


Abstract
The study of the effects of sex and hormones on brain chemistry and neurotransmission is of increasing importance as evidence emerges of sex differences in behavioral symptoms and treatment response in neuropsychiatric disorders. The nicotinic acetylcholine receptor (nAChR) system has been implicated in a variety of psychiatric disorders, including tobacco smoking, for which there is strong evidence supporting sex differences in behaviors and response to smoking cessation treatments. We examined the availability of nAChR containing the β2 subunit in healthy men and women and the influence of menstrual phase among women. Methods: Ten men and 19 women nonsmokers underwent one 123I-5-IA-85380 (123I-5-IA) SPECT scan and one MRI scan. A subset of 9 women, aged 18–39 y, underwent a second 123I-5-IA scan. These 9 women were scanned during the early follicular (days 4–7 in 8 subjects and day 11 in 1 subject) and mid-luteal (days 19–25) phases of their menstrual cycle. Hormone levels were measured in all women to confirm the phase of the cycle. Results: Regional brain activity (kBq/cm3) was higher (39%–54%) in women than in men nonsmokers. When regional brain activity was normalized to total plasma parent to correct for individual differences in radiotracer metabolism (VT′), differences of 10%–16% were observed, with women greater than men. In contrast, when regional brain activity was normalized to free plasma parent (VT), there was less than a 4% difference by sex in regional brain β2-nAChR availability. These sex differences in kBq/cm3 and VT′ resulted from significantly higher levels of total plasma parent, free fraction (f1), and free plasma parent in women than in men nonsmokers. No differences in plasma measures or brain β2-nAChR availability were observed across the menstrual cycle for any outcome measure. Conclusion: Overall, these findings demonstrate no significant difference in brain β2-nAChR availability between men and women nonsmokers or across the menstrual cycle. Importantly, these findings demonstrate sex differences in radiotracer metabolism and plasma protein binding and highlight the critical need to measure plasma radiotracer levels and f1 in studies that include both sexes.
The importance of studying sex differences in brain neurochemistry has become increasingly evident with the recent demonstration of differences between men and women in symptomatology and treatment for many neuropsychiatric disorders. Evidence of differences in brain structure, function, neurotransmission, and receptor availability between men and women is mounting (1). Sex differences exist in a variety of behaviors related to tobacco smoking, such as craving, sensitivity to nicotine, and response to nicotine replacement therapy (2,3); however, the mechanisms underlying these differences are unclear. Women often experience more difficulty in quitting smoking than men (4–9), and rates of success may differ by menstrual phase. For example, women who quit smoking during the luteal phase of the menstrual cycle reported significantly more tobacco withdrawal and depressive symptoms than did women who quit smoking during the follicular phase, and this effect was related to the attempt to quit, not to a worsening of mood due to phase of cycle (10). These behavioral differences may be due to underlying sex differences in brain chemistry. Several preclinical studies suggest sex differences at the receptor level. The nicotinic acetylcholine receptor (nAChR) containing the β2 subunit (β2-nAChR), one of the initial sites of action of nicotine in the brain and the most critical site for the reinforcing effects of nicotine, is a likely neurochemical substrate mediating these sex differences. Preclinical studies in nicotine-naïve rats (11) and mice (12) suggested higher nAChR numbers in female than in male rats. It is not known whether nicotinic receptor availability differs between living men and women.
Menstrual cycle–related fluctuations in the neuroendocrine milieu may contribute to the observed sex differences in smoking-related behaviors. At the receptor level, the actions of sex steroids on nAChRs are complex, and preclinical evidence indicates that progesterone and estrogen have opposing effects on nAChR. Progesterone (13–15) and its A-ring metabolites (14) noncompetitively inhibit nAChR. Progesterone (16) and the neurosteroid pregnenolone (17) also have been shown to block nicotinic receptor response. Conversely, estradiol has been shown to potentiate acetylcholine-evoked currents in human neuronal nAChRs by interacting at the α4 subunit (18). However, the steroid hormone β-estradiol and others also inhibit acetylcholine-induced currents in rat neurons (19). Thus, there appears to be a relationship between steroid hormones and nAChR numbers. Ovariectomy also reduces coupled acetylcholine synthesis and high-affinity choline uptake, which is reversed by the administration of estradiol in rat brain (20). The effect of estradiol on the state of neuronal nAChRs has also been studied. Chronic estradiol treatment (i.e., 2 mo) in female rats decreases the affinity and increases the binding of 3H-methylcarbamylcholine, a nicotinic agonist, in the hippocampus and hypothalamus, suggesting that estradiol decreases presynaptic cholinergic function (21).
The development and validation of 123I-5-IA-85380 (123I-5-IA) for brain SPECT allows the in vivo examination of β2-nAChRs in living men and women. High test–retest reliability and validity were recently reported for 123I-5-IA SPECT (22), and 123I-5-IA SPECT has demonstrated significantly higher β2-nAChR availability in recently abstinent tobacco smokers than in age- and sex-matched nonsmokers (23). In the present study, we hypothesized that the sex and cyclical differences in response to nicotine are due to variations in nAChR availability in the brain. This question was addressed by imaging healthy nonsmoking men and women with 123I-5-IA SPECT to examine the effects of sex and menstrual cycle in women on the availability of β2-nAChRs.
MATERIALS AND METHODS
Subjects
Study 1: The Effect of Sex on 123I-5-IA Uptake.
Ten men (mean age ± SD, 27.7 ± 7.3 y; range, 20–41 y; 10 Caucasian) and 19 women (mean age, 26.2 ± 6.8 y; range, 20–39 y; 9 Caucasian, 5 Hispanic, 4 African American, and 1 Asian) underwent one 123I-5-IA scan and one MRI scan. Women in study 1 who did not participate in study 2 were scanned during the follicular (n = 6) or mid-luteal (n = 3) phase of the menstrual cycle, and 1 woman who was taking hormonal contraceptives at the time of the scan was included. Hormones were analyzed to confirm the phase of the cycle and the premenopausal status.
Study 2: The Effect of Menstrual Cycle on 123I-5-IA Uptake.
Nine women (mean age, 27.0 ± 7.8 y; range, 18–39 y; 4 white, 2 Hispanic, 2 African American, and 1 Asian) who participated in study 1 were recruited to participate in study 2, which involved undergoing a second 123I-5-IA scan. With the first day of menstrual flow anchored as day 1, SPECT scans were performed during the early follicular (days 4–7 in 8 subjects and day 11 in 1 subject) and mid-luteal (days 19–25) phases of the menstrual cycle. The subjects were not counterbalanced for the order of follicular versus mid-luteal scans. Of the 9 subjects, 8 completed the 2 scans within 2 consecutive cycles, and 1 completed the scans within 3 consecutive cycles. Hormone levels were measured on scan days to confirm the phase of the cycle. We used estradiol levels of less than 55 pg/mL during the early follicular phase and progesterone levels of more than 3 ng/mL during the mid-luteal phase to confirm cycle phase.
This study was approved by the Yale University School of Medicine Human Investigation Committee, the West Haven Veterans Administration Human Subjects Subcommittee, the Radiation Safety Committee, and the Food and Drug Administration. Subjects were recruited from the community by word of mouth, posters, or newspaper advertisements. Eligibility was determined as follows. All subjects received a medical examination by a study physician to exclude any major medical issues or neurologic disorders. This medical examination included a physical examination, electrocardiogram, serum chemistries, thyroid function studies, complete blood count, urinalysis, and urine toxicology screening. The subjects were interviewed using the Structured Clinical Interview of the Diagnostic and Statistical Manual of Mental Disorders to rule out any Axis I disorder. All subjects were never smokers (defined as <100 cigarettes in a lifetime) and had no history of significant medical illness or major head trauma. Nonsmoking status was confirmed by plasma cotinine levels of less than 15 ng/mL, urine cotinine levels of less than 100 ng/mL and carbon monoxide levels of less than 11 ppm on the day of intake and the day of the scan. All women of childbearing age were required to have a negative pregnancy test during the screening process and before radiotracer injection on each study day. All women participating in study 2 were required to have no gynecologic problems and have a history of regular menstrual cycles 26–32 d in length. In the menstrual cycle study, use of hormonal contraceptives within the previous 6 mo was exclusionary.
MRI
MRI was performed on a 1.5-T camera (Sonata; Siemens) in a standard orientation (5- to 7-ms echo time, 24-ms repetition time, 256 × 192 matrix, 1 excitation, 30-cm field of view, and 124 contiguous slices with a 1.2-mm thickness), and the MRI images were used for coregistration to the SPECT images to provide an anatomic guide for placement of the regions of interest.
123I-5-IA SPECT Scan
All subjects received a 0.6-g saturated solution of potassium iodide in the hour before radiotracer administration. 123I-5-IA was synthesized as previously described (24) and administered as a bolus to constant infusion at a ratio of 7.0 for 8 h. In study 1, men were injected with a bolus of 150.6 ± 23.6 MBq and a continuous infusion of 21.6 ± 3.4 MBq/h and women with a bolus of 152.6 ± 18.5 MBq and a continuous infusion of 22.1 ± 2.9 MBq/h. Thus, the total dose was 345.1 ± 54.3 MBq for men and 349.6 ± 41.2 MBq for women. In study 2, women were injected in the follicular phase with a bolus of 146.8 ± 26.2 MBq and a continuous infusion of 22.4 ± 2.7 MBq/h and in the mid-luteal phase with a bolus of 160.7 ± 8.1 MBq and a continuous infusion of 23.3 ± 0.06 MBq/h. Thus, the total dose was 333.5 ± 56.6 MBq in the follicular phase and 363.8 ± 15.6 MBq in the mid-luteal phase. Three consecutive 30-min emission scans and one 15-min simultaneous transmission and emission scan were obtained between hours 6 and 8 on a PRISM 3000 XP (Picker) SPECT camera. The PRISM 3000 XP is a 3-head camera equipped with a low-energy, ultra-high-resolution fanbeam collimator (photopeak window, 159 keV ± 10%; matrix, 128 × 128) with a uniform sensitivity across the field of view. A 57Co-distributed source was measured with each experiment to control for day-to-day variation in camera sensitivity. The axial resolution (full width at half maximum) was 12.2 mm, measured with a 123I line source in water in a cylindric phantom. Blood was drawn before injection and at the beginning and end of the emission scans for analysis of plasma total parent and free fraction (f1) of parent tracer in plasma.
Image Analysis and Outcome Measures
Images were reconstructed and analyzed as previously described, including nonuniform attenuation correction (22), with a single exception. Specifically, the second SPECT scan for the subjects in study 2 was coregistered to the same position as the first scan in order to apply the same region-of-interest template for each subject. MR images were coregistered to the SPECT images to provide an anatomic guide for placement of the regions of interest using MEDx software (Medical Numerics, Inc.). The chosen regions of interest were those known to contain β2-nAChRs and included the frontal, parietal, anterior cingulate, temporal, and occipital cortices; the thalamus, the striatum (an average of caudate and putamen), and the cerebellum. Two raters conducted the analysis. Variability between the raters was less than 12% across regions of interest. The mean of the analysis from the 2 raters is reported.
123I-5IA regional activity (kBq/cm3) is reported along with the outcome measures VT′ (regional activity divided by total plasma parent between 6 and 8 h), which has shown the best test–retest reliability for 123I-5-IA SPECT (25), and VT (regional activity divided by free plasma parent between 6 and 8 h), to correct for possible differences in plasma protein binding. Both VT′ and VT are proportional to the binding potential (binding potential, in mL/g, equals receptor number divided by affinity), which is proportional to the receptor number at equilibrium, given the assumptions that there is no change in affinity and that nondisplaceable (nonspecific and free) uptake does not differ between subjects or between comparison groups. As described previously (25), there is no appropriate reference region for this radiotracer, so nondisplaceable 123I-5-IA uptake could not be measured. Also, to control for differences in metabolism and protein binding, the measures of total plasma parent, f1, and free parent, which is defined as total parent × f1, were compared between groups.
Statistical Analysis
For study 1, preliminary analyses of kBq/cm3, VT′, and VT revealed that values for these measures were highly correlated between brain regions. Pairwise correlations between brain regions ranged from 0.67 to 0.95 for VT′, from 0.77 to 0.96 for VT, and from 0.89 to 0.99 for kBq/cm3. Therefore, to obviate issues of multicollinearity in the interpretation of these data, measures of kBq/cm3, VT′, and VT across brain regions were each reduced to single, main components using principal-component analysis. A contemporary, technical account of principal-component analysis has previously been reported (26). Sex differences in these principal components representing more global measures of kBq/cm3, VT′, and VT across brain regions were then examined using unpaired t tests. Bonferroni correction for multiple comparisons across 8 brain regions indicated that P values of less than 0.00625 were highly significant. Because of the high correlations between brain regions, the P values in Table 1 cannot be treated as independent. For study 2, differences in VT′ and VT between the early follicular and mid-luteal phases were examined with paired t tests.
Sex Differences Between Men and Women in 123I-5-IA Activity by Outcome Measure
RESULTS
Study 1: The Effect of Sex
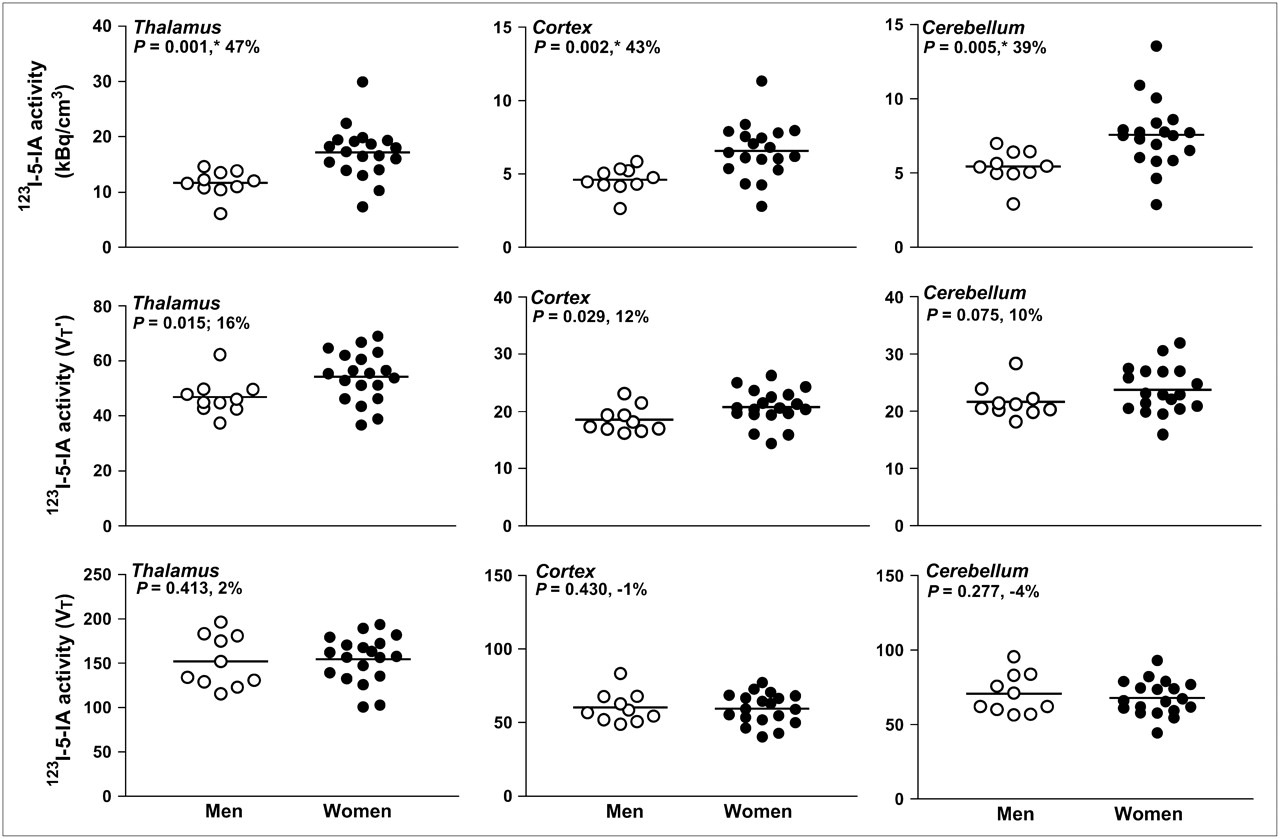
Based on principal-component analyses, single components explained 96% of the variance in kBq/cm3, 88% of the variance in VT′, and 91% of the variance in VT across brain regions. Sex differences in 123I-5-IA uptake varied by outcome measure (Table 1; Figs. 1 and 2). Across brain regions, the main kBq/cm3 and VT′ components were significantly greater in women than in men (for kBq/cm3: t27 = 4.02, P = 0.004; for VT′: t27 = 2.20, P = 0.0362). The main VT component did not differ significantly between women and men.

123I-5-IA activity is shown in 10 men (○) and 19 women (•) in 3 selected brain regions: thalamus, cortex (average of cortical regions), and cerebellum. Top row depicts data as kBq/cm3 uncorrected for metabolism or protein binding, middle row is VT′, and bottom row is VT.*P values < 0.00625 are significant after Bonferroni correction.
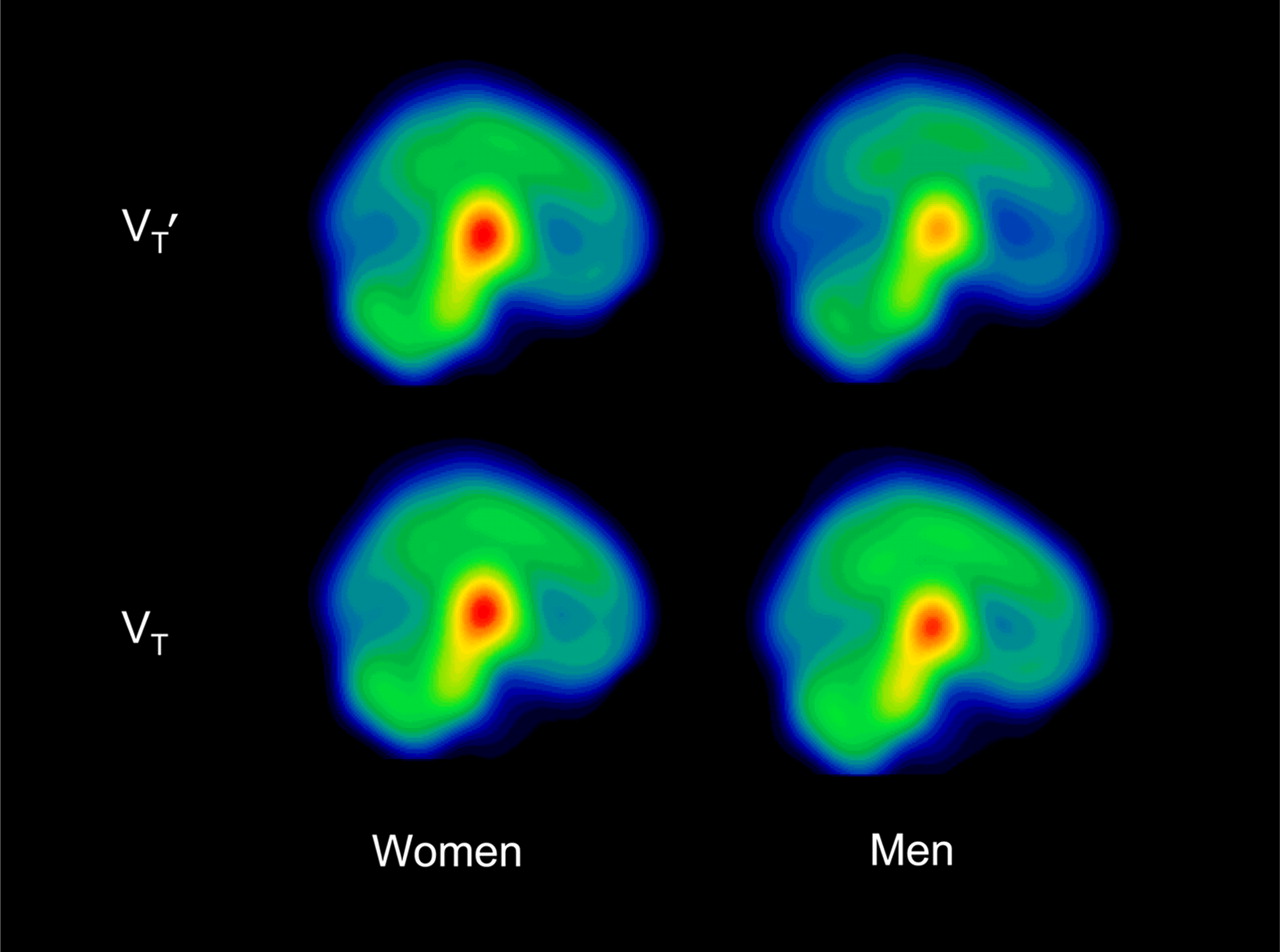
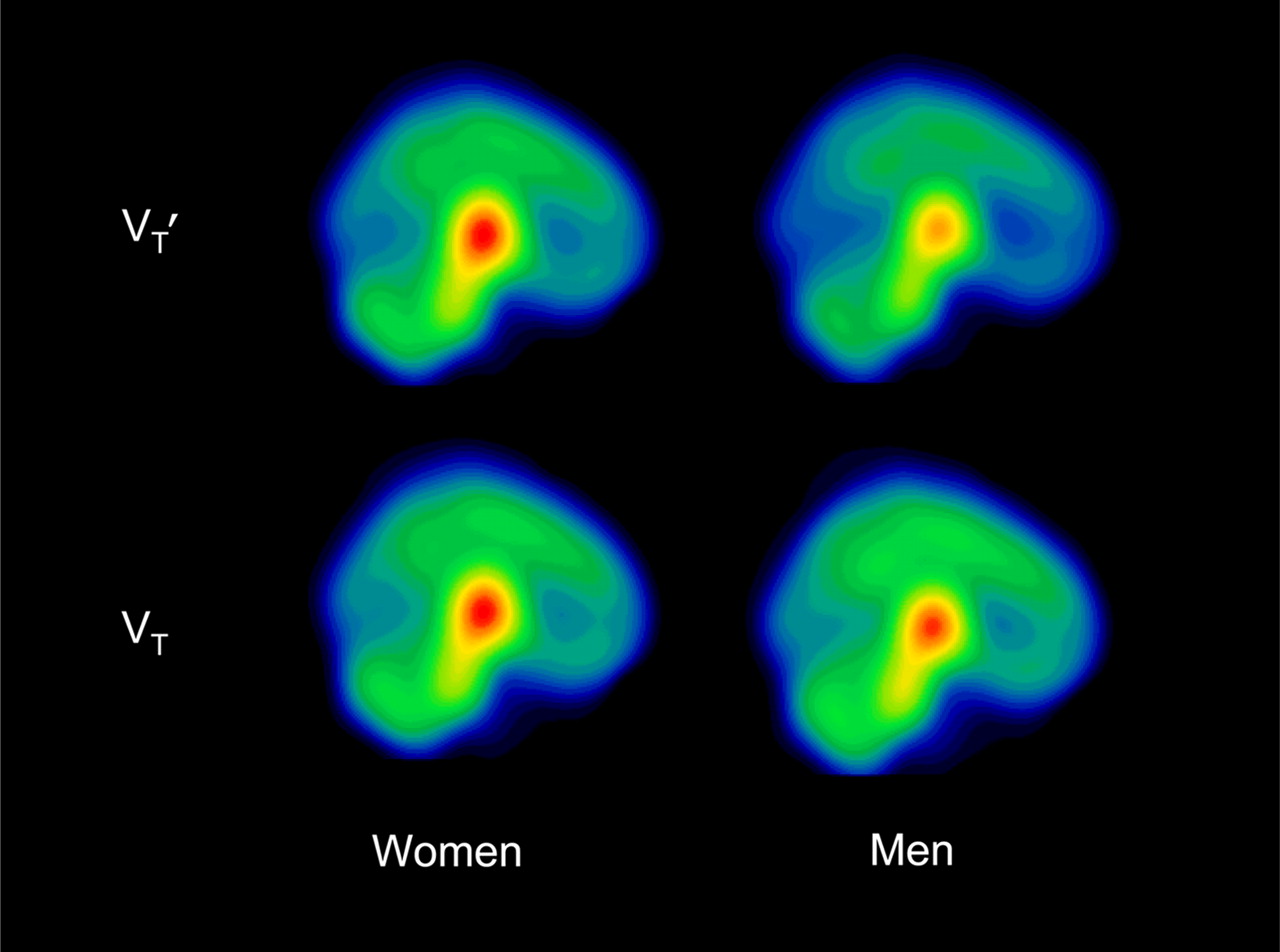
Mean parametric images illustrating 123I-5-IA activity in 10 men and 19 women in VT′ and VT.
Statistical analyses (t test) also determined that women have significantly greater f1 (t27 = 2.50, P = 0.02), total parent (t27 = 2.03, P = 0.05), and plasma free parent (t27 = 2.05, P = 0.05) than do men (Fig. 3).

Radiotracer metabolism and protein binding are shown in 10 men (○) and 19 women (•).
Study 2: The Effect of Menstrual Cycle
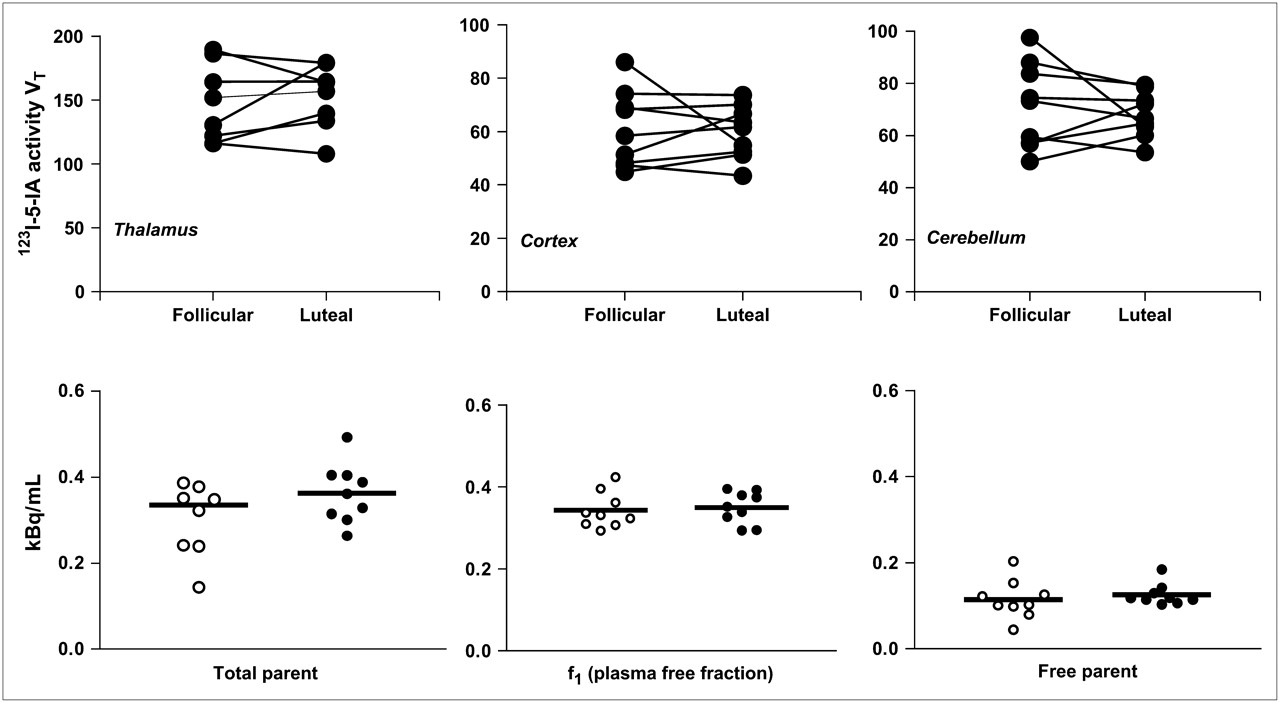
Hormone levels obtained on each SPECT scan day during the early follicular (mean estrogen, 38.1 pg/mL; mean progesterone, 0.7 ng/mL) and mid-luteal (mean estrogen, 121.1 pg/mL; mean progesterone, 6.4 ng/mL) phases confirmed the accuracy of the self-reported phase of the cycle. However, although all subjects evidenced a luteal-phase rise in progesterone, 4 subjects did not have luteal-phase progesterone levels greater than 3 ng/mL, suggesting an anovulatory cycle as occurs in as many as 38% of cycles in young women (27) and is consistent with other studies (28). Results did not differ when these subjects were eliminated from the analyses. Radiotracer metabolism and plasma protein binding—for example, total parent and f1—did not change between the early follicular and mid-luteal phases of the menstrual cycle (Fig. 4). No significant differences in VT′ were observed in women between the early follicular and mid-luteal phases in any region of interest (thalamus: t = 0.64, P = 0.54; cortex: t = 0.48, P = 0.64; striatum: t = 0.93, P = 0.38, and cerebellum: t = 1.62, P = 0.14 [data not shown]). Similarly, no significant differences in VT were observed (thalamus: t = 0.35, P = 0.74; cortex: t = 0.26, P = 0.80; striatum: t = 0.39, P = 0.71 [data for striatum not shown in Fig. 4]; and cerebellum: t = 0.66, P = 0.53) [Fig. 4]). Additionally, there were no correlations between estrogen or progesterone obtained on scan days and β2-nAChR availability using the outcome measure VT′ or VT.

(Top) Within-subject 123I-5-IA activity (VT) in thalamus, cortex (average of cortical regions), and cerebellum in 9 nonsmoking women in early follicular and mid-luteal phases of menstrual cycle. (Bottom) Radiotracer metabolism and protein binding in same women in early follicular (○) and mid-luteal (•) phases of menstrual cycle. No significant differences in VT, total parent, f1, or free parent were found between early follicular and mid-luteal phases of menstrual cycle.
DISCUSSION
This SPECT study examined the effect of sex and menstrual cycle phase on regional brain uptake of 123I-5-IA. The most notable finding was the statistically significant sex differences in total parent (unmetabolized 123I-5-IA) and in f1, suggesting that the radiotracer 123I-5-IA is metabolized differently between men and women and that the amount of 123I-5-IA bound to plasma proteins differs between men and women. Importantly, if not corrected for in the brain outcome measures, these differences may lead to the appearance of a sex difference in nAChR availability. This appearance was clearly demonstrated in the present dataset by the profound sex difference in uncorrected (kBq/cm3) measures of regional brain uptake (39%–54%), as compared with the outcome measure that corrected for metabolism only (but not protein binding), VT′ (10%–16%).
A higher f1 is associated with a higher radiotracer uptake by the brain (29); thus, because VT′ does not correct for protein binding, the sex difference is due to a higher 123I-5-IA uptake in women than in men but not to a greater β2-nAChR availability. Accordingly, there was no difference in β2-nAChR availability as evaluated by the outcome measure (VT) that corrected for both radiotracer metabolism and the f1 of radiotracer. Radiotracer metabolism, plasma protein binding, and brain β2-nAChR availability did not differ between the early follicular and mid-luteal phases of the menstrual cycle in women nonsmokers.
The Effect of Sex on β2-nAChR Availability
This study found no difference in β2-nAChR availability between healthy men and women nonsmokers. Preclinical studies indicated that nicotine-naïve female animals had higher global nAChR numbers than did male animals (30,31); however, this finding could not be generalized to humans. Differences between species and between radioligands may account for the discrepancy. Differences in nAChR distribution between species have been reported (32); and the radioligands used, 3H-cytisine and (±)-exo-2-(2-123I-iodo-5-pyridyl)-7-azabicyclo[2.2.1]heptane), an analog of epibatidine, do not selectively measure the β2 subtype of nAChR. Additionally, it is likely that radioligand metabolism or protein binding were different between male and female animals. Unfortunately, the small size and blood volume of mice and rats precludes appropriate plasma analysis.
In an earlier preliminary analysis of 12 subjects (6 men and 6 women), we reported a sex difference in β2-nAChR availability (33). These analyses were based on the outcome measure VT′ because the difference in total parent and f1 was not statistically different and because VT′ had been shown to be a more reliable and less variable outcome measure than VT (34). As reported here, when the sample size increased to 29 (19 women and 10 men), the sex difference in f1 and total parent became significant and a contributing factor to the observed sex difference in brain β2-nAChR availability. Specifically, in this study, significant sex differences were found, with higher f1 and total and free parent in women than in men. The significant difference in f1 led us to evaluate the data with the outcome measure VT, which indicated that there is no sex difference in β2-nAChR availability between nonsmoking men and women.
A limitation of many neuroreceptor imaging studies is that f1 often is not measured or reported. It is therefore difficult to determine whether apparent differences in receptor availability between groups may actually be due to a difference in available unbound radiotracer in plasma. Some sex differences in the metabolism of radiotracers do exist. For example, men had a significantly greater percentage of unmetabolized 18F-altanserin in plasma, and thus, women metabolize 18F-altanserin faster than men (35), which led to potentially discrepant findings regarding sex differences in the serotonin 2A receptor (35,36). Specifically, the study that reported sex differences (36) did not correct for metabolism of 18F-altanserin, and when metabolism was accounted for, no sex difference was found (35). No sex difference in f1 between men and women was specifically reported for 18F-altanserin (35) and for 11C-WAY-100635 (37). The reason for a sex difference in f1 in this study is unclear. In humans, both age and sex influence cytochrome p450 enzymatic activity. For example, sex and menstrual phase influence cytochrome p450 3A activity, with women having higher activity than men, specifically in the preovulatory phase (38); however, it has been suggested that the effect of sex is more limited than the effect of intersubject variation (39). There may be a sex difference in protein binding, or it may be that sex-steroid hormones in women influence protein binding. One study reported that young adult women have lower basal protein oxidation than men (40), but no sex differences were reported for protein turnover (40), plasma protein secretory rates (40), or albumin concentrations (41,42). There was no evidence in the current study that estrogen or progesterone levels were associated with f1.
One limitation of this study was that we could not determine the effect of race on β2-nAChR availability. There are differences in the metabolism of nicotine both by race (43) and by sex (44,45); thus, it is possible that race may also affect the metabolism and plasma protein binding of 123I-5-IA. Unfortunately, a stringent analysis of race requires a larger sample and is beyond the scope of this study.
Another limitation of this study was that we examined men and women nonsmokers under the age of 45 y. Thus, we do not know yet if the sex differences in metabolism and protein binding of the radiotracer persist across the life span. We chose this age range to focus on a population of premenopausal women, because we had hypothesized that the sex differences in β2-nAChR availability were due to the sex-steroid hormones β-estradiol and progesterone, which vary over the course of the menstrual cycle. Although a finding of no sex difference in β2-nAChR availability in nonsmokers does not preclude a sex difference in β2-nAChR availability in smokers, it does raise the question of whether availability of β2-nAChRs is involved in the sex differences in tobacco-smoking behavior, such as differences in quitting smoking and in response to nicotine replacement therapy. Although 123I-5-IA SPECT measures the availability of β2-nAChR, this measure may be confounded by occupancy of the receptor by endogenous acetylcholine (46). However, no reports of sex differences in endogenous acetylcholine levels could be found, and plasma acetylcholine does not vary in women with gonadal hormone levels (47). Also, it is not clear if 123I-5-IA is measuring only cell-surface receptors or also internalized receptors; thus, these findings do not rule out sex differences in receptor internalization or synthesis. Finally, other nAChR subtypes may underlie behavioral sex differences. For example, a study found sex differences in β3-nAChR expression in the nasal mucosa of human subjects (48). Future studies are needed to evaluate these alternatives.
The Effect of Menstrual Cycle Phase on β2-nAChR Availability
This study also found no difference in β2-nAChR availability between the early follicular and mid-luteal phases of the menstrual cycle in healthy women nonsmokers. Additionally, in correlational analyses, we observed no relationship between β2-nAChR availability and plasma estrogen or progesterone levels or in their ratio. These results suggest that sex-steroid hormones do not acutely regulate β2-nAChRs. Alternatively, because we imaged during the early follicular phase, when estrogen and progesterone levels are low, and the mid-luteal phase, when both estrogen and progesterone levels are high, we cannot rule out the possibility that estrogen and progesterone have opposing regulatory effects on β2-nAChR availability. Preclinical data suggest an inhibitory role of progesterone and a possible potentiating effect of estrogen on nAChR; thus, during the mid-luteal phase, the high levels of progesterone may be directly inhibiting nAChR or negating the potentiating effects of estrogen on nAChR. β2-nAChR availability may appear similar between the early follicular and mid-luteal phases but may be increased in the absence of progesterone, when only estrogen levels are high—for example, the late follicular phase. One limitation of having studied women during the early to mid-follicular and mid-luteal phases of the menstrual cycle is that the relative effects of estradiol and progesterone on β2-nAChR could not be discerned. The influence of menstrual phase and sex-steroid hormones on other neurotransmitter and receptor systems has been underresearched. Striatal dopamine transporters do not vary with menstrual cycle phase (28), and there is conflicting evidence on variations in dopamine D2 receptors by phase of cycle (49,50). Serotonin transporter availability also does not fluctuate (28), whereas the serotonin 2A receptor (51) may fluctuate with menstrual cycle phase. Finally, γ-aminobutyric acid levels were tightly regulated by the female menstrual cycle (52). Generally, sex steroids may not have acute effects on receptor numbers in the brain, but effects may be more likely to occur in utero, during other developmental periods such as adolescence, or over time because of chronic exposure to sex-steroid hormones.
CONCLUSION
Few in vivo PET or SPECT neuroreceptor imaging studies have examined sex differences or changes in neurotransmitter levels or receptors across the menstrual cycle in living humans, and there are virtually no data on the effects of menstrual phase on human brain derived from postmortem studies. Studies examining the effect of sex and menstrual phase on the brain are important because many psychiatric disorders are marked by sex differences in signs and symptoms that may be a consequence of sex differences in brain chemistry. Here, we have demonstrated no significant difference in brain β2-nAChR availability between men and women nonsmokers or across the menstrual cycle. Importantly, these findings demonstrate sex differences in radiotracer metabolism and plasma protein binding and highlight the critical need to measure plasma radiotracer levels and f1 in studies that include both sexes.
Acknowledgments
This work was supported by grants from NIH (RO1DA015577, P50AA15632, KO1DA020651, KO2DA021863, and NS044316) and by the Ethel F. Donaghue Women's Health Investigator Program of Women's Health Research at Yale. We thank Louis Amici, Nina Sheung, Meredith Terlecki, Tracy Kloczynski, Stephanie Stiklus, and Noah McGuire-Schwartz for technical assistance.
Footnotes
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- Received for publication March 30, 2007.
- Accepted for publication July 2, 2007.





{kind=link}
{kind=link}
{kind=link}
{kind=link}