nuclei were counted using a hemocytometer. The purity of the nuclear fraction was confirmed by visual microscopic inspection of the nuclei diluted in trypan blue counting solution (Sigma-Aldrich). All nuclei stained blue with a uniform circular pattern, confirming the high purity of nuclei. We have previously confirmed the high quality of this nuclear isolation procedure (13).
Clonogenic Assay
Cells were harvested by trypsinization and seeded in triplicate into 6-well plates containing DMEM. After being cultured at 37°C and 5% CO2 for 12 d, cells were stained with methylene blue (1% in a 1:1 mixture of ethanol:water). The number of colonies in each dish was counted using light microscopy. The plating efficiency was determined by dividing the number of colonies formed in the control dishes by the number of cells seeded. The surviving fraction (SF) was calculated by dividing the number of colonies by the number of cells seeded and then multiplying by the plating efficiency. The P values were determined using the 2-tailed nonparametric t test on the SFs in control and treated cultures. The data points and error bars in the figures represent the mean ± SD of 3 experiments. All the curves were generated using GraphPad Prism (version 4.03; GraphPad).
RESULTS
All data are presented as mean ± SD.
γ-H2AX Foci Induced by 111In-Acetate, DTPA-hEGF, 111In-DTPA-hEGF, and γ-Irradiation
Exposure to 111In-DTPA-hEGF resulted in a significant increase in the number of γ-H2AX foci, compared with untreated control cells, in MDA-MB-468 (10 ± 2 at 1 h; 30 ± 15 at 20 h) and MDA-MB-231 cells (7 ± 3 at 1 h; 14 ± 10 at 20 h) (Figs. 1A–1C). 111In-DTPA-hEGF did not result in a significant increase in the number of γ-H2AX foci at 1 or 20 h in MCF-7 cells (Figs. 1A–1C). DTPA-hEGF and 111In-acetate did not result in a statistically significant increase in the number of γ-H2AX foci, compared with controls, at 1 or 20 h in any of the cell lines. A marked increase above control values in the number of γ-H2AX foci per nucleus in all 3 cell lines 30 min after γ-radiation (1 Gy), with values of 16 ± 3, 10 ± 2, and 21 ± 2 for MDA-MB-468, MDA-MB-231, and MCF-7, respectively (Figs. 1A and 1C), was shown. When γ-H2AX foci were evaluated after a higher dose of γ-radiation (10 Gy) and after a longer postradiation interval (20 h), the number of foci per nucleus above control levels was 36 ± 12, 21 ± 6, and 2 ± 4 in MDA-MB-468, MDA-MB-231, and MCF-7 cells, respectively.

Induction of γ-H2AX foci by 111In-DTPA-hEGF in breast cancer cell lines. (A) Cells were incubated with DTPA-hEGF (21 nM), 111In-acetate (3.2 MBq/mL), or 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h; were exposed to γ-irradiation (1 Gy); or were untreated (control). (B) Cells were incubated with DTPA-hEGF (43 nM), 111In-acetate (5.2 MBq/mL), or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h; were exposed to γ-irradiation (10 Gy); or were untreated (controls). (C) Confocal microscopy of MCF-7, MDA-MB-231, and MDA-MB-468 cells exposed to DTPA-hEGF (21 nM), 111In-acetate (3.2 MBq/mL), or 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h; exposed to γ-irradiation (1 Gy); or untreated (control). Cells were immunostained for γ-H2AX. 4′,6-diamidino-2-phenylindole was used to visualize cell nucleus. Exposure to 111In-DTPA-hEGF caused increase in number of γ-H2AX foci in MDA-MB-468 and MDA-MB-231 cells but not in MCF-7 cells.
Subcellular Localization of 111In
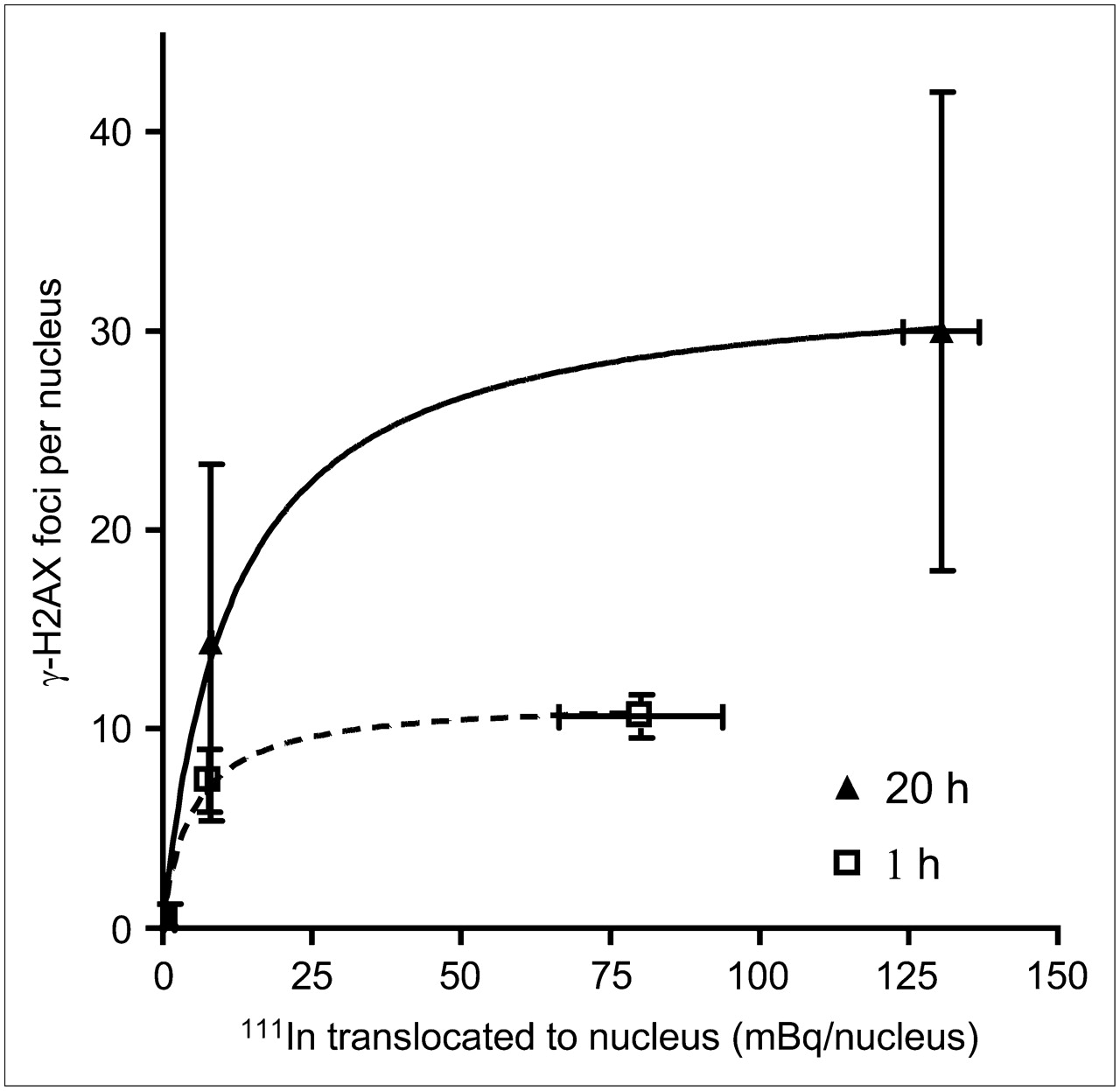
111In-DTPA-hEGF induced γ-H2AX foci in MDA-MB-468 and MDA-MB-231 cells, which have high and moderate EGFR expression levels, respectively, but did not significantly induce γ-H2AX foci in MCF-7 cells, which have low EGFR expression. This observation could be explained by greater nuclear localization of 111In-DTPA-hEGF and, therefore, a higher absorbed radiation dose in DNA due to the emission of short-range Auger electrons in cells with high EGFR expression. To confirm this, cells were exposed to 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h, and cell fractionation experiments were performed. Cells were harvested, and the amount of 111In in the cell membrane, cytoplasm, and nucleus was measured (Figs. 2A and 2B). The amount of 111In per cell decreased in the order of MDA-MB-468 > MDA-MB-231 > MCF-7. The amount of 111In-DTPA-hEGF that translocated to the nucleus (in mBq/nucleus) in MDA-MB-468, MDA-MB-231, and MCF-7 cells treated with 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h was 131 ± 6, 8.1 ± 0.1, and 1.1 ± 0.9, respectively (Fig. 2B). Perfect linear dependency for both cytoplasmic (R2 = 0.9932) and nuclear 111In (R2 = 0.9884) on surface-bound 111In (Fig. 2C) was exhibited. The dependence of γ-H2AX foci induction by 111In-DTPA-hEGF on the amount of nuclear uptake of 111In (at 1 and 20 h) is shown in Figure 3. The number of γ-H2AX foci increased as the nuclear uptake of 111In increased but leveled off at an average of 11 and 30 γ-H2AX foci per nucleus after 1- and 20-h incubations, respectively.

MDA-MB-468, MDA-MB-231, and MCF-7 cells were treated with 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h (A) or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h (B). Amount of 111In in cell membrane, cytoplasm, and nucleus was measured in cell fractionation experiments. Amount of 111In per cell decreased in order of MDA-MB-468 > MDA-MB-231 > MCF-7. (C) Amount of 111In that localizes in cytoplasm and nucleus is linearly dependent on amount of 111In that binds to cell membrane.
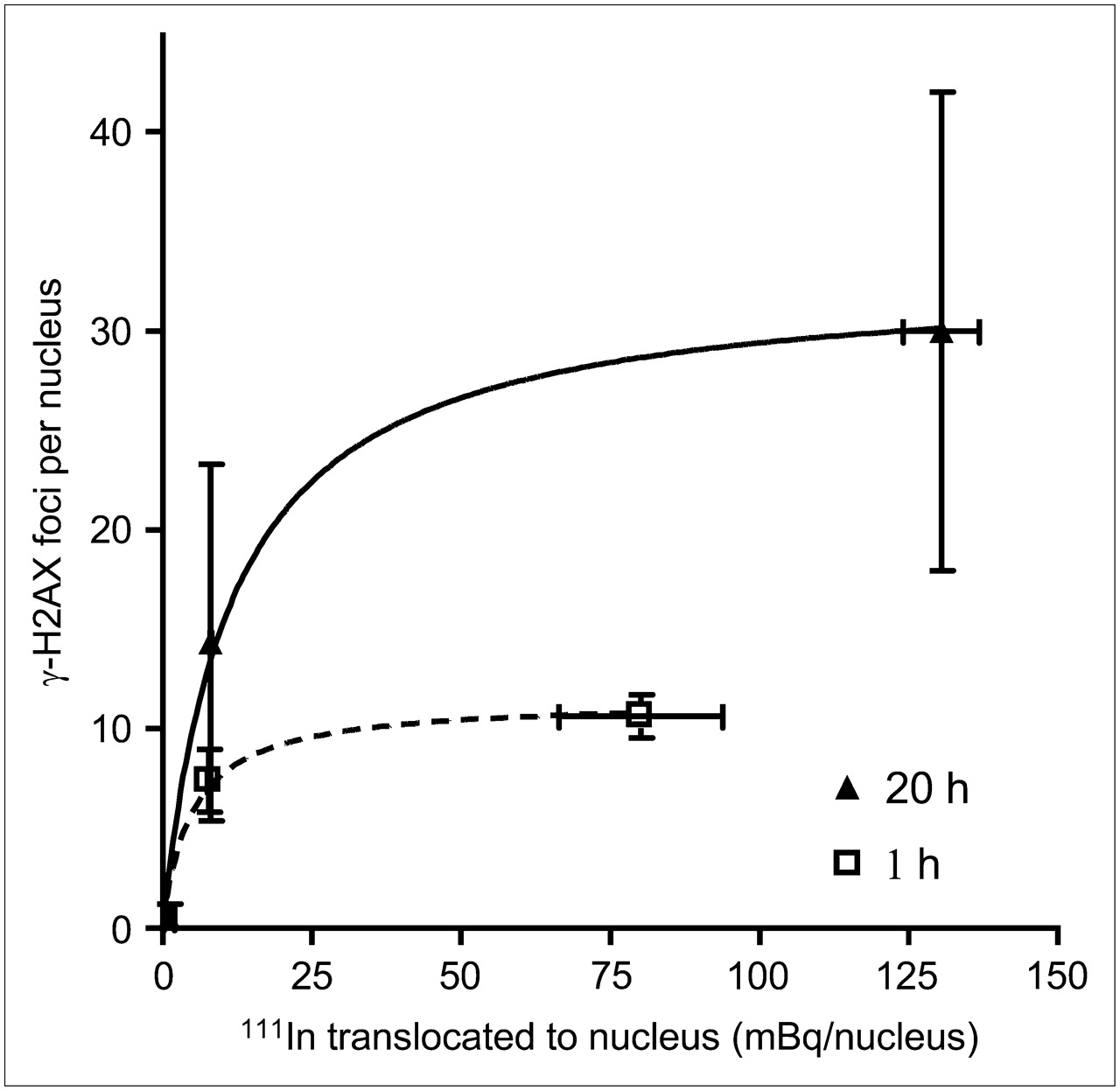
MCF-7, MDA-MB-468, and MDA-MB 231 cells were exposed to 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h. Relationship between amount of nuclear 111In and induction of γ-H2AX foci is shown. Data were fitted to 1-site binding equation.
Cytotoxicity of 111In-DTPA-hEGF and γ-Irradiation
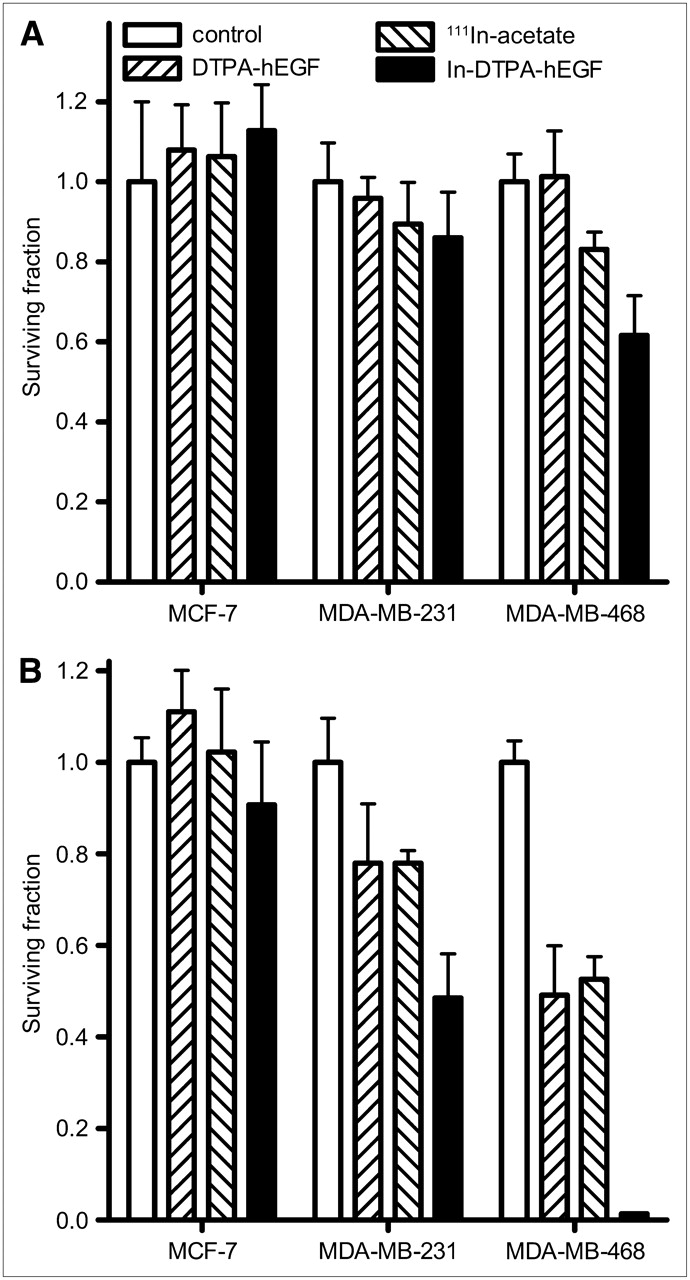
To explore the relationship between cytotoxicity of 111In-DTPA-hEGF and the level of EGFR expression, the SF of the 3 cell lines treated with 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h was measured (Figs. 4A and 4B). After exposure to 111In-DTPA-hEGF, a significant decrease in the SF, compared with untreated cells, was observed in MDA-MB-468 (0.6 ± 0.1 at 1 h and 0.013 ± 0.001 at 20 h, P < 0.0001) and MDA-MB-231 cells (0.5 ± 0.1 at 20 h, P = 0.0001) but not in MCF-7 cells. After a 20-h exposure, DTPA-hEGF (43 nM) resulted in a reduction in the SF of MDA-MB-468 (0.5 ± 0.1) and MDA-MB-231 cells (0.8 ± 0.1), consistent with the reported observation that high concentrations of EGF inhibit the growth of some EGFR-overexpressing cell lines (14,15). The same concentration of 111In-DTPA-hEGF, 43 nM, was approximately 38-fold more cytotoxic than DTPA-hEGF (SF, 0.013 ± 0.001 vs. 0.5 ± 0.1, respectively; P < 0.0001) in MDA-MB-468 cells. After a 20-h exposure, 111In-acetate caused a significant reduction in the SF of MDA-MB-468 and MDA-MB-231 cells (Fig. 4B). However, 111In-acetate, compared with 111In-DTPA-hEGF, was significantly less cytotoxic in both cell lines. To investigate whether intrinsic cellular radiosensitivity plays a role in the cytotoxicity of 111In-DTPA-hEGF, clonogenic survival curves as a function of the γ-absorbed dose (0–10 Gy) were generated for the 3 cell lines (Fig. 5). MDA-MB-468 was the most sensitive of the cell lines to γ-irradiation, with an SF after 2 Gy of 0.45 ± 0.04, compared with 0.7 ± 0.1 and 0.8 ± 0.1 for MCF-7 and MDA-MB-231, respectively.

Clonogenic assays were performed using MDA-MB-468, MDA-MB-231, and MCF-7 cells incubated with DTPA-hEGF (21 nM), 111In-acetate (3.2 MBq/mL), or 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h (A) or DTPA-hEGF (43 nM), 111In-acetate (5.2 MBq/mL), or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h (B). After exposure to 111In-DTPA-hEGF, significant decrease in SF was observed in MDA-MB-468 (0.6 ± 0.1 at 1 h and 0.013 ± 0.001 at 20 h, P < 0.0001) and MDA-MB-231 cells (0.5 ± 0.1 at 20 h, P = 0.0001) but not in MCF-7 cells, compared with untreated cells. Error bars represent SD of mean SF, calculated from 3 experiments.

Dependence of SF on γ-absorbed dose. Clonogenic assays were generated for MDA-MB-468, MDA-MB-231, and MCF-7 cells. Curves are nonlinear regression fits to experimental data, using linear quadratic function for MDA-MB-468 and first-order exponential function for other cell lines.
The Effects of 111In-DTPA-hEGF Concentration, Specific Activity, and Incubation Time on the Induction of γ-H2AX Foci in MDA-MB-468 Cells
The results of the clonogenic assays confirmed that 111In-DTPA-hEGF was most deleterious in cells that overexpress EGFR (Fig. 4). We proceeded to evaluate the effect of varying the specific activity, concentration, and incubation time of 111In-DTPA-hEGF on the induction of γ-H2AX foci in the EGFR-overexpressing cell line, MDA-MB-468. Cells were incubated with 111In-DTPA-hEGF (20 MBq/μg) over a concentration range of 0–5.2 MBq/mL for 20 h and then fixed and used for quantification of γ-H2AX foci. The number of γ-H2AX foci per nucleus increased as the concentration of 111In-DTPA-hEGF increased (Fig. 6A). MDA-MB-468 cells were incubated with 111In-DTPA-hEGF (17 nM) over a specific activity range of 0–24 MBq/μg for 20 h. The number of γ-H2AX foci per nucleus increased as the specific activity of 111In-DTPA-hEGF increased (Fig. 6B). The data were fitted to a second-order polynomial equation with R2 = 0.9438. MDA-MB-468 cells were incubated with 111In-DTPA-hEGF (1.9 MBq/mL, 11 nM) for between 0 and 20 h. The number of γ-H2AX foci per nucleus increased initially with incubation time and then reached a plateau at around 2.5 h (Fig. 6C).

Induction of γ-H2AX foci in MDA-MB-468 cells depends on concentration of 111In-DTPA-hEGF (specific activity, 20 MBq/μg) (A), specific activity of 111In-DTPA-hEGF (hEGF concentration, 17 nM) (B), and incubation time of 111In-DTPA-hEGF (1.9 MBq/mL, 11 nM) (C).
DISCUSSION
Overexpression of cell-surface receptors is a common feature of malignant cells. It is sometimes possible to exploit these receptors as molecular targets by radiolabeling peptides that bind to them. This approach has been termed peptide receptor radiation therapy (16). 111In-DTPA-hEGF is a radiolabeled peptide that binds the EGFR, is internalized, and translocates to the nucleus. 111In decays via electron capture and emits an average of 14.7 Auger electrons, 0.16 internal conversion electrons, 0.89 x-ray photons, and 1.84 γ-ray photons per decay (1). The range of γ- and x-ray photons and internal conversion electrons far exceeds the diameter of a cell, from tens of centimeters to 205 μm for the lowest energy internal conversion electron (145 keV). However, Auger electrons have much lower energy (8.5–25.5 keV) and a subcellular range of 0.25 nm to 13.6 μm (1). Because the track length of Auger electrons is so short, the amount of 111In-DTPA-hEGF that localizes in the nucleus is an important determinant of its lethality. The main purpose of the current study was to investigate the relationship between EGFR expression level and the extent of DNA damage and cytotoxicity caused by 111In-DTPA-hEGF. It will be important to understand the relationship between these parameters to assist in the selection of patients with tumors that are likely to respond to 111In-DTPA-hEGF and to provide a rational basis for calculating dose estimates.
To characterize the relationship between the extent of cell binding and cytoplasmic and nuclear uptake of 111In-DTPA-hEGF in detail, a panel of cell lines with a range of EGFR expression levels was exposed to the radiopharmaceutical at 2 different concentrations for different durations (1 and 20 h) (Fig. 2). Perfect linear dependency on surface-bound 111In for both cytoplasmic (R2 = 0.9932) and nuclear accumulation of 111In (R2 = 0.9884) (Fig. 2C) was demonstrated. The amount of cell membrane–bound 111In increased linearly as the EGFR density increased, with R2 > 0.99 after both short and long exposure times (Fig. 7A). These results indicate that the number of EGFR per cell determines how much 111In translocates to the cell nucleus after treatment with 111In-DTPA-hEGF.

(A) Linear dependence of membrane-bound 111In on number of EGFR per cell. (B) Correlation of γ-H2AX foci formation (difference between foci number in 111In-DTPA-hEGF–treated and control cells) with EGFR number per cell. Data are fitted to 1-site binding equation. (C) Correlation of γ-H2AX foci formation with SF. Cells were treated with 111In-DTPA-hEGF (3.2 MBq/mL, 21 nM) for 1 h or 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h.
The phosphorylated form of the histone H2AX, γ-H2AX, occurs at the site of DNA-dsbs. When cells are exposed to high–dose-rate γ-radiation, the number of γ-H2AX foci reaches a peak at around 30 min and then diminishes after the repair of DNA-dsbs (17,18). The peak number of γ-H2AX foci represents the maximal number of DNA-dsbs induced by radiation, whereas the residual number several hours after irradiation represents the nonrepairable DNA-dsbs and predicts subsequent cell death (17). It is likely that the kinetics of γ-H2AX foci formation after exposure to internalizing radioisotopes differ markedly from the kinetics of foci formation after high–dose-rate γ-radiation. The number of foci after prolonged exposure to an internalized radioisotope would represent a combination of residual unrepaired DNA-dsbs as well as new DNA-dsbs that are formed as a result of continuing decay of the radionuclide.
It has been shown that when MDA-MB-468 cells are exposed to 111In-DTPA-hEGF, the proportion of cell-bound radioactivity that reaches the nucleus by 1 and 24 h is approximately 3% and 10%, respectively (4). Accordingly, in this study an incubation time of 1 h was chosen to allow evaluation of the effect of initial internalization and nuclear uptake of 111In-DTPA-hEGF on DNA damage. A 20-h time point was chosen to allow evaluation of the combined effect of greater nuclear accumulation of 111In-DTPA-hEGF and DNA repair. It was found that the extent of DNA damage after both 1- and 20-h exposures to 111In-DTPA-hEGF is dependent on the number of EGFR per cell (Fig. 1). At neither time point was induction of γ-H2AX foci observed in MCF-7, the cell line with the lowest number of receptors per cell. Intermediate and high numbers of foci were observed in cell lines with moderate and high receptor number per cell, MDA-MB-231 and MDA-MB-468, respectively. The 2 control treatments used in these studies were nonradiolabeled EGF (DTPA-hEGF) and 111In-acetate. Because 111In-acetate is not specifically taken up by cells, it would not be expected to cause significant DNA damage. Neither of the control treatments caused a statistically significant induction of γ-H2AX foci, indicating that specific binding of 111In-DTPA-hEGF to EGFR is a prerequisite for the significant induction of DNA-dsbs. A 3-fold and 2-fold increase in the number of γ-H2AX foci above the background number in MDA-MB-468 and MDA-MB-231 cells, respectively, after a 20-h incubation, compared with a 1-h incubation, with the radiopharmaceutical was shown. This observation is consistent with increasing amounts of DNA damage as the radioisotope accumulates in the nucleus over time and the higher specific activity of 111In-DTPA-hEGF used for the 20- versus the 1-h exposures. It suggests that at 20 h the amount of DNA damage caused by 111In-DTPA-hEGF exceeded the capacity for DNA repair in the MDA-MB-468 and MDA-MB-231 cell lines. In contrast to the positive relationship between the extent of DNA damage caused by 111In-DTPA-hEGF and receptor number (Fig. 7B), no correlation was observed between the number of γ-H2AX foci and EGFR density at 30 min after γ-irradiation (Figs. 1A and 1C). Twenty hours after γ-radiation, a significant number of residual γ-H2AX foci in MDA-MB-468 and MDA-MB-231 cells but not in MCF-7 cells was demonstrated, suggesting that in MDA-MB-468 and MDA-MB-231 DNA repair is less efficient than in MCF-7 (Fig. 1B).
We found an increase in the number of γ-H2AX foci as the amount of 111In per cell nucleus increased to 50 mBq/nucleus, after a 20-h exposure. However, as the nuclear concentration of 111In increases, the number of γ-H2AX foci approaches a plateau at approximately 30 foci per cell (Fig. 3). A similar pattern was observed after a 1-h exposure. Others have observed, for both photon irradiation (18) and Auger electron radiation from incorporated 125I-iododeoxyuridine (19), a loss of linearity in the relationship between the number of γ-H2AX foci and absorbed radiation dose at high radiation doses. One explanation is that it is difficult, using standard image-analysis techniques, to resolve the boundaries of foci when large numbers are present. Another possible but unproven explanation for the loss of linearity of the dose–response curve shown in Figure 3 is the existence of sites within the genome that are specifically targeted by 111In-DTPA-hEGF. As such sites become fully occupied, further accumulation of isotope in the nucleus would not be expected to cause more DNA damage because of the short range of Auger electrons, which cause damage mainly in the vicinity of the decay site. It has been shown that the EGFR associates with DNA and is involved in the transactivation of specific target genes (20,21). It is therefore possible that 111In-DTPA-hEGF is directed to specific sites within the genome via its association with EGFR, although at present this is speculative.
After we demonstrated a correlation between EGFR number per cell, nuclear uptake, and the amount of DNA damage caused by 111In-DTPA-hEGF, we explored the relationship between DNA-dsb induction and cytotoxicity of the agent. Clonogenic survival assays were performed to gain insight into the response to γ-radiation of the cell lines used in this study. These assays showed that the sensitivity of the cell lines to γ-irradiation increases in the order of MDA-MB-468 > MCF-7 > MDA-MB-231 (Fig. 5). This result is consistent with published data (22,23). We found that clonogenic survival after exposure to 111In-DTPA-hEGF correlated strongly with EGFR number (Fig. 4). A significant reduction in the SF in MDA-MB-468 (0.013 ± 0.001) and MDA-MB-231 (0.5 ± 0.1) but not in MCF-7 after exposure to 111In-DTPA-hEGF (5.2 MBq/mL, 43 nM) for 20 h was shown. The high number of residual γ-H2AX foci after γ-radiation (Fig. 1B) and low SF after 2 Gy indicate that the MDA-MB-468 cell line has low DNA repair capacity and is radiosensitive. It is therefore likely that intrinsic radiosensitivity contributes to the deleterious effect of 111In-DTPA-hEGF in this cell line. However, although MDA-MB-231 cells are less sensitive to γ-irradiation than are MCF-7 cells (Fig. 5), 111In-DTPA-hEGF caused a significant reduction in SF in MDA-MB-231 but not in MCF-7 (Fig. 4B). This result indicates that EGFR expression level and not only intrinsic radiosensitivity is a determinant of the cytotoxicity of 111In-DTPA-hEGF.
It was observed that although DTPA-hEGF did not cause DNA-dsbs (Fig. 1), it did cause a reduction in SF in cell lines, with moderate and high EGFR per cell (Fig. 4). This finding is consistent with the paradoxic growth inhibitory effect of EGF reported for several cell lines, including MDA-MB-468 (15,24), and suggests that the cytotoxicity of 111In-DTPA-hEGF in MDA-MB-231 and MDA-MB-468 cells is due to a combination of the inhibitory effect of the EGF moiety of the drug plus the DNA damage caused by intranuclear radioactivity. 111In-DTPA-hEGF, then, is an Auger electron–emitting radiopharmaceutical with growth-inhibitory properties that are due to a combination of the effects of radiation and the delivery vehicle. Another example of this is 111In-trastuzumab, in which trastuzumab and the Auger electron emissions both contribute to growth inhibition (25).
We assumed that the loss of γ-H2AX foci through DNA-dsb repair during a 1-h incubation with 111In-DTPA-hEGF is negligible and estimated the yields of γ-H2AX foci per decay of 111In bound to the cell nucleus to be ≈0.2 for MDA-MB-231 and ≈0.03 for MDA-MB-468. Karamychev et al. reported that when 111In is incorporated into duplex and triplex DNA strands through carbohydrate linkers of various length, it causes breaks within 10 bases of the decay site (2). The yield of strand breaks per decay depends on the length of the linker, as this determines the exact spatial relationship between the radioactive atom and DNA. The yield was 0.05 for a triplex with a C-6 linker (1.53 nm) and 0.38 for a duplex with a C-3 linker (1.13 nm) (2). The calculated yield of γ-H2AX foci per decay of 111In in the current study is consistent with these data and suggests that after treatment with 111In-DTPA-hEGF, 111In is near DNA. However, the precise spatial relationship between 111In and DNA may vary between cell types, explaining the greater yield of DNA-dsbs in MDA-MB-231, compared with MDA-MB-468, cells. For example, differences in chromatin structure between cell lines might affect the accessibility of 111In to DNA.
Many investigators have explored the relationship between γ-H2AX foci formation and cytotoxicity for a variety of anticancer agents (17,26,27) including Auger electron–emitting pharmaceuticals (28). In the current study, it has been demonstrated that a strong correlation between the number of γ-H2AX foci and survival of breast cancer cells exposed to 111In-DTPA-hEGF (Fig. 7C, P = 0.0015, R2 = 0.9383) exists and that the number of γ-H2AX foci per nucleus in MDA-MB-468 cells increases with the concentration, specific activity, and incubation time of 111In-DTPA-hEGF (Fig. 6). The γ-H2AX assay has been applied successfully to clinical samples to evaluate intrinsic DNA repair capacity (29) and radiation exposure (30), individual radiosensitivity of DNA (31), and as a biomarker for early cancer (32). These successful trials suggest that the γ-H2AX assay could be applied to tumor samples after a test dose of 111In-DTPA-hEGF and used as a biomarker to predict response to this treatment.
CONCLUSION
The induction of phosphorylated H2AX correlates with the survival of breast cancer cells after exposure to 111In-DTPA-hEGF. The number of γ-H2AX foci also correlates with the EGFR expression level of the exposed cells and with the concentration, specific activity, and incubation time of 111In-DTPA-hEGF. The γ-H2AX assay may be a useful biomarker to predict and monitor the outcome of treatment with 111In-DTPA-hEGF.
Acknowledgments
We thank Shichong Liu for her assistance with image analysis. This research was supported through grants from the Canadian Breast Cancer Foundation (Ontario Chapter) and Cancer Research-U.K. Zhongli Cai and Kristy Bailey received support from the Canadian Institutes for Health Research (through the Excellence in Radiation Research for the 21st Century Training Program).
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication September 5, 2007.
- Accepted for publication May 7, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Molecular Radiotherapy Using Cleavable Radioimmunoconjugates That Target EGFR and {gamma}H2AX
- Inactivation of HNSCC Cells by 90Y-Labeled Cetuximab Strictly Depends on the Number of Induced DNA Double-Strand Breaks
- Auger Electron Radioimmunotherapeutic Agent Specific for the CD123+/CD131- Phenotype of the Leukemia Stem Cell Population
- Imaging DNA Damage In Vivo Using {gamma}H2AX-Targeted Immunoconjugates
- ErbB-2 Blockade and Prenyltransferase Inhibition Alter Epidermal Growth Factor and Epidermal Growth Factor Receptor Trafficking and Enhance 111In-DTPA-hEGF Auger Electron Radiation Therapy
- Cellular Dosimetry of 111In Using Monte Carlo N-Particle Computer Code: Comparison with Analytic Methods and Correlation with In Vitro Cytotoxicity
- Monitoring Drug-Induced {gamma}H2AX as a Pharmacodynamic Biomarker in Individual Circulating Tumor Cells