


Abstract
18F-FDG, a marker of the enhanced metabolism characteristic of activated inflammatory cells, and 99mTc-annexin A5, a marker of apoptosis, are both widely believed to be useful for the imaging of unstable atheroma (rupture-prone vulnerable plaques [VP]). Serum cholesterol functions as a proinflammatory factor, driving the formation of VP, and affects the immune responses of aortic tissues systemically. It is therefore reasonable to postulate that prolonged cholesterol loading may alter the aortic uptake of these tracers. Here, we evaluated the aortic uptake of 18F-FDG and 99mTc-annexin A5 in apolipoprotein E-deficient (apoE−/−) and wild-type mice placed on high-fat diets. Methods: Male apoE−/− and wild-type (C57BL/6J) mice were maintained on high-fat diets after the age of 5 wk. Wild-type mice fed regular chow were used as controls. At the ages of 10, 18, and 25 wk (5–15 mice per group at each time point), mice were injected with 18F-FDG or 99mTc-annexin A5 after 12 h of fasting. At 1 h after 18F-FDG injection (or 2 h after 99mTc-annexin A5 injection), mice were sacrificed, and the aortas were removed for well-type scintillation counting of radioactivity. The results were expressed as percentage injected dose per gram of tissue and normalized by animal body weight [(ID%/g) × kg]. En face staining was then performed to assess the location and size (surface area) of the lipid pool within each aortic specimen. Concurrent blood samples were obtained to determine the plasma lipid profile of each group. Results: No atherosclerotic lesions were found in wild-type mice regardless of the diet, whereas the lesion area progressively increased with age in apoE−/− mice. Mean plasma cholesterol levels remained stable with the regular diet in wild-type mice (73–78 mg/dL) but increased with cholesterol feeding in wild-type mice (143–179 mg/dL) and in apoE−/− mice (>1,300 mg/dL). Aortic tracer uptake [(ID%/g) × kg] remained stable with the regular diet in wild-type mice (0.054–0.053 and 0.021–0.023 for 99mTc-annexin A5) but increased with cholesterol feeding in wild-type mice (0.164 for 18F-FDG and 0.036 for 99mTc-annexin A5 at 25 wk) and in apoE−/− mice (0.249 for 18F-FDG and 0.047 for 99mTc-annexin A5 at 25 wk). Conclusion: The accumulation of 18F-FDG and 99mTc-annexin A5 in aortic tissues is influenced not only by the progression of atherosclerotic disease but also by cholesterol loading over time.
The spontaneous rupture of vulnerable plaques (VP) is a common cause of both myocardial and cerebral infarctions (1,2). The detection of VP is therefore clinically important for patient risk stratification and the delivery of early dietary interventions and lipid-lowering treatments. Nuclear imaging can noninvasively quantify 2 of the most important determinants of plaque vulnerability, namely, active inflammation (with 18F-FDG PET) and apoptosis (with 99mTc-annexin A5 SPECT) (3–7).
Serum cholesterol, particularly oxidized low-density lipoprotein (LDL) cholesterol, acts as a proinflammatory factor (8) that increases the systemic levels of many immune system markers (9,10). For example, modified LDL stimulates endothelial cells to express monocyte chemotactic protein 1, which in turn recruits monocytes into the arterial wall (11,12). Monocyte chemotactic protein 1 also promotes the differentiation of infiltrating monocytes into macrophages and foam cells (13). Apoptosis of inflammatory and vascular smooth muscle cells of the arterial wall is an integral feature of the pathogenesis of VP (14). An increased rate of apoptosis in the diseased arterial wall is associated with high levels of circulating oxidized LDL (15–17). Elevated levels of serum cholesterol may therefore influence the extent of aortic plaque inflammation and apoptosis, leading to the increased uptake of 18F-FDG and 99mTc-annexin A5, respectively. In this investigation, we attempted to determine the effects of prolonged cholesterol loading on the aortic uptake of 18F-FDG and 99mTc-annexin A5.
We used the apolipoprotein E-deficient (apoE−/−) murine model of atherosclerosis (18). When fed a high-fat diet, apoE−/− mice develop severe hypercholesterolemia and atherosclerotic disease similar to human disease. In contrast, wild-type mice show only moderate hypercholesterolemia while on a high-fat diet. We evaluated the effects of serum cholesterol levels on the aortic uptake of 18F-FDG and 99mTc-annexin A5 in both wild-type and apoE−/− mice placed on high-fat diets over time.
MATERIALS AND METHODS
Preparations of 18F-FDG and 99mTc-Annexin A5
18F-FDG was obtained from Hokkaido University Hospital, which produces the tracer for clinical use. Recombinant human annexin A5 derivatized with hydrazinonicotinamide was kindly donated by Theseus Imaging Corp. Hydrazinonicotinamide-annexin A5 was labeled with 99mTc by use of tricine as the coligand as described previously (specific activity: 4.8 MBq/μg of protein) (19).
Animal Studies
Animal care and all experimental procedures were performed with the approval of the Animal Care Committee at Hokkaido University. Studies were performed with male apoE−/− mice (5–14 per group) and male wild-type mice (C57BL/6J; 5–15 per group) obtained from Taconic Gnotobiotic Center and Clea Japan Inc., respectively. After the age of 5 wk, apoE−/− mice and wild-type mice were maintained on a high-fat, high-cholesterol diet (21% fat, 0.15% cholesterol, no cholate; diet TD88137 [Harland Teklad]) (ChD–apoE−/− mice and ChD–wild-type mice, respectively). Additional age-matched male wild-type mice were maintained on a laboratory chow basal diet (Oriental MF [Oriental Yeast Co. Ltd.]) for use as controls (normal diet–wild-type mice). All animals were kept on a 12-h light cycle with free access to food and water in a temperature-controlled facility at the Laboratory of Animal Experiments at Hokkaido University.
At the ages of 10, 18, and 25 wk, the animals were fasted, anesthetized with pentobarbital (0.025 mg/kg of body weight, intraperitoneally), and injected with either 18F-FDG (18.5 MBq/mouse) or 99mTc-annexin A5 (18.5 MBq/mouse). The blood glucose levels of all animals were measured immediately before the injection. At 1 h after the 18F-FDG injection or 2 h after the 99mTc-annexin A5 injection, blood samples (0.9–1.5 mL/mouse) were collected under deep pentobarbital anesthesia for subsequent analyses. Aortas were fixed by cardiac perfusion with a cold 0.1 M phosphate buffer solution (pH 7.4) and then a cold fixative (4% paraformaldehyde and 0.1 M phosphate buffer solution [pH 7.4]), removed, and weighed. Radioactivity in each aorta was measured in a well-type scintillation counter. The results were expressed as percentage injected dose per gram of tissue and normalized by animal body weight [(ID%/g) × kg].
Evaluation of Atherosclerotic Lesions
After radioactivity counting was complete, aortic specimens were longitudinally incised, mounted on clean glass slides, and stained with Oil Red O (Sigma) by use of the following protocol (20). In brief, each specimen was rinsed in 60% 2-propanol for 3 min, incubated in Oil Red O solution at 37°C for 40 min, destained in 60% 2-propanol for 6 min, and then mounted on a glass slide with aqueous mounting medium (Biomeda Corp.). Stained specimens were examined microscopically (BX50; Olympus), and the percentage of positively stained surface area for each aortic specimen was calculated.
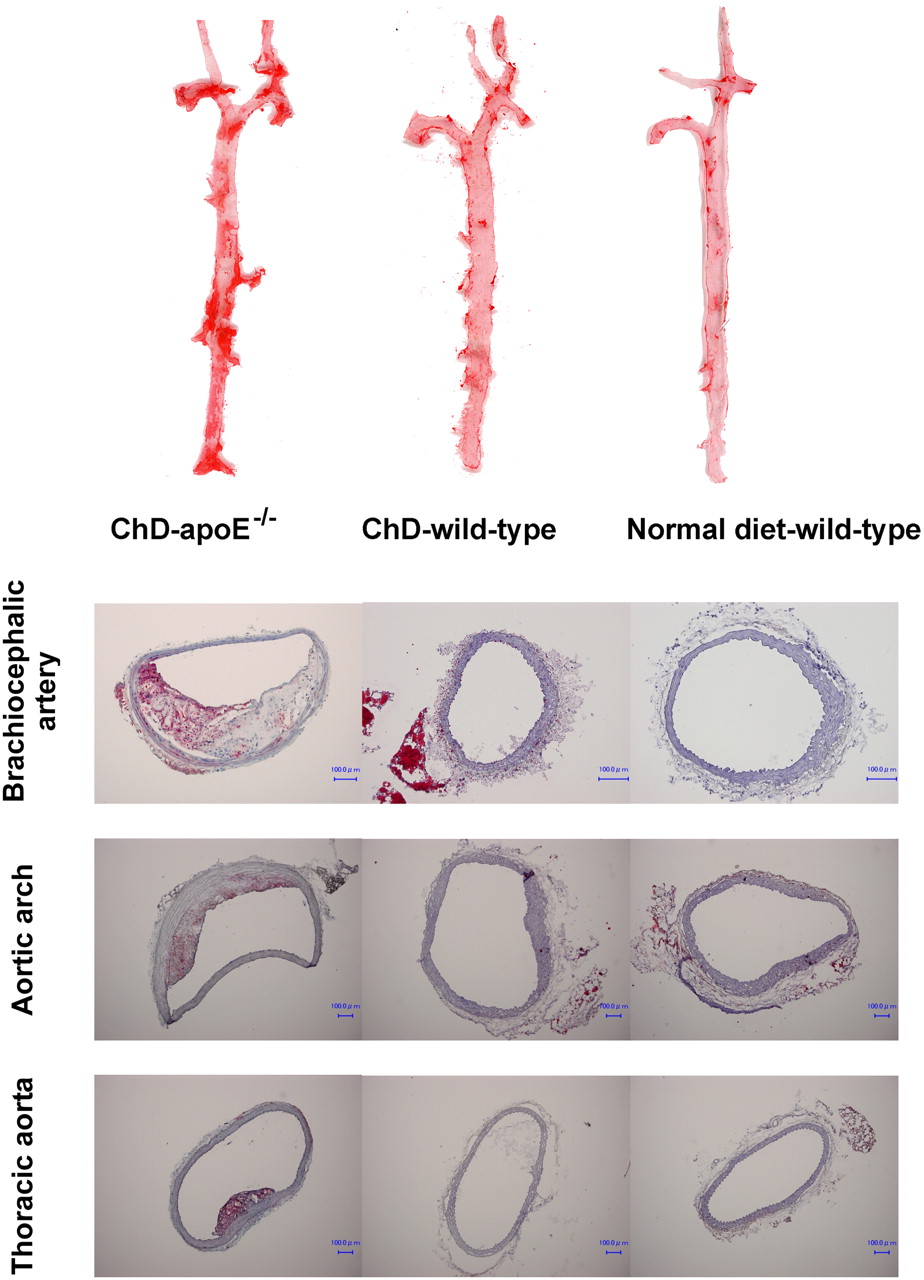
Oil Red O staining and immunohistochemical staining with a mouse macrophage–specific antibody (Mac-2, clone m3/38; Cedarlane) were performed by use of standard procedures (20) with 5-μm-thick cryostat cross sections of aortic tissues from 4 additional mice at 25 wk of age in each group (ChD–apoE−/−, ChD–wild-type, and normal diet–wild-type mice) to confirm the presence of atherosclerotic disease (Fig. 1) and macrophage infiltration (Supplemental Fig. 1) (supplemental materials are available online only at http://jnm.snmjournals.org).
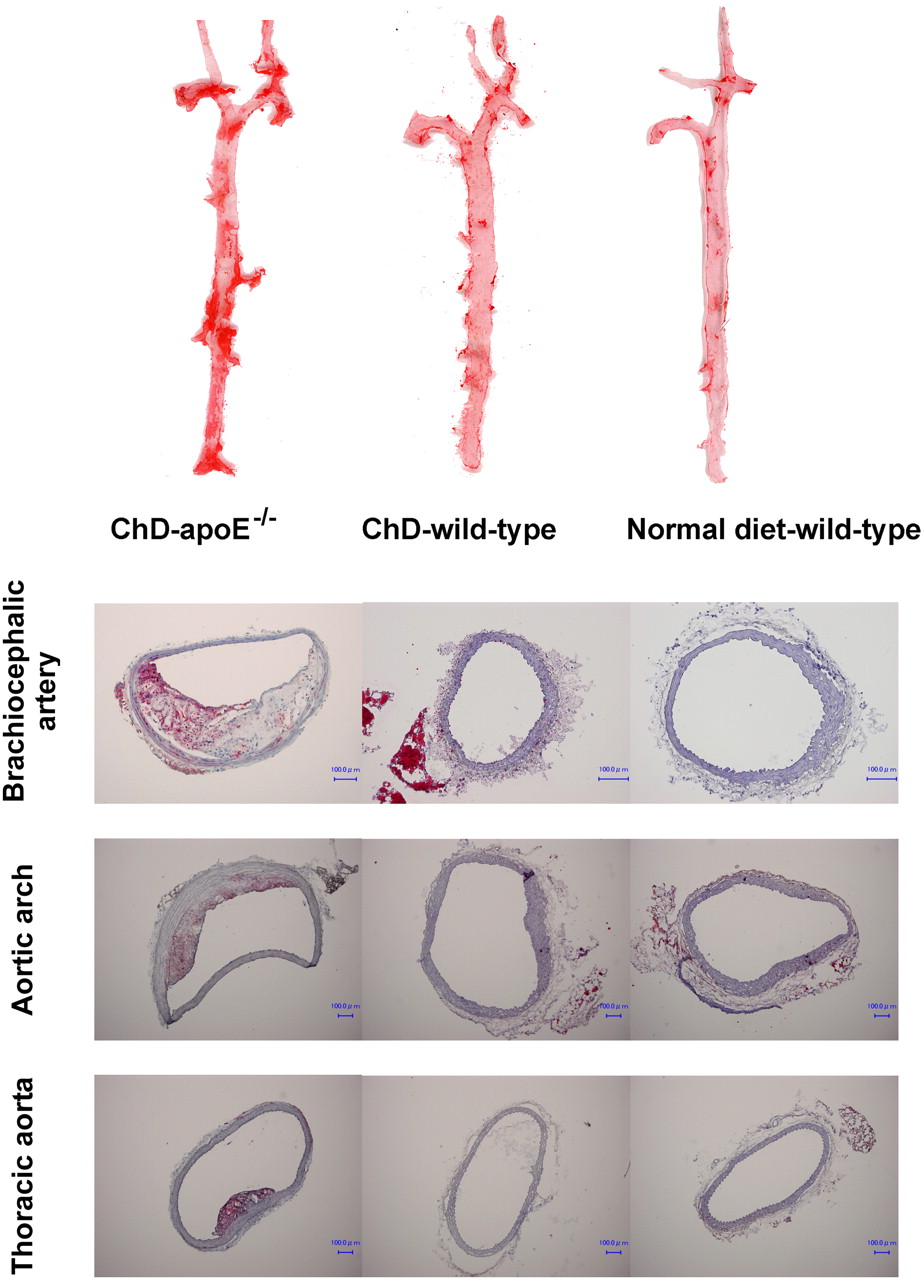
(Top) Examples of longitudinally incised aortas obtained from ChD–apoE−/−, ChD–wild-type, and normal diet–wild-type mice and stained with Oil Red O (at age of 25 wk). Regions stained red with Oil Red O represent atherosclerotic lesions. (Bottom) Aortic tissues were also examined by cryostat sectioning at same age, and atherosclerotic plaques were confirmed histologically in ChD–apoE−/− mice but not in ChD–wild-type or normal diet–wild-type mice. Bar = 100 μm.
Statistical Analysis
Numeric parameters were expressed as mean ± SD. To evaluate the effects of animal treatments (regular diet feeding, cholesterol feeding, and cholesterol feeding plus apolipoprotein E deficiency) or age (duration of treatment) on body weight, serum cholesterol levels, blood glucose levels, and aortic tracer uptake, the data were first tested with Bartlett's test for homogeneity of variance. If Bartlett's test confirmed homogeneity of variance, that is, a Bartlett's test value of greater than 0.05, then a 1-way ANOVA followed by a Bonferroni post hoc test was performed; if Bartlett's test indicated heterogeneity of variance, that is, a Bartlett's test value of less than 0.05 (cross-sectional comparisons for parameters; longitudinal comparisons for body weight and aortic 18F-FDG uptake), then a nonparametric Kruskal–Wallis test followed by a Games–Howell multiple comparisons test was performed (Tables 1–3⇓⇓). A 2-way factorial ANOVA was performed to evaluate the degree of atherosclerotic lesion development with age in ChD–apoE−/− mice given 18F-FDG and 99mTc-annexin A5 (Fig. 2).
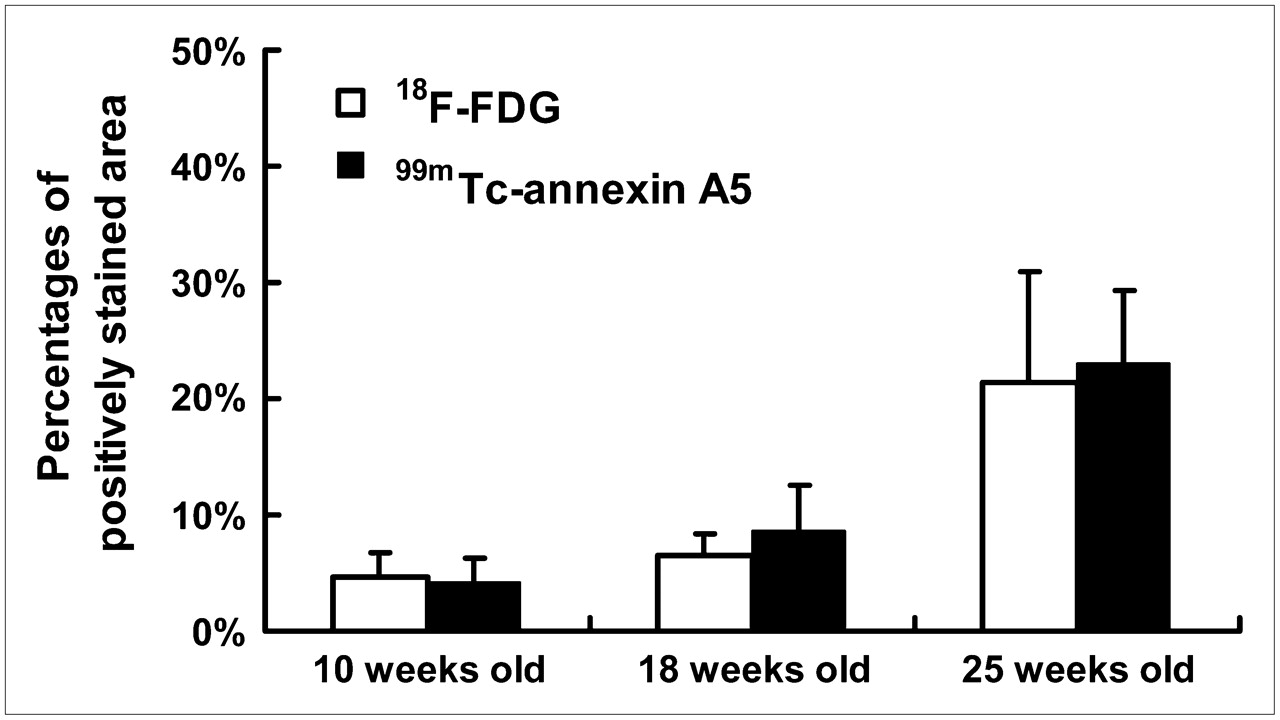
Percentages of surface area positively stained with Oil Red O in descending aorta of apoE−/− mice after different periods of high-fat feeding (ChD–apoE−/− mice). Percentages of positively stained surface area significantly increased with age (F = 53.38; P < 0.001). No significant differences were observed in percentages of positively stained area between groups given 18F-FDG and 99mTc-annexin A5 (F = 0.51; P = 0.48).
Body Weight and Serum Lipid Levels
Blood Glucose Levels in Group Given 18F-FDG
Aortic Tracer Uptake Levels
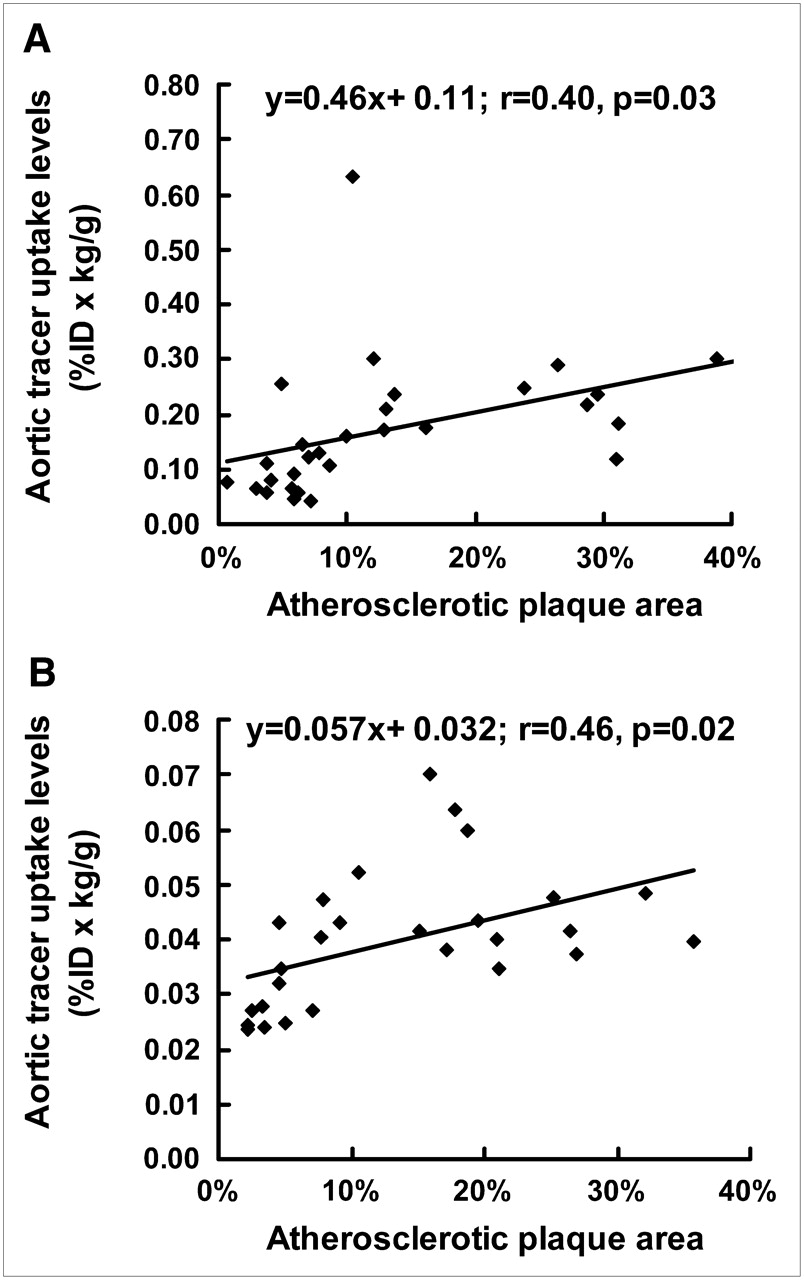
A linear regression analysis correlating the percentages of aortic surface area occupied by atherosclerotic plaques with aortic tracer uptake in ChD–apoE−/− mice given 18F-FDG and 99mTc-annexin A5 was performed (Fig. 3). The correlation between LDL cholesterol levels and aortic tracer uptake was assessed with the Spearman correlation coefficient by rank analysis in mice given 18F-FDG and 99mTc-annexin A5 at each age (Supplemental Fig. 2).

Correlations between percentages of aortic surface area occupied by atherosclerotic plaques and aortic tracer uptake in ChD–apoE−/− mice. Correlations were calculated with linear regression analysis for groups given 18F-FDG (A) and 99mTc-annexin A5 (B).
A 2-tailed P value of less than 0.05 was considered statistically significant.
RESULTS
Body Weight, Serum Cholesterol Levels, and Blood Glucose Levels
Table 1 shows the age, body weight, and serum cholesterol levels for each group of treated mice. Body weight increased substantially with age in all treatment groups. ChD–apoE−/− mice showed the greatest gain in body weight, followed by ChD–wild-type and normal diet–wild-type mice. Severe hypercholesterolemia and moderate hypercholesterolemia were observed in ChD–apoE−/− and ChD–wild-type mice, respectively, throughout the experimental period. Total cholesterol and LDL cholesterol levels were significantly higher in ChD–apoE−/− mice than in ChD–wild-type and normal diet–wild-type mice. Total cholesterol and LDL cholesterol levels in ChD–wild-type mice were also significantly higher than those in normal diet–wild-type mice. High-density lipoprotein (HDL) cholesterol levels in ChD–apoE−/− mice were significantly lower than those in ChD–wild-type and normal diet–wild-type mice for each age group. ChD–wild-type mice showed significantly higher HDL cholesterol levels than normal diet–wild-type mice. Blood glucose levels were stable in each treatment group of mice at different ages and were within normal ranges (Table 2). No significant differences were observed among treatment groups at each age, except for a significant but slight difference between ChD–apoE−/− mice and ChD–wild-type mice at 25 wk old.
Atherosclerotic Lesion Development
Early to histologically advanced atheromatous lesions were observed throughout the aortas of older ChD–apoE−/− mice. Advanced lesions (VP) were predominantly located in the brachiocephalic arteries and lesser curvature of the aortic arch. No atherosclerotic lesions were found in wild-type mice regardless of diet (ChD–wild-type mice and normal diet–wild-type mice) (Fig. 1). Immunohistochemical staining with Mac-2 showed infiltration of macrophages in atherosclerotic plaques of ChD–apoE−/− mice but not ChD–wild-type or normal diet–wild-type mice (Supplemental Fig. 1).
Figure 2 shows a summary of atherosclerotic lesion development in ChD–apoE−/− mice as a percentage of total surface area occupied by regions positively stained with Oil Red O. In these mice, the total atherosclerotic lesion area increased significantly with age. No significant differences were noted in atherosclerotic lesion development between mice given 18F-FDG and those given 99mTc-annexin A5.
Aortic Tracer Distribution
The aortic uptake of 18F-FDG and 99mTc-annexin A5 is shown in Table 3. In normal diet–wild-type mice, the uptake of 18F-FDG and 99mTc-annexin A5 remained unchanged throughout the experimental period. The level of aortic uptake of 18F-FDG in ChD–wild-type mice increased progressively with age (P < 0.0167 for 10- vs. 18-wk-old mice and 18- vs. 25-wk-old mice), whereas that of 99mTc-annexin A5 reached a plateau at 18 wk and remained unchanged at 25 wk (P < 0.0167 for 10- vs. 18-wk-old mice). ChD–apoE−/− mice showed significantly higher levels of aortic uptake of 18F-FDG and 99mTc-annexin A5 with age (P < 0.0167 for 10- vs. 18-wk-old mice and 18- vs. 25-wk-old mice for 18F-FDG and 10- vs. 18-wk-old mice for 99mTc-annexin A5).
ChD–apoE−/− and ChD–wild-type mice showed significantly higher levels of aortic uptake of 18F-FDG and 99mTc-annexin A5 than normal diet–wild-type mice at 18 and 25 wk. ChD–apoE−/− mice also tended to have higher levels of uptake of 18F-FDG and 99mTc-annexin A5 than ChD–wild-type mice of the same age.
Correlation Between Atherosclerosis Progression and Aortic Tracer Accumulation
The surface area of the aorta that stained positively with Oil Red O was determined for ChD–apoE−/− mice after different periods of high-fat feeding. For both tracers, the aortic tracer uptake levels correlated directly with the progression of atherosclerosis (r = 0.40, P = 0.03, for mice given 18F-FDG; r = 0.46, P = 0.02, for mice given 99mTc-annexin A5) (Fig. 3).
Correlation Between LDL Cholesterol Levels and Aortic Tracer Accumulation
Positive correlations were observed between LDL cholesterol levels and aortic tracer uptake levels in both mice given 18F-FDG and mice given 99mTc-annexin A5 for each age group, except for 10-wk-old mice given 18F-FDG (Supplemental Fig. 2).
DISCUSSION
To determine the effects of prolonged cholesterol loading on the aortic uptake of 18F-FDG and 99mTc-annexin A5, we used the following models: apoE−/− mice fed a high-fat diet (ChD–apoE−/− mice), wild-type mice fed a high-fat diet (ChD–wild-type mice), and wild-type mice fed a regular chow diet (normal diet–wild-type mice).
Using these diets and different genetic backgrounds, we were able to separate mice into 3 distinct categories based on serum cholesterol levels or atherosclerotic status (Table 1; Figs. 1 and 2). ChD–apoE−/− mice showed severe hypercholesterolemia, with extremely high serum LDL cholesterol levels, and all histologic grades of atherosclerotic disease. ChD–wild-type mice showed elevated serum LDL cholesterol levels but no histologic evidence of atherosclerotic disease. Normal diet–wild-type mice showed low serum cholesterol levels with increasing age and no evidence of atherosclerotic disease. These murine models also demonstrated different levels of aortic uptake of 18F-FDG and 99mTc-annexin A5 (Table 3). For ChD–apoE−/− and ChD–wild-type mice, the levels of aortic accumulation of 18F-FDG and 99mTc-annexin A5 tended to increase with age. A higher level of aortic uptake of each tracer was noted for ChD–apoE−/− mice than for age-matched ChD–wild-type mice. In the last model (normal diet–wild-type mice), the levels of aortic uptake of both 18F-FDG and 99mTc-annexin A5 showed no significant change at any age. In addition, these values were significantly lower than the values obtained in ChD–wild-type and ChD–apoE−/− mice. Our results indicated that the aortic uptake of 18F-FDG and 99mTc-annexin A5 increased with the progression of atherosclerotic disease, underscoring the value of both tracers in the differential diagnosis of atherosclerosis. A caveat for the use of 18F-FDG and 99mTc-annexin A5 in the diagnosis of atherosclerotic disease, however, is our observation that cholesterol feeding can actually increase the accumulation of these tracers in the aorta without any visible atheroma.
On the other hand, the uptake of 18F-FDG and 99mTc-annexin A5 correlated directly with the extent of atherosclerotic lesions and the development of lesion severity in ChD–apoE−/− mice (Fig. 3), with uptake values absolutely higher than that in aging wild-type mice on either diet (Table 3). The uptake of 18F-FDG and 99mTc-annexin A5 into atherosclerotic lesions and the correlation between tracer uptake levels and lesion vulnerability have also been confirmed by other experimental and clinical studies (20–25); our results are in agreement with those of previous investigations.
In addition to accelerating atherosclerosis in apoE−/− mice, high-fat feeding significantly increased plasma cholesterol levels and body weight in both apoE−/− and wild-type mice. These factors may also play important roles in the alteration of aortic tracer uptake. LDL cholesterol acts as a “bad cholesterol”; in particular, oxidized LDL cholesterol acts as a proinflammatory factor (8) that increases the levels of inflammatory mediators (9,10), decreases nitric oxide levels, and stimulates atherogenesis (26). Therefore, ChD–wild-type mice with elevated LDL cholesterol levels may in fact have chronic low-grade vascular inflammation. Older ChD–wild-type mice in our study were obese and had moderate hyperlipidemia. In previous investigations, C57BL/6J mice were used for the study of diet-induced obesity and diabetes (27–29). Furthermore, clinical investigations have shown that the plasma concentrations of tumor necrosis factor α, interleukin 6, C-reactive protein, and other inflammatory mediators are significantly elevated in obese individuals (30–32). Both lines of evidence suggest that there were elevated levels of inflammatory mediators in the ChD–wild-type mice used in the present study. Another line of evidence supporting our hypothesis was that described by Molnar et al. (33), who found that although high-fat feeding induced endothelial cell dysfunction, it did not enhance neointimal formation in C57BL/6J mice. In a recent clinical study, Yun et al. demonstrated that among several atherosclerosis risk factors (such as hypertension and diabetes), only hypercholesterolemia and age were consistently correlated with 18F-FDG uptake levels in large arteries (34). Hypercholesterolemia expressed as high LDL levels may relate to the grade of vascular inflammation, and age representing the duration of cholesterol loading may also be required for delayed functional changes. In agreement with the results of Yun et al. (34), both total cholesterol and LDL levels and the duration of cholesterol loading played integral roles in driving aortic tracer uptake in the present study. When subjects underwent sufficiently prolonged cholesterol loading, the enhancement of aortic tracer uptake appeared to be associated with the severity of hypercholesterolemia. Assessment with the Spearman correlation coefficient by rank analysis indicated positive correlations between aortic tracer uptake and total cholesterol and LDL levels in both mice given 18F-FDG and mice given 99mTc-annexin A5 for each age group, except for 10-wk-old mice given 18F-FDG (Supplemental Fig. 2); however, a detailed correlation analysis could not be performed because of the large differences in LDL cholesterol levels between ChD–apoE−/− mice (>1,300 mg/dL) and wild-type mice (15–19 mg/dL for normal diet and 44–74 mg/dL for high-fat diet).
The duration of hypercholesterolemia is also an important factor. In the present study, prolonged high-fat feeding appeared to be necessary to change tracer uptake in the aorta. For example, although LDL cholesterol levels were 30-fold higher in ChD–apoE−/− mice than in ChD–wild-type mice at 10 wk (after 5 wk of cholesterol feeding), there were no significant differences in aortic tracer uptake. However, ChD–wild-type mice at 25 wk showed far lower LDL cholesterol levels but significantly higher levels of aortic tracer uptake than ChD–apoE−/− mice at 10 wk. Therefore, it is possible that the duration of cholesterol loading as opposed to the severity of hypercholesterolemia was the predominant factor driving tracer uptake.
With regard to HDL levels, Tahara et al. (35) demonstrated that the reduction in 18F-FDG uptake was related to the enhancement of HDL levels with statin therapy for 3 mo. In contrast, the high-fat diet used in the present study increased not only LDL but also HDL levels in C57BL/6J mice, a result consistent with those of a previous report (36). The enhanced HDL levels did not prevent the increases in aortic tracer uptake in ChD–wild-type mice; therefore, no relationship was observed between aortic tracer uptake and HDL cholesterol levels in the present study. It has been reported that dietary effects on HDL levels are different in humans and mice. In humans, most cholesterol is carried in LDL, whereas mice carry most of their plasma cholesterol in HDL (37). Therefore, the relationship between tracer uptake and HDL levels and the species differences with regard to this point remain to be clarified.
It is known that 18F-FDG is transported into aortic tissues through 2 types of glucose-transporting protein: Glut-1 and Glut-4 (38–40). Glut-1 is primarily responsible for the basal (i.e., insulin-independent) transport of glucose. Glut-4 is an insulin-sensitive glucose transporter that is greatly affected by blood insulin levels. Elevated blood glucose levels commonly enhance insulin levels and consequently increase 18F-FDG as well as glucose accumulation in aortic tissues through Glut-4. In the present study, despite the higher blood glucose levels in ChD–wild-type mice than in ChD–apoE−/− mice at 25 wk (Table 2), ChD–apoE−/− mice still showed significantly higher levels of 18F-FDG uptake in aortic tissues (Table 3). Accordingly, the slight difference in blood glucose levels observed in the present study appeared not to significantly affect aortic 18F-FDG uptake.
In the present study, we could not apply dual-tracer techniques to measure aortic tracer uptake. Consequently, the aortic uptake of 18F-FDG was determined separately from that of 99mTc-annexin A5 in different groups of mice; this study design prevented us from directly evaluating the correlation of the aortic uptake of the 2 tracers in the same animals. It is of great interest to determine the correlation between18F-FDG uptake and 99mTc-annexin A5 uptake, because both 18F-FDG and 99mTc-annexin A5 are widely believed to be useful for the detection of unstable atheroma. Further studies with dual-tracer techniques are required for comparisons of 18F-FDG uptake and 99mTc-annexin A5 uptake.
CONCLUSION
In the present study, we reported time-dependent changes in the aortic uptake of 18F-FDG and 99mTc-annexin A5 in 3 murine models. After a prolonged high-fat diet, apoE−/− mice showed the highest levels of aortic uptake. Wild-type mice fed a high-fat diet showed higher aortic uptake levels than those fed a regular diet—but still lower than those of apoE−/− mice fed a high-fat diet. These findings indicate that not only the progression of atherosclerosis but also other proinflammatory factors, including prolonged cholesterol loading, alter the aortic uptake of 18F-FDG and 99mTc-annexin A5. Tracer uptake should be assessed with caution in patients with proinflammatory risk factors, because the specificity for atherosclerotic disease may be adversely affected.
Acknowledgments
This study was performed through Special Coordination Funds for Promoting Science and Technology of the Ministry of Education, Culture, Sports, Science and Technology of Japan. This research was also partly supported by a Grant-in-Aid for General Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan and from the Japan Society for the Promotion of Science and by research grants for cardiovascular diseases from the Ministry of Health, Labor and Welfare (15C-5 and 16C-8). The authors thank the staff members of the Department of Nuclear Medicine and Central Institute of Isotope Science, Hokkaido University, and the Facility of Radiology, Hokkaido University Medical Hospital, for supporting this work. We also thank Theseus Imaging Corp. for providing annexin A5.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication February 18, 2008.
- Accepted for publication June 9, 2008.




{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Evaluation of Apoptosis with 99mTc-rhAnnexin V-128 and Inflammation with 18F-FDG in a Low-Dose Irradiation Model of Atherosclerosis in Apolipoprotein E-Deficient Mice
- Elevated 18F-FDG Levels in Blood and Organs After Angiotensin II Receptor Blocker Administration: Experiment in Mice Administered Telmisartan
- Determinants of FDG Uptake in Atherosclerosis
- The Year in Molecular Imaging
- Imaging Atherosclerotic Plaque Inflammation by Fluorodeoxyglucose With Positron Emission Tomography: Ready for Prime Time?
- Feasibility of FDG Imaging of the Coronary Arteries: Comparison Between Acute Coronary Syndrome and Stable Angina