


Abstract
Thyroid stunning refers to reduced uptake of 131I in the thyroid tissue (or tumor) during radioiodine (131I) therapy compared with the uptake measured after the previous administration of 131I for diagnostic purposes. The phenomenon is clinically important, as it can potentially lead to the undertreatment of thyroid cancer or to unnecessarily high absorbed doses in critical organs. Previous clinical and experimental studies indicated that thyroid stunning is absorbed dose dependent. The aim of this study was to investigate the effects of 131I irradiation on 125I− transport and cell proliferation at low absorbed doses in vitro. Methods: Primary cultured porcine thyroid cells were grown to form a confluent monolayer of epithelial cells on a filter in a bicameral culture system. The cells were continuously irradiated with 131I in the culture medium for 48 h to obtain 0.0015–1.5 Gy. At 3 d after irradiation was stopped, the transepithelial iodide transport capacity was evaluated by measuring 125I− transport from the basal chamber compartment to the apical chamber compartment. The effect of 131I irradiation on DNA synthesis was estimated by pulse labeling with 3H-thymidine of both subconfluent and confluent cells irradiated with up to 9 Gy. Total DNA content was measured to quantify cell numbers. Results: A statistically significant reduction in 125I− transport was seen at absorbed doses of ≥0.15 Gy, with a 50% reduction at 1.5 Gy, compared with the results observed for nonirradiated control cells. 3H-Thymidine incorporation was already statistically significantly reduced at absorbed doses of 0.01–0.1 Gy, but 0.15–0.3 Gy did not affect DNA synthesis. However, absorbed doses of ≥1 Gy again resulted in reduced DNA synthesis. A 50% reduction was obtained at 4 Gy. Total DNA measurements revealed a statistically significant reduction in cell numbers at 8 Gy. Conclusion: The lowest absorbed dose from 131I that reduced iodide transport was 0.15 Gy. Because stunning was found at low absorbed doses, it might occur for 131I treatment not only of thyroid cancer but also of thyrotoxicosis. On the basis of differences in dose responses, radiation-induced thyroid stunning and cell cycle arrest may be independent phenomena.
Thyroid stunning, implicating diminished uptake of a therapeutic amount of 131I compared with the uptake of a previously administered diagnostic amount of 131I, has primarily been studied in vivo, and there is controversy over whether this effect may influence the uptake of 131I during therapy (1–11). To what extent thyroid stunning may limit the efficacy of 131I therapy has not yet been investigated in a prospective randomized study. Previous studies from our laboratory revealed an absorbed-dose–dependent reduction in transepithelial iodide transport in 131I-irradiated cultured thyroid cells and revealed that 3 Gy (lowest absorbed dose studied) caused a 50% inhibition of 125I− transport compared with the results observed for nonirradiated control cells (9). In vitro data also indicated that the cell numbers in 131I-irradiated cultures did not change, indicating that stunning is not related to a loss of cells attributable to a therapeutic effect of 131I (9).
The precise mechanism of thyroid stunning is still unknown. It is conceivable, however, that the expression or function of iodide transport molecules, of which the sodium/iodide symporter (NIS) is the most important, is suppressed by 131I irradiation in some way. One possibility is that the stress response to ionizing radiation affects specialized and finely regulated cell functions, ultimately leading to reduced iodide uptake.
At present, however, there is no information on the lowest absorbed dose from 131I required to induce damage to thyroid cells that, on the one hand, is nonlethal (in the short-term perspective of days, as discussed here) but, on the other hand, may give rise to secondary changes that negatively influence iodide transport. The aim of this study was to investigate the effects of low absorbed doses from 131I on 125I− transport and cell proliferation in vitro.
MATERIALS AND METHODS
Cell Culture
Thyroid cells aggregated in small follicle segments were prepared from porcine thyroid glands obtained from the local abattoir as previously described (12). The cells were plated on a collagen-coated micropore filter (pore size, 0.4 μm) in a bicameral culture system (Transwell 3413; Corning Costar) and cultured at 37°C in an incubator with 5% CO2 and Earle's minimum essential medium supplemented with 5% fetal calf serum, penicillin (200 U/mL), streptomycin (200 μg/mL), and amphotericin B (Fungizone; 2.5 μg/mL) (all culture medium constituents were purchased from PAA Laboratories GmbH). The medium in the culture chambers was replaced every 2–3 d. Before irradiation, the cells were allowed to grow either for 1 d, resulting in highly proliferative subconfluent cells, or for 7 d, resulting in a growth-arrested cell monolayer with properties similar to those of the native thyroid follicular epithelium. Subconfluent cells were used to study cell proliferation, whereas experiments evaluating iodide transport were conducted with cells that were cultured to confluence and then growth arrested. In the model, the basal and apical compartments correspond to the extrafollicular space and the follicular lumen, respectively (Fig. 1). To assess confluence and the establishment of a tight cell monolayer, we monitored the barrier function by measuring transepithelial resistance with a Millicell ERS ohmmeter (Millipore Corp.).

Bicameral culture system. Pig thyrocytes were grown on micropore filter (pore size, 0.4 μm) that divided well into apical and basal compartments corresponding to follicular lumen and extrafollicular space, respectively.
Irradiation Procedures
Confluent Cultures.
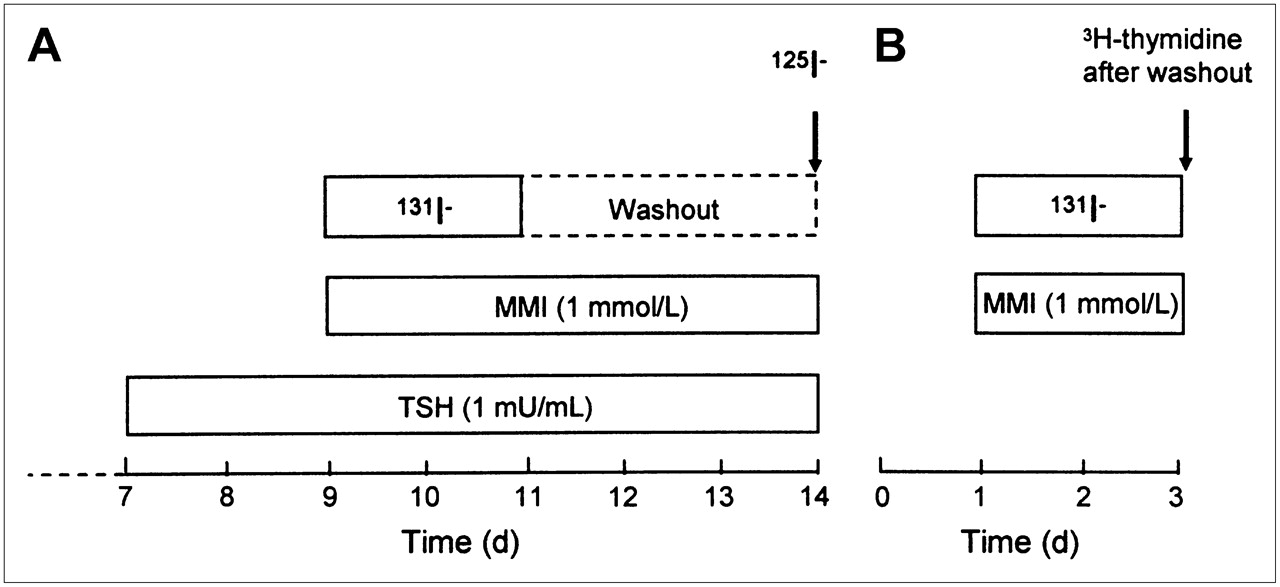
The experimental design is shown in Figure 2A. Confluent thyroid cell cultures were prestimulated with thyroid-stimulating hormone (TSH; 1 mU/mL; Sigma Chemical Co.) for 48 h before irradiation to upregulate NIS expression and the basal-to-apical transepithelial iodide transport capacity (9,12). TSH stimulation was continued throughout the experiment until iodide transport was evaluated. The culture medium in both the apical (200 μL) and the basal (400 μL) compartments was replaced with medium that had the same activity concentration of 131I. The cells were irradiated for 48 h at 37°C in 5% CO2. The amount of 131I was carefully adjusted to obtain mean absorbed doses to the cells of 0.0015–1.5 Gy (dosimetric calculations are given later in the article). Lead shields surrounded the plates to prevent external cross-irradiation between groups which are irradiated to different absorbed doses, and methimazole (MMI; 1 mmol/L; Sigma-Aldrich Sweden AB) was added to the medium to prevent protein binding of the 131I− and the Wolff–Chaikoff effect (if iodination is allowed, high iodide concentrations per se may downregulate NIS and thereby inhibit iodide uptake in thyroid cells) (13). Samples of stock solutions (medium containing 131I) were used to determine the activity concentration. After irradiation, samples of medium from both the apical and the basal compartments were used to study the redistribution of the activity between the compartments and to discover possible paracellular leakage in accidentally damaged cultures. The 131I activity in the samples was measured with a γ-counter (Wallac 1480 WIZARD 3; Wallac Oy). The 131I activity in the cultures was washed out by changing the medium at least 3 times.

Experimental protocols for confluent cells (A) and subconfluent cells (B). Cell culturing was started at time 0.
Subconfluent Cultures.
Subconfluent cells were used to study the effects of 131I irradiation on cell proliferation (Fig. 2B). The mean absorbed doses to subconfluent cells ranged from 0.01 to 9 Gy. The irradiation, washing, and sampling procedures were similar to those used for confluent cells. However, the subconfluent cells were not exposed to TSH, because such stimulation per se inhibits porcine thyroid cell growth in the model used (Mikael Nilsson, unpublished data, 2006). The 131I activity concentrations remained equal in the basal and apical compartments throughout the irradiation procedure because of free paracellular leakage. After irradiation, samples of medium from both the apical and the basal compartments were used to confirm equal activity concentrations.
Absorbed Dose Calculations
Monte Carlo (MC) simulations were used to estimate the absorbed doses to the cells from 131I distributed in the basal and apical compartments. MC simulations were performed by use of the PENELOPE code, with the main program PENCYL handling cylindric geometries (14). The geometry was in accordance with that shown in Figure 1 and was composed of cylinders. The PENELOPE code handles lower electron energies than do commonly used codes, and the low-energy cutoff is 100 eV. PENELOPE simulation results have been compared with experimental data from the literature, with the conclusion that there is good agreement in the energy range of a few kiloelectron volts to 1 GeV (15). Calculations were done separately for photons, β-particles, and atomic electrons, that is, conversion and Auger electrons, with 105–107 primary particles of each type. The full β-spectrum was used as input data (16). The average deposited energy per primary particle (electron or photon) was used to calculate the absorbed doses. Decay data for 131I were obtained from the Table of Radioactive Isotopes (16). In TSH-stimulated confluent cultures, a gradual change in activity concentrations between the apical and the basal compartments took place during the 48-h exposure because of basal-to-apical 131I transport (9). This effect was taken into account in the calculations for confluent cultures. Even though TSH stimulation increases the amount of 131I in thyrocytes (17), the absorbed doses to the cells should not be affected by the degree of TSH stimulation, because corrections were made for basal-to-apical transport and because the major portion of the absorbed doses was deposited by electrons emitted from 131I in the basal and apical media. The average ranges in water of the electrons were about 0.4 mm for the average energy (182 keV) and about 3 mm for the maximum energy (807 keV).
Iodide Transport Measurements
Transepithelial 125I− transport in the confluent cultures was assessed 3 d after the termination of 131I irradiation in a manner similar to that used in previous studies (9,12,18). A trace amount of 125I− (150 kBq/mL; Nycomed Amersham PLC) was added to the basal culture medium (400 μL; containing MMI at 1 mmol/L) of irradiated and corresponding nonirradiated control cell cultures. After 30 min, 100-μL samples were taken from both the apical and the basal media, and 125I activity was measured with the γ-counter. The relative amount of 125I− transported to the apical compartment was determined after correction for background, radioactive decay, and spillover from remaining trace amounts of 131I when necessary. The 131I-irradiated and nonirradiated control cell cultures were treated alike. Reproducible data were obtained from 3 separate experiments each comprising 6 cultures (bicameral cultures) per group.
3H-Thymidine Incorporation and DNA Quantification
Irradiated subconfluent cultures were analyzed for DNA synthesis and total cell numbers. In addition, nonirradiated cultures were analyzed in the same way every second day after plating until confluence was obtained. This procedure was done to be able to monitor time- and contact-dependent changes in cell proliferation. For both experimental situations, cells were incubated for 4 h with 3H-thymidine (37 Bq/mL; 925 GBq of 3H per millimole of thymidine; Nycomed Amersham PLC) added to both the apical and the basal compartments by medium exchange (200 and 400 μL, respectively). Incorporation was terminated by fixation of the cells with 10% trichloroacetic acid (Merck) for 1 h or more at 4°C, after which free 3H-thymidine was removed by excessive washes with the same solution. The insert filters with attached cells were cut out and solubilized with 500 μL of Soluene-350 (a mixture of toluene [60%–80%], dodecul(dimethyl)(tetradecyl)ammonium hydroxide [20%–40%], and methanol [2.5%–10%]) (Perkin-Elmer) in plastic scintillation vials (Packard Bioscience). After the addition of 10 mL of a liquid scintillation solution (Ultima Gold; Packard Bioscience), the activity of 3H in each sample was measured by use of a liquid scintillation detector (1217 Rackbeta; Wallac Oy) equipped with Utmac 1.1 (Wallac Oy) software. The experiments were reproducible and were performed with at least triplicate samples per group. The cell numbers in irradiated and nonirradiated cultures were estimated by fluorometric analysis of total DNA content (19).
Statistical Analysis
The Student t test was used to determine the statistical significance (P < 0.05) of differences between the experimental groups. Results are given as mean ± SEM.
RESULTS
As shown in Figure 3, the reduction in 125I− transport in confluent cells at 72 h after 131I irradiation was dependent on the absorbed dose from 131I. Cells that received 1.5 Gy showed a statistically significant reduction in transport of about 50% compared with nonirradiated control cells, and the reduction was about 20% at 0.15 Gy (P < 0.05). Absorbed doses of 0.015 Gy or lower had no statistically significant effect.
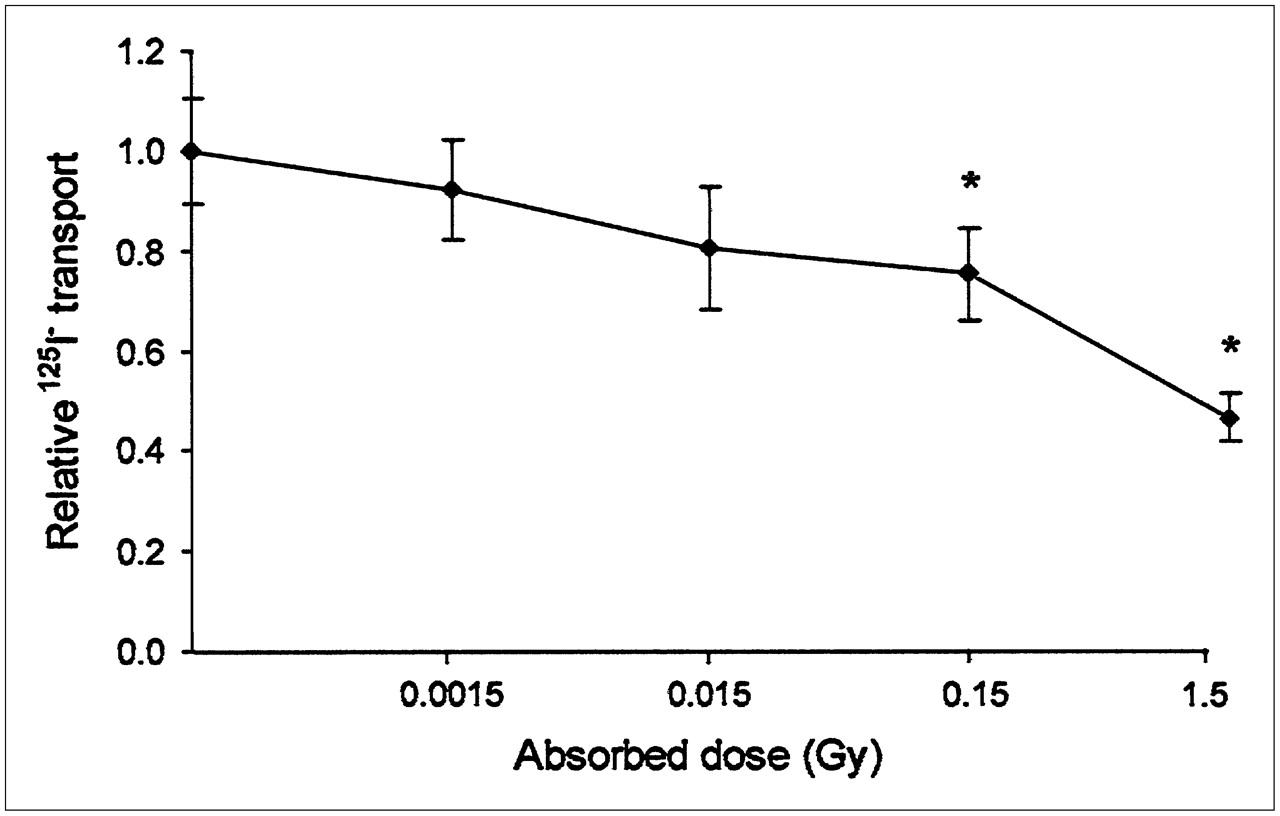
125I− transport through 131I-irradiated cells relative to nonirradiated cells. Relative 125I− transport is ratio of 125I− transported through irradiated cells to 125I− transported through nonirradiated control cells at different absorbed doses. * = data that were statistically significantly different from control data (P < 0.05). Data are given as mean ± SEM (n = 6).
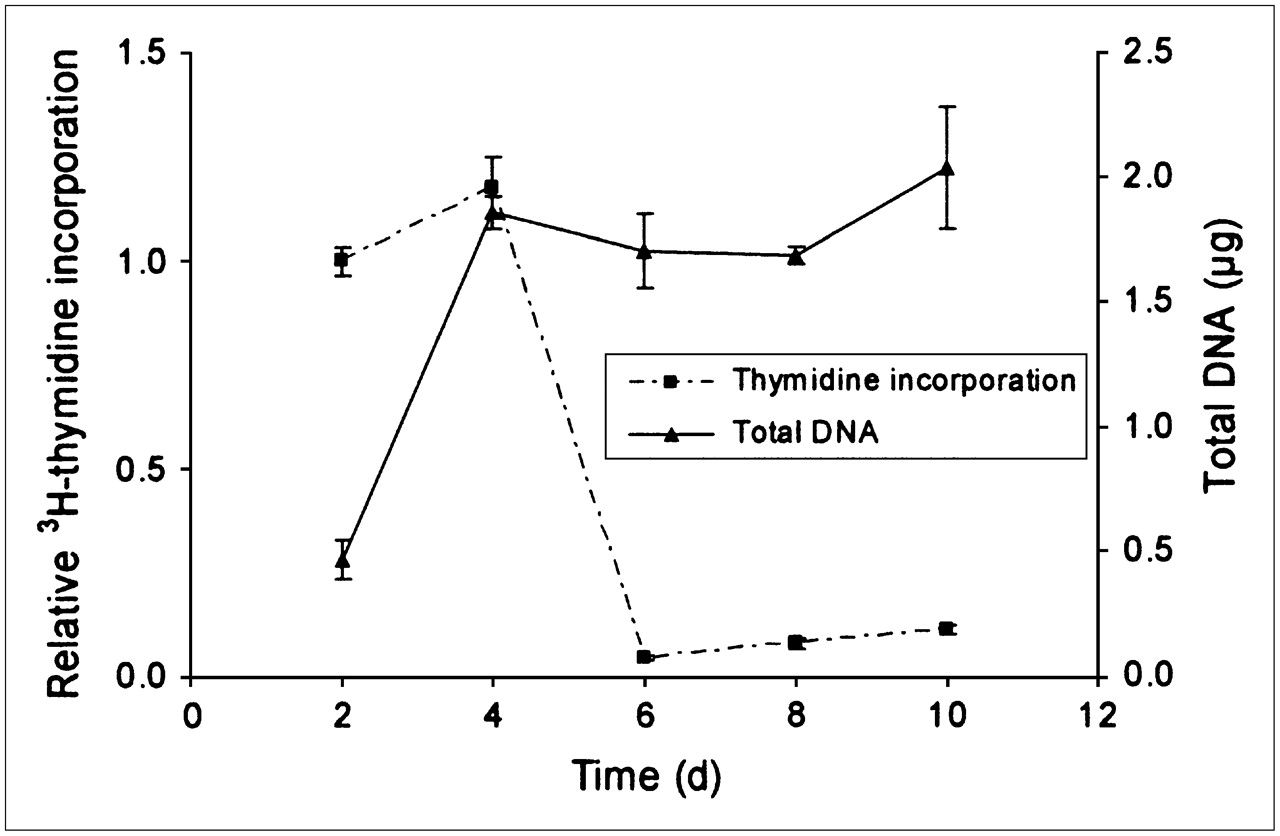
Kinetic analysis of DNA synthesis (3H-thymidine labeling) and total DNA content confirmed the notion that the cells ceased to proliferate once they became confluent after 4–6 d of culturing (Fig. 4). Therefore, the influence of 131I irradiation on growth could be examined only in subconfluent cells; for these cells, the results showed that low absorbed doses, 0.01 and 0.1 Gy, produced a statistically significant decrease in 3H-thymidine incorporation, whereas slightly higher absorbed doses, 0.15 and 0.3 Gy, produced no statistically significant change (Fig. 5). Absorbed doses of ≥1 Gy produced a more pronounced and dose-dependent reduction in the incorporation of 3H-thymidine. To obtain a reduction in proliferation of greater than 50%, an absorbed dose of about 4 Gy was required (Fig. 5). Quantification of total DNA content showed that the cell numbers did not change significantly in cultures subjected to absorbed doses of up to 3 Gy. An absorbed dose of 8 Gy, however, produced a statistically significant decrease in DNA content (Fig. 6). According to the results of MC simulations, about 98% of the absorbed dose to the cells was delivered by electrons, and only 2% was delivered by photons.

Relative 3H-thymidine incorporation normalized to first time point measured and total amount of DNA per sample in nonirradiated porcine thyrocytes at different times after start of culturing (day 0). 3H-Thymidine incorporation shows how cells initially proliferated, incorporating large amounts of thymidine, until they became confluent at about day 6. Total DNA content increased until confluence was reached. Data are given as mean ± SEM (n = 3).

Effect of 131I irradiation on subsequent 3H-thymidine incorporation. Results are given as 3H-thymidine incorporation in irradiated cultures relative to that in nonirradiated cultures. Hypersensitivity was obtained at 0.01 and 0.1 Gy (see inset). * = data that were statistically significantly different from control data (P < 0.05). Data are given as mean ± SEM (n = 3–21).
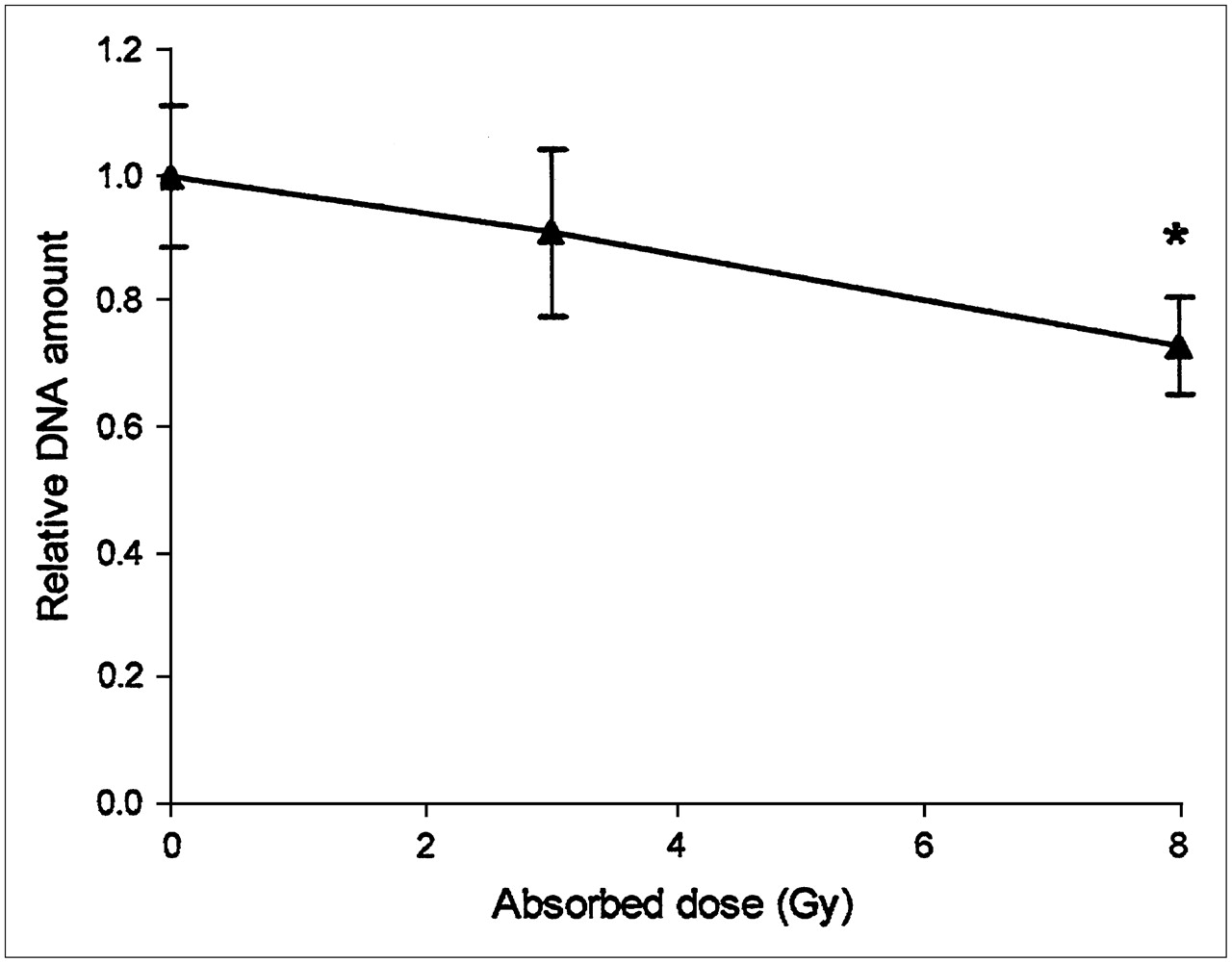
Total DNA per sample in 131I-irradiated cell cultures relative to that in nonirradiated cultures vs. absorbed doses to cells. * = data that were statistically significantly different from control data (P < 0.05). Data are given as mean ± SEM (n = 3).
DISCUSSION
Decreased thyroid iodide uptake after exposure to diagnostic test amounts of 131I is a clinically important problem demonstrated in patients with differentiated thyroid cancer. Thyroid stunning should be considered when radioiodine therapy is performed because it may affect the outcome of the therapy. However, the lowest absorbed dose required to produce stunning in vivo is not known. Thyroid carcinoma patients usually receive test amounts of 131I of about 20–200 MBq. Their thyroid primary tumors and regional lymph nodes have already been surgically removed, and only small remnants usually remain. Test amounts of up to 185 and 74 MBq have been estimated to deliver mean absorbed doses to thyroid remnants in the ranges of 10–58 and 4–38 Gy, respectively (1,10). Analyses of several in vivo and in vitro studies have confirmed that absorbed doses of ≥4 Gy may result in thyroid stunning (11). Nevertheless, in the present study, we showed that absorbed doses of ≥0.15 Gy produced a statistically significant reduction (20%) in iodide transport 3 d after the end of irradiation. A 50% reduction in iodide transport was obtained after irradiation with 1.5 Gy (Fig. 3).
We investigated whether the lowest absorbed dose that causes stunning also affects cell cycle checkpoints that are known to regulate the cell proliferation rate and that are activated in response to DNA damage (20). Repeated analyses of 3H-thymidine incorporation and total DNA content early after plating and onward showed that nonirradiated thyroid cells ceased to proliferate after 4–6 d, when they were crowded and contact inhibited, features designating the confluent state—that is, at the time when stunning was studied (Fig. 4). To be able to determine the growth-inhibitory response to irradiation, it was therefore necessary to use subconfluent cells, in which cell multiplication was fast and likely involved most of the cells. As already pointed out, reduced iodide transport was seen at a very low dose (0.15 Gy); in contrast, much higher absorbed doses (>3 Gy) were required to affect the proliferation rate and reduce DNA synthesis by 50% in subconfluent cells (Fig. 5). These data, together with the fact that stunning was documented in confluent and growth-arrested cells, suggest that 131I irradiation likely inhibits iodide transport independently of its action on cell cycle control mechanisms. This notion is further emphasized by the fact that thyroid stunning, as observed with the present experimental model, can readily be induced by 131I in growth-arrested G0 cells.
Subconfluent cultures receiving 0.01–0.1 Gy were found to incorporate 10%–15% less 3H-thymidine than corresponding nonirradiated cultures. This finding may correspond to earlier documented curve forms seen for several human cancer cell lines exposed to an x-ray beam in vitro in studies with cell survival as the end point; hyperradiosensitivity was observed at absorbed doses of less than ∼0.5 Gy (21–24). This phenomenon has also been shown during external radiation therapy in vivo for both mice (skin, lungs, and kidneys) and human skin (24). However, it should be emphasized that altered nucleotide transport may be an alternative mechanism explaining small changes in 3H-thymidine incorporation.
Postgård et al. (9) previously demonstrated thyroid stunning in vitro with the same cell culture system as that used in the present study. The irradiation geometry used in the present study was slightly different from that used in the preceding study with regard to the volumes in the apical and basal compartments. Because of the different geometries, the dosimetric correction factor for the gradual accumulation of 131I− in the apical medium used by Postgård et al. (9) was 2.7, and the corresponding value in the present study was 1.5. Furthermore, in the present study, an MC code was used to simulate the average deposited energy in the cell layer. The results of the MC simulations and the method used by Postgård et al. (9) for calculations of the absorbed doses to thyrocytes were similar (i.e., 1 Gy obtained by the method of Postgård et al. and 1 Gy calculated from the MC simulations). The maximum total uncertainty in the determination of the absorbed dose was estimated to be ±30% and was mainly attributable to variations in the rate of transport of 131I (and thus different activity concentrations over time in the apical and basal compartments). However, MC simulations were performed with a high statistical certainty and a large number of events.
A homogeneous dose distribution is expected at absorbed doses that are not too low. With the present experimental model, we can only determine the total iodide transport by all cells and not distinguish radiobiologic effects of 131I on individual cells. Therefore, a microdosimetric approach was not needed with this model even for the lowest absorbed dose studied, at which heterogeneous cellular energy might be expected.
Thus, the use of different dose calculation methods is not responsible for the discrepancy in the results of the 2 studies; in the study of Postgård et al. (9), the absorbed dose that produced a 50% reduction in iodide transport was 3 Gy (9), whereas in the present study, this dose was 1.5 Gy. This discrepancy is most likely attributable to physiologic variations between primary cell cultures. The findings in the 2 studies concerning significant reductions in the amounts of DNA (significant reduction at 8 Gy in the present study but nonsignificant reduction at 13.5 Gy in the study of Postgård et al. (9) with confluent cells) are not directly comparable. In the present study, DNA measurements were performed on highly proliferative subconfluent cells to avoid alterations in radiosensitivity that may occur when nonproliferative cells are used (25).
Previous studies revealed an early increase in iodide transport/uptake after irradiation (26,27). Meller et al. reported increased iodide uptake in 2 thyroid cell lines at various stages of differentiation up to 72 h after external irradiation (26). A similar time dependence was reported by Postgård et al. (27); at times of less than 48 h after 131I irradiation of primary cultured pig thyrocytes, there was an increase in iodide transport, which was followed by stunning (27). These findings indicate that the time elapsed between diagnostic administration and therapeutic administration of 131I is crucial when the stunning phenomenon is being studied.
The cell culture system used in the present study seems to be a good model for the in vivo situation, mimicking the basal extrafollicular space, the apical lumen, and the cell layer with different compartments. The in vitro model is designed for studies of natural transepithelial iodide transport through cells involving, for example, the NIS. However, there are important differences. In the in vitro model, there are no closed follicles, and there is no or very little iodination of thyroglobulin because of the effect of MMI. In the in vitro model, there is a constant TSH level, but this level varies both with time and among individuals in clinical cases. On the other hand, in thyroid tumor cells that might be affected by stunning, both follicle architecture and iodination capacity have often deteriorated. Although experimental data always should be carefully interpreted, we believe that the main results obtained with this model for studying the stunning phenomenon are valid for differentiated normal thyroid cells. The data should therefore be valuable in attempts to understand the clinical situation.
Beside reducing iodide transport, stunning can also reduce the effective half-life of iodine in thyroid or tumor tissue. Stunning may be attributable to early effects of the therapeutic administration of 131I. However, in our in vitro studies, stunning occurred solely as a result of the test administration of 131I. The stunning effect in vivo may be an additive effect of both test administration and therapeutic administration of 131I. If that is the case, then the stunning effect is underestimated with our model. Other parameters that may affect stunning are linear energy transfer and absorbed dose rate. However, to our knowledge, studies of these mechanisms have not yet been presented. Studies to elucidate the dependence of thyroid stunning on absorbed dose rate and linear energy transfer are ongoing.
Although it is advisable to be careful in extrapolating in vitro data to the clinical situation, the findings of the present study suggest that thyroid stunning may also be a problem in the management of benign thyroid disease. From a test activity of 0.5 MBq, a typical patient with Graves' disease will receive absorbed doses to the thyroid gland of approximately 0.1–0.5 Gy (28), absorbed doses shown to cause stunning in the present study.
Several differences between the in vitro and the in vivo situations, such as different concentrations of TSH and different timing, make it necessary to perform similar studies in vivo before it can be conclusively stated how the stunning phenomenon occurs in the clinical situation.
CONCLUSION
The findings of the present study indicate that thyroid stunning, that is, 131I-induced inhibition of iodide uptake, is significant at absorbed doses as low as 0.15 Gy in an in vitro model mimicking normal thyroid tissue compartments. These data suggest that stunning may occur with 131I therapy not only of thyroid cancer but also of benign thyroid disease. On the basis of differences in responses to absorbed doses, thyroid stunning and cell cycle arrest may be independent phenomena.
Acknowledgments
The authors would like to acknowledge Bertil Arvidsson, Pernilla Jonasson, Stig Palm, and Agneta Lundström for their help with supplying 131I. We are also thankful to Therese Carlsson, whose technical expertise is highly appreciated. This study was supported by grants from the Swedish Cancer Society (grants 3427 and 4567), the Swedish Radiation Protection Authority, and the King Gustav V Jubilee Clinic Cancer Research Foundation.
Footnotes
-
↵* Contributed equally to this work.
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication June 20, 2006.
- Accepted for publication November 27, 2006.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The effect of prior antithyroid drug use on delaying remission in high uptake Graves' disease following radioiodine ablation
- Radiation-Induced Thyroid Stunning: Differential Effects of 123I, 131I, 99mTc, and 211At on Iodide Transport and NIS mRNA Expression in Cultured Thyroid Cells
- Down-regulation of the Sodium/Iodide Symporter Explains 131I-Induced Thyroid Stunning