


Abstract
Adenosine appears to play an important role in tumor growth and metastasis. Synthesized 11C-adenosine 5′-monophosphate (AMP) has recently been reported as a potential tumor-imaging radiotracer. Methods: A variety of human tumor cell lines (SKOV-3, SCC-15, U251, U87, Raji, and Daudi) were incubated with 3.7 kBq (0.1 μCi) of [2-3H]AMP (3H-AMP), [5,6-3H]FDG (3H-FDG), or [2-3H]adenosine (3H-adenosine) in low-physiologic-glucose serum-free medium. Selected cells were exposed to caffeine, dipyridamole, adenosine 5′-(α,β-methylene)diphosphate (APCP), or unlabeled adenosine before exposure to the radiotracer. R-phycoerythrin–conjugated mouse antihuman monoclonal antibody to human CD73 was used for immunophenotyping. High-performance liquid chromatography was used to characterize the intracellular metabolites of 3H-AMP after intracellular uptake. Results: Intracellular uptake of 3H-AMP was significant—10 to 100 times the uptake of 3H-FDG, depending on the particular tumor cell line. Preexposure of SKOV-3 cells to caffeine, a competitive inhibitor of adenosine receptors, did not affect cellular uptake of 3H-AMP. However, preexposure of SKOV-3 cells to dipyridamole, an equilibrative nucleoside transporter inhibitor; APCP, a CD73 (ecto-5′-nucleotidase) inhibitor; or cold adenosine significantly inhibited cellular uptake of 3H-AMP. SKOV-3 uptake of 3H-adenosine was inhibited by dipyridamole but not APCP. U251 uptake of 3H-AMP was significantly inhibited by dipyridamole and APCP. U87 uptake of 3H-AMP was only partially inhibited by dipyridamole and APCP. However, Raji and Daudi cells had significantly lower uptake of 3H-AMP than of 3H-FDG but had significantly increased uptake of 3H-adenosine, which was inhibited by dipyridamole. Raji and Daudi cells were negative, but the SKOV-3 cells positive, for CD73 cell-surface expression. 3H-Adenosine metabolites were persistently retained after influx into the cell, predominantly as 3H-adenosine triphosphate and 3H-adenosine diphosphate. Conclusion: Cancer cell lines evaluated in vitro had significantly elevated uptake of radiolabeled AMP, on the order of 10- to100-fold, in comparison to radiolabeled FDG. The mechanism of intracellular uptake depends predominantly on equilibrative nucleoside transporters after conversion of AMP to adenosine by CD73 in SKOV-3, SCC-15, and U251 cells. Intracellular retention is due to phosphorylation to adenosine triphosphate and adenosine diphosphate. Raji and Daudi cells have low uptake of radiolabeled AMP because of a lack of CD73 expression. This in vitro evaluation using 3H-AMP with tumor cell lines supports the potential of 11C-AMP for use in targeting the nucleoside transport pathway in PET imaging of tumors.
Nucleoside and nucleoside analogs are a pharmacologically diverse family of molecules that have been synthesized and used for cytotoxic, antiviral, and immunosuppressive therapies (1,2). Adenosine, a purine nucleoside, is increasingly being found to play an important role in tumor growth and metastasis (3). Adenosine is reported to be present at high concentrations in solid tumors, with accumulation in the intracellular and extracellular tumor microenvironments, at sites of local tissue injury, and under conditions of hypoxia, and is reported to stimulate tumor growth and angiogenesis and to have immunosuppressive effects (3,4).
Local areas of hypoxia and high metabolic rates of proliferating tumor cells, with increased rates of glucose use and lactate release, characterize the microenvironment of solid tumors. These aberrant conditions can lead to increased levels of extracellular or intracellular adenosine. Increased extracellular adenosine levels can be a consequence of increased adenosine production and release due to cell death and nucleotide degradation, ischemia and adenosine triphosphate (ATP) breakdown, ATP/adenosine diphosphate (ADP) release and subsequent dephosphorylation, adenosine 5′-monophosphate (AMP) release, and S-adenosylhomocysteine hydrolysis (3,5).
Under specific physiologic or pathologic conditions, intracellular adenosine accumulation is thought to coordinate changes in the patterns of expression of adenosine-metabolizing enzymes to maintain low or high intracellular adenosine concentrations. The differentiation of the promyelocytic cell line HL-60 along the macrophage lineage by a differentiating agent, phorbol 12-myristate 13-acetate, changed the pattern of expression of 3 adenosine-metabolizing enzymes and caused a 2.5-fold increase in intracellular adenosine uptake and retention in phorbol 12-myristate 13-acetate–stimulated cells (3,6). Spychala et al. (3,7) theorized that most malignant cells of epithelial origin, macrophages, and ischemic myocardium may have an adenosine-accumulating phenotype whereas thymocytes and lymphoid cells may have adenosine-removing phenotypes and decreased intracellular adenosine accumulation. In another study (8), C6 cells were shown to take up and accumulate adenosine under ATP-depleting conditions. This was shown to result from ATP metabolism via an inosine 5′-monophosphate pathway, which does not include adenosine, resulting in formation and release of hypoxanthine rather than adenosine.
Nucleoside transporters play a key role in the physiologic control of adenosine by regulating the extracellular adenosine available to cell-surface receptors and the intracellular concentration of adenosine and adenylates. Nucleoside transport is an important determinant of the pharmacokinetics, plasma and tissue concentrations, disposition, and in vivo biologic activity of adenosine and of nucleoside analogs used for antiviral and anticancer therapies (2,9).
The 2 major classes of nucleoside transporters consist of the equilibrative nucleoside transporters (ENTs) and the concentrative nucleoside transporters (CNTs). In humans, the ENTs mediate the transport of nucleosides across the cell membrane by a concentration gradient. Human ENTs (hENTs) exhibit broad substrate specificity for pyrimidine and purine nucleosides and are subdivided on the basis of their sensitivity to inhibition by 6-[(4-nitrobenzyl)thio]-9-β-d-ribofuranosylpurine (NBMPR) (2,10). hENT1 is sensitive to NBMPR inhibition at low nanomolar concentrations and has broad tissue distribution. hENT2 is resistant to NBMPR inhibition at micromolar concentrations and is expressed predominantly in the skeletal muscle and brain. Dipyridamole inhibits both hENT subtypes, although hENT1s are more susceptible than hENT2s (2,11). CNTs use the electrochemical gradient of either a sodium or hydrogen ion to drive the transport of nucleosides against a concentration gradient. They have a more limited tissue distribution, are typically insensitive to NBMPR inhibition, and are subdivided on the basis of their substrate selectivity.
Knowledge of the relative abundance of different transporters, and of their differing abilities to transport nucleoside analogs, may provide clues to the origin of the selectivity and efficacy of individual nucleoside drugs. Nucleoside transporters may play a role in resistance to nucleoside chemotherapeutic agents (12–14). Absence of hENT1 was found to be associated with reduced survival in patients with pancreatic cancer treated with gemcitabine (15). Reduction or loss of hCNT1 was found to correlate with a poor prognosis for various histologic types of gynecologic tumors (16). hENT2 protein expression was found to correlate with in vitro sensitivity to fludarabine in chronic lymphocytic leukemia cells (17), and ENT expression was found to correlate with the in vitro sensitivity of acute leukemia cells (18).
Prior attempts to study adenosine metabolism and biodistribution with phosphorus- or technetium-labeled adenylates have been hampered by rapid hydrolysis with loss of the radioactive label. 18F-labeled adenosine analogs have been reported, but metabolic defluorination was a problem and high bone uptake of 18F-fluoride was seen (19,20). The synthesis and biodistribution of 11C-AMP have recently been published; AMP was intrinsically labeled at the C-8 position with 11C (21). Ex vivo biodistribution in normal mice showed the highest uptake of 11C-AMP in the lungs, heart, and blood, and clearance was predominantly through the urinary system. As an important mediator and reporter of cellular damage, hypoxia, and inflammation, a positron-emitting radiolabeled adenylate may be a useful radiotracer for studying adenosine metabolism in normal physiologic and altered pathologic conditions. Here, we report an in vitro study of the intracellular accumulation of [2-3H]AMP, ammonium salt (3H-AMP), in various human tumor cell lines to evaluate the potential of 11C-AMP for use as a positron-emitting radiotracer for oncologic PET.
MATERIALS AND METHODS
Radiochemicals
3H-AMP and [2-3H]adenosine, ammonium salt (3H-adenosine), were purchased from Amersham Biosciences. 3H-AMP was provided in an ethanol:water (1:1) solution at 37 MBq/mL (1 mCi/mL) and a specific activity of 0.37–1.1 TBq/mmol (10–30 Ci/mmol). 3H-Adenosine was provided in a sterile aqueous solution containing 0.1% ethanol at 37 MBq/mL (1 mCi/mL) and a specific activity of 740–925 GBq/mmol (20–25 Ci/mmol). [5,6-3H]FDG (3H-FDG) was purchased from American Radiolabeled Chemicals, Inc., and was provided in a sterile saline solution at 37 MBq/mL (1 mCi/mL) and a specific activity of 0.74–2.22 TBq/mmol (20–60 Ci/mmol).
Cell Lines and Media
SKOV-3 (ovarian adenocarcinoma), Raji (B-cell lymphoma), Daudi (B-cell lymphoma), SCC-15 (tongue squamous cell carcinoma), and U87 (anaplastic astrocytoma) cell lines were purchased from American Type Culture Collection and cultured in medium under conditions specified by the supplier. The U251 cells (human glioblastoma multiforme) (gift of the laboratory of Howard A. Fine at the National Cancer Institute and National Institute of Neurological Disorders and Stroke, National Institutes of Health) were maintained in RPMI 1640 medium containing 5% fetal bovine serum. Medium was purchased from American Type Culture Collection or Gibco BRL.
Chemicals
Dipyridamole was purchased from Bedford Laboratories at 10 mg/2 mL and stored at room temperature. Caffeine citrate (CAFCIT) was purchased from Boehringer Ingelheim Pharmaceuticals, Inc., at 60 mL/3 mL and stored at room temperature. Adenosine 5′-(α,β-methylene)diphosphate (APCP), adenosine, N-methyl-D-glucamine (NMG), and NBMPR were purchased from Sigma-Aldrich. APCP was reconstituted with sterile water to a stock solution of 50 mg/mL and stored at −20°C. Adenosine was reconstituted with sterile water to a 1 mol/L stock solution and stored at 4°C.
In Vitro Uptake
Cell lines at log phase were incubated for 1 h with 0.5 mL of RPMI 1640 plus glucose (0.8 g/L) in the individual wells of a 24-well plate. Low-physiologic-glucose medium was used for the in vitro experiments to simulate a fasting human serum glucose level, as is standard for FDG imaging. For conditions requiring dipyridamole, caffeine, APCP, or adenosine, varying concentrations of these reagents were added to the RPMI 1640 plus glucose for the 1-h initial incubation. After the initial incubation, 3.7 kBq (0.1 μCi) of radiotracer (3H-AMP, 3H-FDG, or 3H-adenosine) at 3.7 kBq/10 μL (0.1 μCi/10 μL) were added to each well and reincubated for a fixed time (15, 45, or 75 min; 75 min for fixed-time-point experiments).
Adherent cells and suspension cells were harvested differently. Adherent cells were washed carefully twice with 0.9 mL of ice-cold phosphate-buffered saline (PBS), followed by the addition of 0.5 mL of trypsin (0.25%) and ethylenediaminetetraacetic acid (1 mmol/L) (Gibco/Invitrogen) and incubation for 15 min at 37°C to detach cells. The cells were collected in a scintillation vial, the well was washed with PBS, and the wash was collected in a scintillation vial to recover additional cells. Suspension cells were transferred to a microcentrifuge tube (Microfuge; Beckman Coulter, Inc.), and the well was washed with ice-cold PBS and collected in a scintillation vial to recover additional cells. The cells were centrifuged at ∼500 relative centrifugal force in a microcentrifuge, the medium was removed, and the cells were resuspended in ice-cold PBS. The cells were recentrifuged and washed with ice-cold PBS one more time. The cells were then lysed with 0.5 mL of NaOH (0.3 mol/L) plus sodium dodecylsulfate (0.1%) and incubated at room temperature for more than 1 h. The lysates were transferred to a scintillation vial, the microcentrifuge tube was washed with 0.9 mL of PBS, and the wash was added to the scintillation vial. All experiments were performed in triplicate (3 wells per experimental condition).
Cell uptake retention was studied with an initial uptake period of 1 h. Afterward, medium containing 3H-AMP was removed, and the cells were washed twice with RPMI 1640 plus glucose medium (0.8 g/L), followed by incubation with the same glucose medium for 30 min, 1 h, or 2 h. The cells were then washed and harvested as already described.
Fluorescence Immunophenotyping
Flow cytometry of SKOV-3, Raji, and Daudi cells was performed to determine their cell-surface expression of CD73 (ecto-5′-nucleotidase). R-phycoerythrin–conjugated mouse antihuman monoclonal antibody to CD73 was purchased from BD PharMingen. In 100 μL of PBS plus 2% fetal bovine serum, 2 × 106 cells were resuspended, incubated with 10 μL of anti-CD73 antibody/106 cells for 15 min at 4°C, centrifuged for 8 min at 1,500 rpm, and resuspended in 400 μL of RPMI. Flow cytometry was performed on the FACSCalibur machine (Becton-Dickinson).
Nucleoside Uptake Assays
SKOV-3 and U87 cells were washed twice in physiologic buffer (NaCl [118 mmol/L], N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid [25 mmol/L], KCl [4.9 mmol/L], K2HPO4 [1.4 mmol/L], MgCl2 [1.2 mmol/L], CaCl2 [1 mmol/L], and glucose [11 mmol/L] brought to pH 7.4 with NaOH) or buffer in which NaCl had been replaced with NMG. The cells were incubated with 3.7 kBq (0.1 μCi) of 3H-adenosine at 3.7 kBq/10 μL (0.1 μCi/10 μL) in sodium-free NMG physiologic buffer with varying concentrations of NBMPR (0.1–10,000 nmol/L) for 15 min and were harvested as described in the cell uptake methods. The cells were also incubated with 3.7 kBq (0.1 μCi) of 3H-adenosine at 3.7 kBq/10 μL (0.1 μCi/10 μL) in either sodium-free NMG physiologic buffer or NaCl containing physiologic buffer for 5, 10, or 20 min and harvested as described in the cell uptake methods.
High-Performance Liquid Chromatography
Acid extracts of cells exposed to 3H-AMP were chromatographed using a reversed-phase gradient elution method described by Glenn et al. (22). Briefly, 0.5N perchloric acid extracts neutralized with potassium hydroxide were chromatographed on a Synergi Hydro-RP column (4.6 × 250 mm; Phenomenex) with potassium phosphate buffer (0.1 mol/L; pH 6.0) at a flow rate of 0.7 mL/min. After elution for 9 min, a 4-min gradient to 15% methanol in buffer (0.1 mol/L) was started, and this solvent was continued for an additional 22 min before returning to buffer (0.1 mol/L) for 1 min. During the chromatography, the eluate was collected as eighty 0.45-min fractions. Radioactivity was determined by scintillation counting. Labeled nucleotides and inosine were identified by cochromatography with standards.
Statistical Analysis
The Student t test was applied for all statistical analyses, using a P value of less than 0.05 to evaluate for statistical significance.
RESULTS
Intratumoral Uptake of 3H-AMP
SKOV-3 and SCC-15 tumor cell lines were exposed to 3.7 kBq (0.1 μCi) of 3H-AMP and 3H-FDG in vitro for 75 min at 37°C after preincubation in a serum-free medium designed to simulate human physiologic fasting levels of serum glucose (0.8 g/L). The uptake of 3H-AMP by SKOV-3 and SCC-15 cells was much higher than the uptake of 3H-FDG. In SKOV-3 cells, intracellular uptake was 1.43 × 106 disintegrations per minute (DPM)/106 cells for 3H-AMP, compared with 2.37 × 104 DPM/106 cells for 3H-FDG (P < 0.05 [P = 9.7 × 10−8]). In SCC-15 cells, intracellular uptake was 8.76 × 105 DPM/106 cells for 3H-AMP, compared with 4.42 × 104 DPM/106 cells for 3H-FDG (P < 0.05 [P = 8.5 × 10−5]). Incubation of SKOV-3 and SCC-15 cells for 15, 45, and 75 min of exposure to 3H-AMP and 3H-FDG indicated a trend toward increasing uptake with increasing incubation time for both radiotracers (data not shown). Other cell lines, including breast adenocarcinoma (MN-1) (data not shown), U87, and U251, tested at 75 min of exposure also showed significantly greater uptake of 3H-AMP than of 3H-FDG.
Nucleoside Transport–Mediated Uptake of 3H-AMP
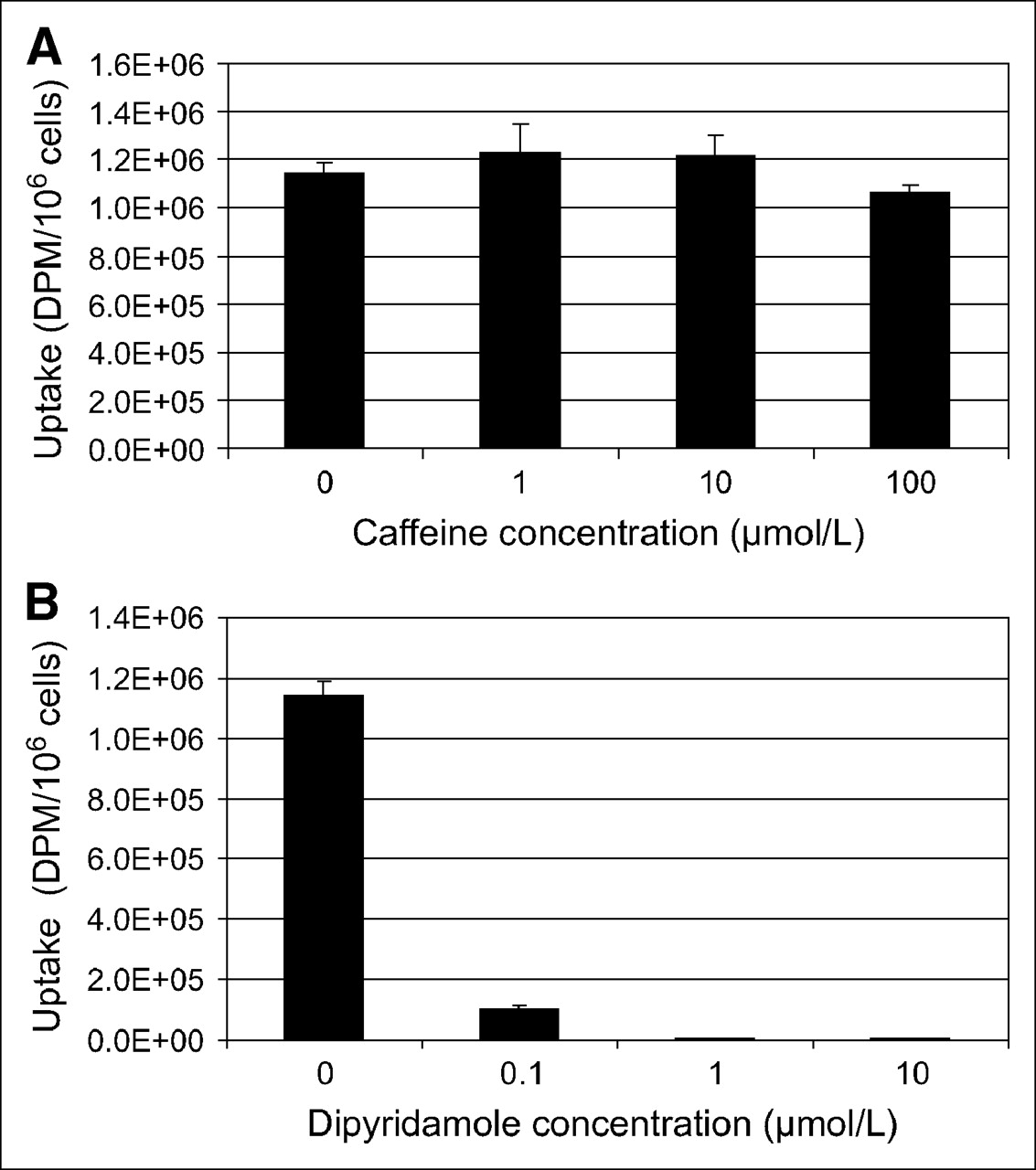
To assess whether 3H-AMP accumulation in tumor cells was mediated by a nucleoside transporter or by binding to an extracellular adenosine receptor, we pretreated SKOV-3 cells with varying concentrations of caffeine, a competitive inhibitor of the adenosine receptors (A1, A2, and A3), and dipyridamole, an inhibitor of hENT (Fig. 1). Dipyridamole at nanomolar to micromolar concentrations significantly inhibited 3H-AMP uptake. Micromolar concentrations of caffeine did not significantly inhibit 3H-AMP uptake. SCC-15 uptake of 3H-AMP was also significantly inhibited by dipyridamole but not by caffeine (data not shown). These findings indicate that 3H-AMP is accumulated in SKOV-3 and SCC-15 cells via ENT rather than by an adenosine receptor–binding mechanism.
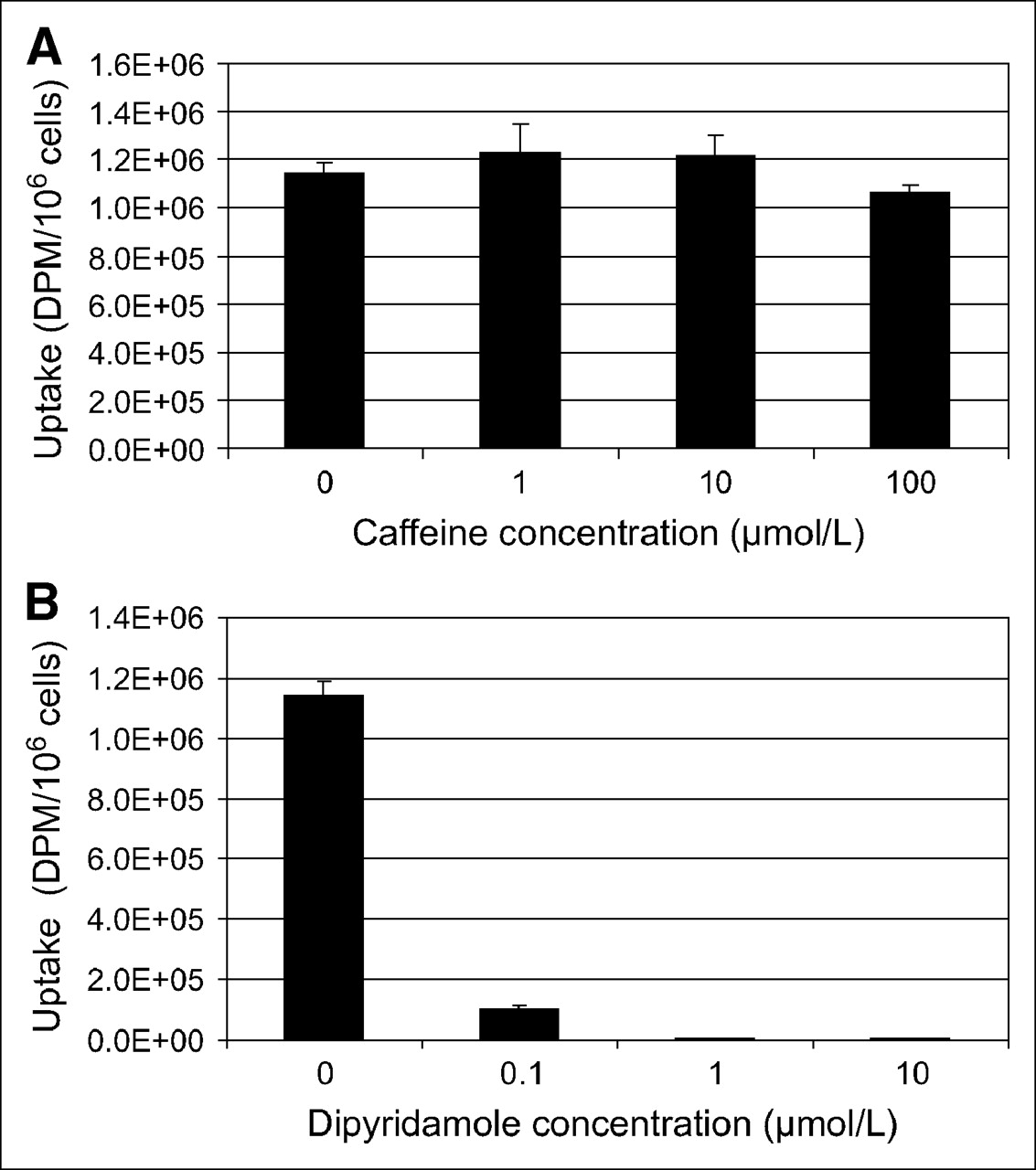
3H-AMP is accumulated in SKOV-3 cells via nucleoside transporter and not by adenosine receptor binding. Uptake in SKOV-3 cells after exposure to 3H-AMP is not inhibited by caffeine (A), an adenosine receptor antagonist, but is inhibited by dipyridamole (B), an ENT inhibitor, in dose-dependent manner.
Need for 3H-AMP Metabolism to 3H-Adenosine Before Cellular Uptake
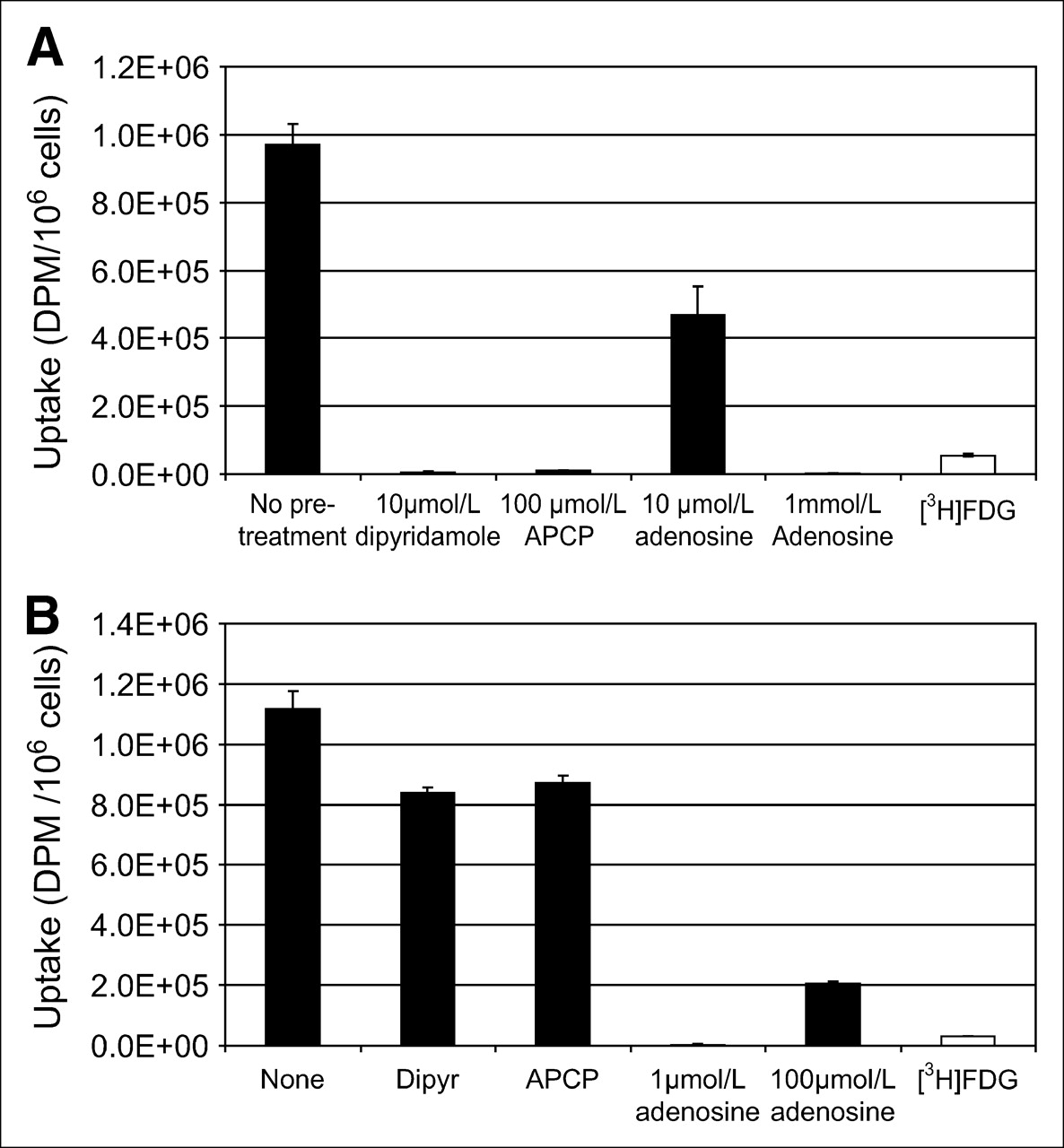
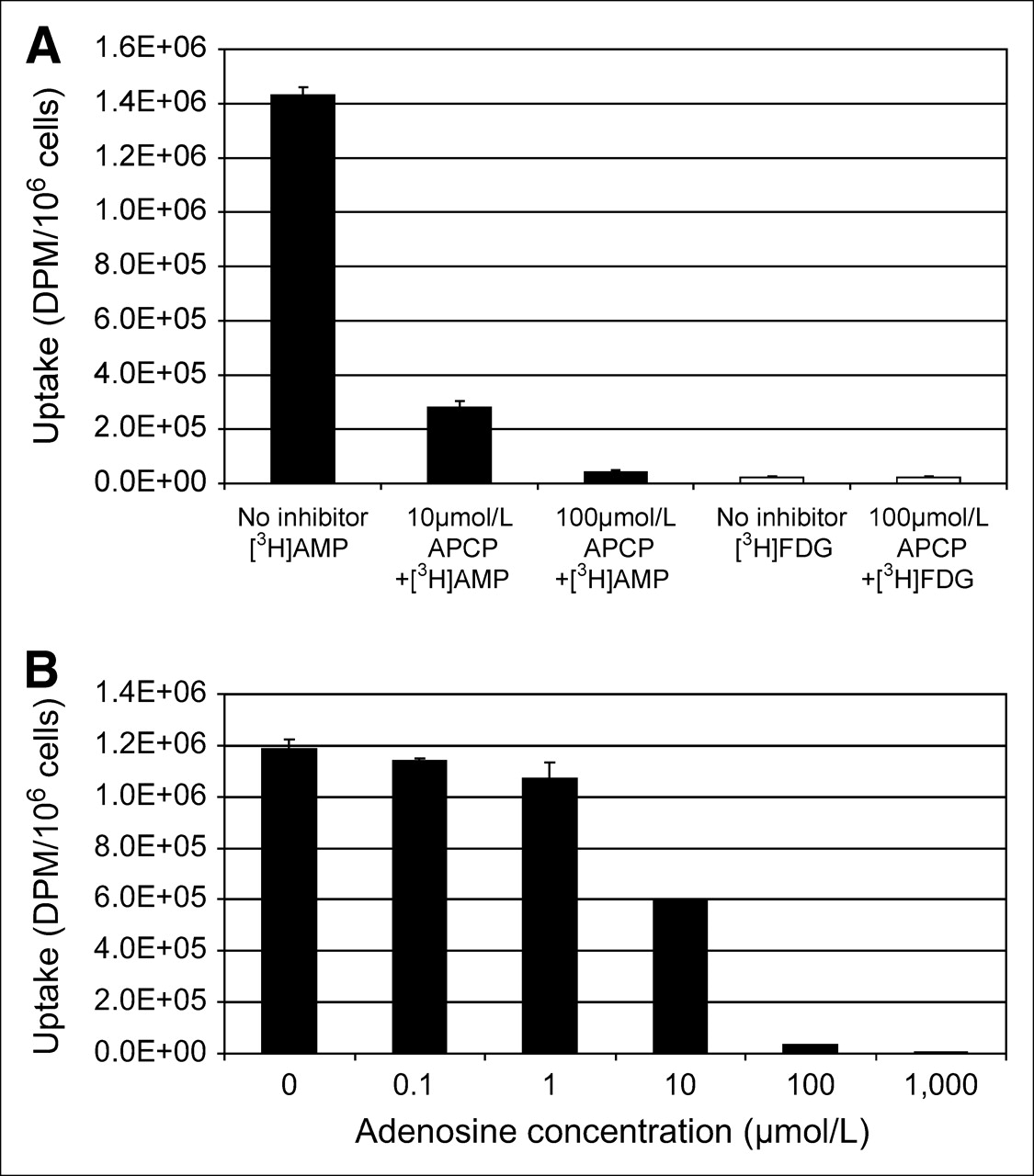
α,β-Methylene adenosine-5′-diphosphate (APCP) is an inhibitor of CD73, an ecto-5′-nucleotidase that dephosphorylates adenosine monophosphate to adenosine nucleoside. Pretreatment of SKOV-3 cells with APCP at micromolar concentrations decreased uptake of 3H-AMP significantly and in a dose-dependent manner (Fig. 2A). Combination of dipyridamole with APCP further inhibited 3H-AMP uptake. Pretreatment of SKOV-3 cells with APCP did not significantly affect 3H-FDG uptake. Pretreatment of SCC-15 cells with APCP significantly inhibited uptake of 3H-AMP but did not affect uptake of 3H-FDG (data not shown). These findings indicate that dephosphorylation of 3H-AMP via CD73 is necessary for tracer accumulation in SKOV-3 and SCC-15 tumor cells.

3H-AMP is converted to 3H-adenosine before tumor cell uptake via nucleoside transporter. (A) Inhibition of CD73 (ecto-5′-nucleotidase) with APCP (α,β-methylene adenosine-5′-diphosphate) blocks uptake of 3H-AMP (black bars), but not same dose of 3H-FDG (white bars), in dose-dependent manner. (B) 3H-AMP uptake in SKOV-3 cells is competitively inhibited by exposure to increasing concentrations of extracellular unlabeled adenosine.
Increasing concentrations of cold adenosine inhibited 3H-AMP uptake by SKOV-3 cells in a dose-dependent manner (Fig. 2B). This effect was strongest at higher millimolar levels, indicating that adenosine competitively inhibits uptake of 3H-AMP after dephosphorylation to 3H-adenosine. Pretreatment of SKOV-3 cells with APCP did not inhibit uptake of 3H-adenosine but significantly inhibited uptake of 3H-AMP. However, pretreatment of SKOV-3 cells with dipyridamole significantly inhibited uptake of both 3H-adenosine and 3H-AMP (data not shown). Uptake of 3H-adenosine was competitively inhibited by increasing micromolar concentrations of unlabeled adenosine (data not shown). These data provide further evidence that 3H-AMP is metabolized near the cell surface by CD73 to 3H-adenosine before intracellular uptake via the ENT.
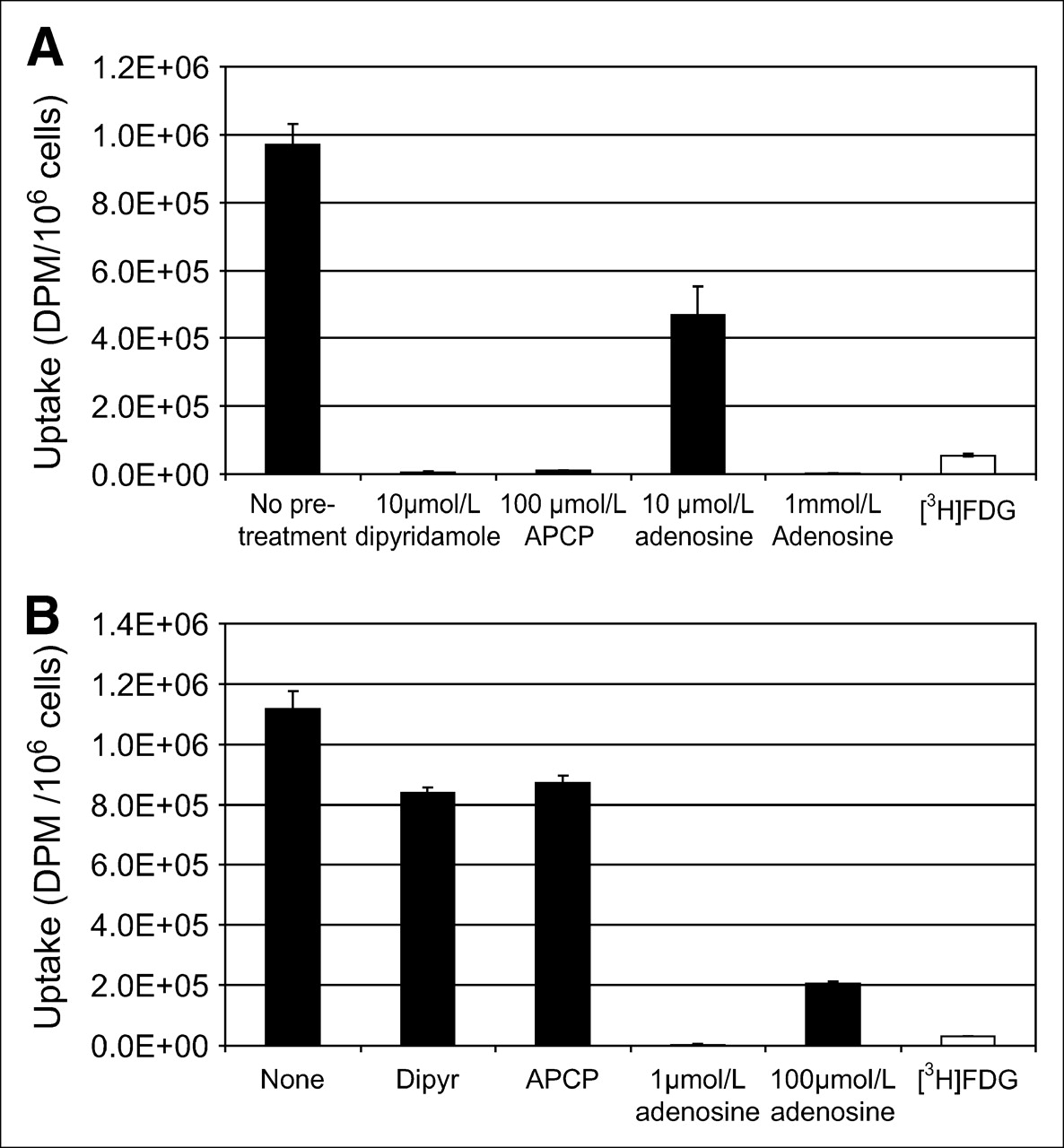
U251 and U87 Glioma Cell Uptake of 3H-AMP
Human glioma cell lines U251 and U87 were evaluated for uptake of 3H-AMP (Fig. 3). U251 cells take up 3H-AMP significantly more than they do 3H-FDG (P < 1.4 × 10−5). 3H-AMP uptake is significantly inhibited by dipyridamole, APCP, and increasing concentrations of cold adenosine (Fig. 3A). This finding provides evidence that the mechanism of uptake of 3H-AMP in U251 cells involves dephosphorylation by CD73 and uptake of 3H-adenosine by hENT. U87 cells also take up 3H-AMP significantly more than they do 3H-FDG (P < 6.2 × 10−6) (Fig. 3B). Intracellular uptake of 3H-AMP in U87 cells is significantly inhibited by increasing concentrations of adenosine; however, pretreatment with dipyridamole and APCP only partially inhibits 3H-AMP uptake in U87 cells. This finding suggests that, before uptake, 3H-AMP is converted to 3H-adenosine predominantly by a mechanism other than via CD73 and ENT, probably through other phosphorylases or nucleoside transporters.
(A) Significantly more 3H-AMP than 3H-FDG accumulates in U251 human glioblastoma cells (P < 0.05). 3H-AMP is dephosphorylated by CD73, and 3H-adenosine is taken up by ENT nucleoside transporter in U251 cells. Cells were exposed to 3.7 kBq (0.1 μCi) of radiotracer in vitro. (B) Significantly more 3H-AMP than 3H-FDG accumulates in U87 human anaplastic astrocytoma cells (P < 0.05). Dephosphorylation to 3H-adenosine is required, but transport is predominantly via non–dipyridamole-inhibited ENT route. Cells were exposed to 3.7 kBq of radiotracer in vitro. Dipyr = dipyridamole.
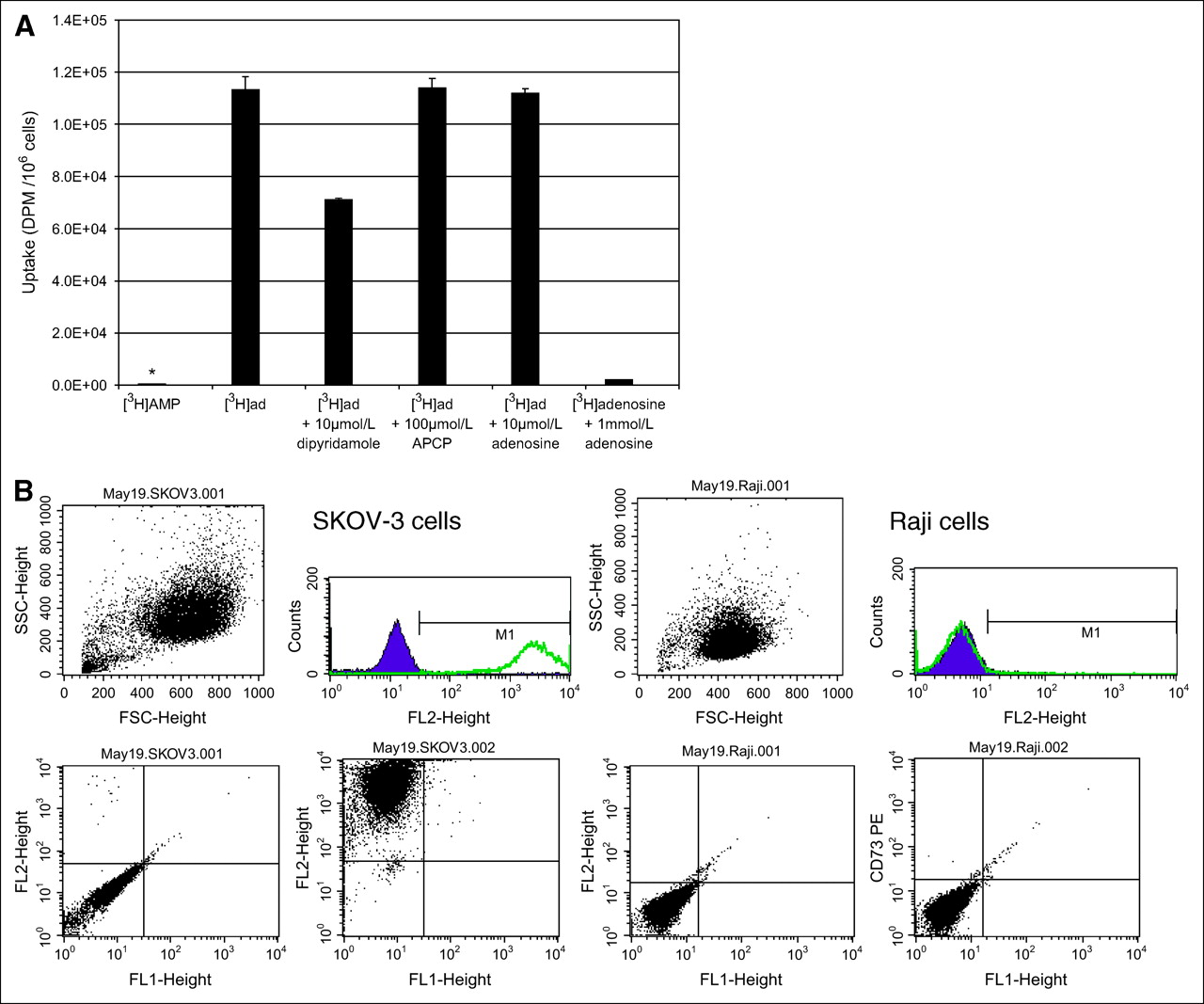
Lack of Significant 3H-AMP Uptake Because of Lack of CD73 Expression in Raji and Daudi Cells
The literature contains evidence that cells of adenosine-generating or adenosine-removing phenotypes, and most malignant cells of epithelial origin, may fall into the adenosine-removing phenotype with increased uptake and accumulation of extracellular adenosine (3). To determine whether nonepithelial malignant cells would take up adenosine, we evaluated the B-cell lymphoma cell lines, Raji and Daudi cells, for their uptake of 3H-AMP. Raji cells had very low 3H-AMP uptake that was not significantly inhibited by dipyridamole and APCP (Fig. 4A). However, uptake of 3H-adenosine in Raji cells was significantly increased and was uninhibited by APCP, partially inhibited by dipyridamole, and inhibited by increasing concentrations of nonradiolabeled adenosine. This finding suggests that Raji cells accumulate 3H-adenosine by ENT and other nucleoside transporters but lack the ability to dephosphorylate 3H-AMP to 3H-adenosine for subsequent uptake by these transporters. We obtained similar results for Daudi cells (data not shown).
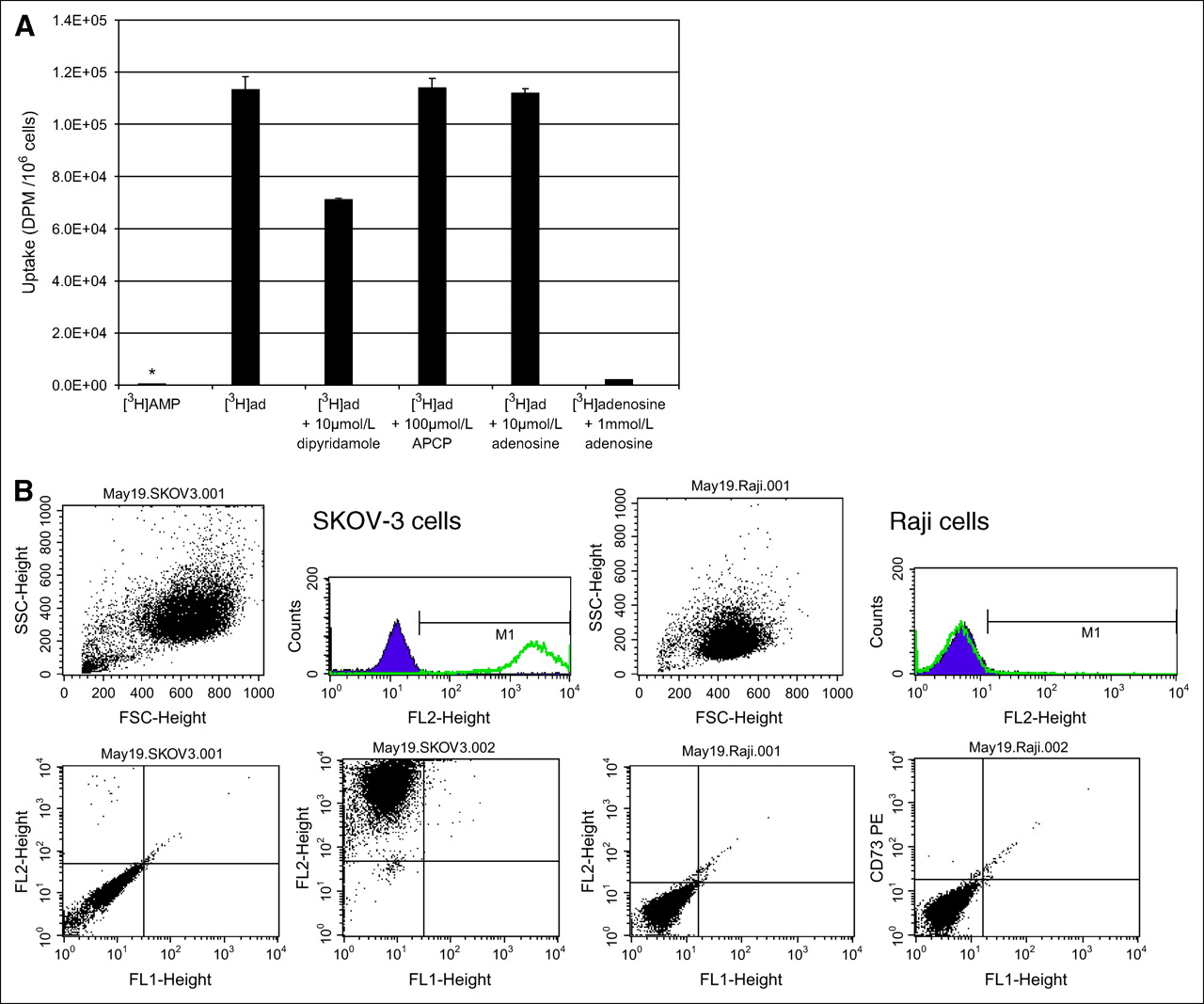
Low uptake of 3H-AMP by Raji cells because of lack of CD73 expression. (A) In Raji cells, uptake of 3H-AMP (*) is very low and uptake of 3H-adenosine is high (P < 0.05). 3H-Adenosine uptake is only partially inhibited by dipyridamole and is competitively inhibited by unlabeled adenosine. CD73 inhibition by APCP does not affect 3H-adenosine uptake. Cells were exposed to 3.7 kBq of radiotracer in vitro. (Daudi cells also showed similarly low uptake of 3H-AMP and high uptake of 3H-adenosine—data not shown.) (B) Extracellular CD73 is expressed on SKOV-3 cells but not on Raji cells. Viable cells were selected by physical parameters, size, and internal complexity (upper left graphs). Immunophenotyping of these viable cells was compared using cells labeled with phycoerythrin-conjugated CD73 antibody and control cells without antibody labeling. Cell fluorescence intensity is graphed on y-axis and cell physical parameter on x-axis for control cells (lower left graphs) and CD73-labeled cells (lower right graphs). These graphs are also represented as number of cells on y-axis versus fluorescence intensity on x-axis (upper right graphs). Consistent with CD73 expression, intensity of fluorescence is greater on SKOV-3 cells labeled with CD73 than on control cells. However, Raji cells did not show difference in fluorescence intensity between cells labeled with CD73 and control cells. (Daudi cells were also negative for CD73—data not shown.)
Flow cytometry of Raji or Daudi cells stained with fluorescent R-phycoerythrin–conjugated monoclonal mouse antihuman CD73 antibody were negative for cell-surface expression of CD73 (Fig. 4B). However, SKOV-3 cells, in which 3H-AMP tracer uptake was inhibited by APCP, were significantly positive for CD73 cell-surface expression. This finding strongly suggests that dephosphorylation of adenosine monophosphate by CD73 expression is an important step in the intracellular uptake of 3H-AMP by tumor cells in vitro.
Nucleoside Transporter Characterization of SKOV-3 and U87 Cells
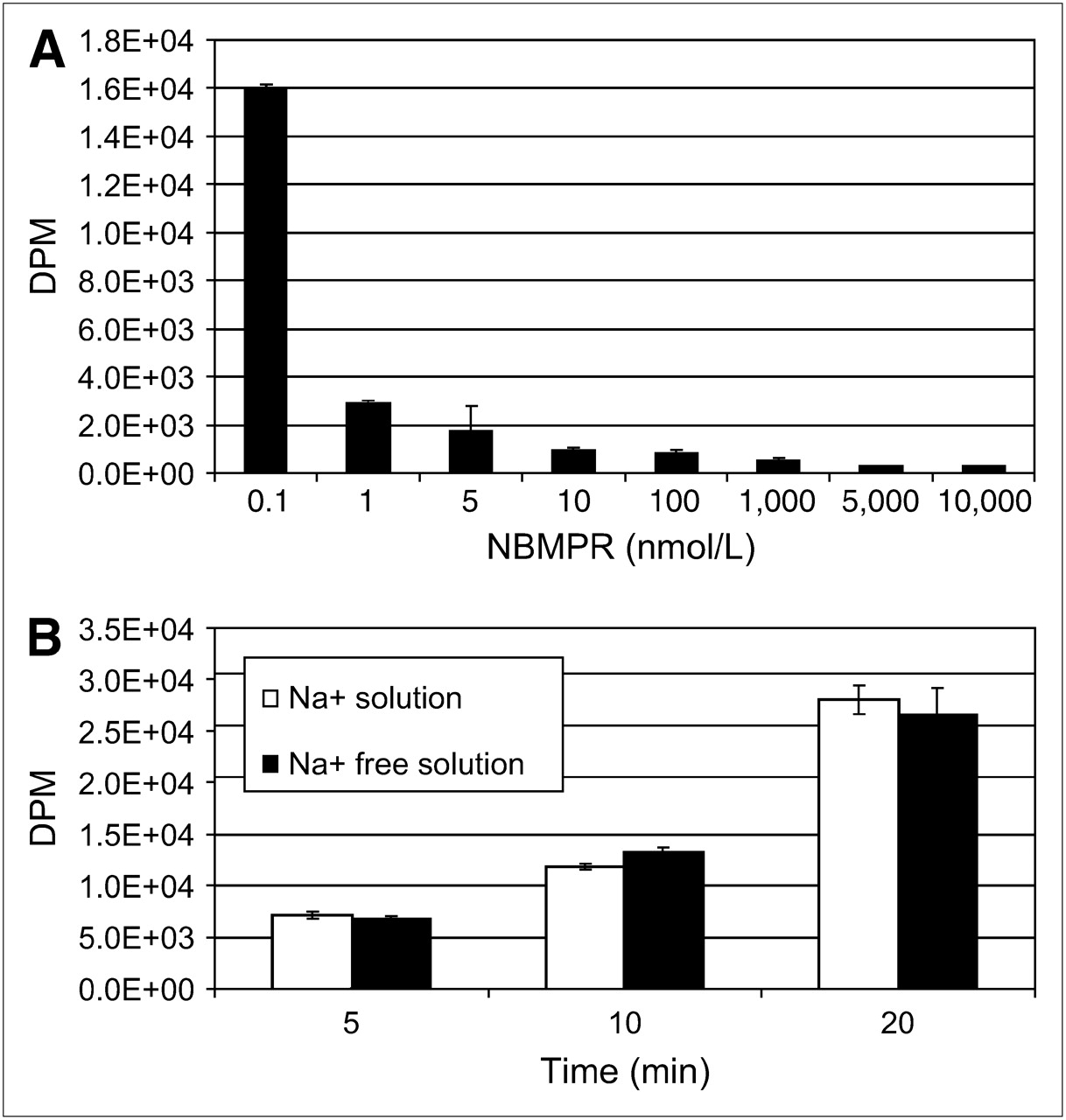
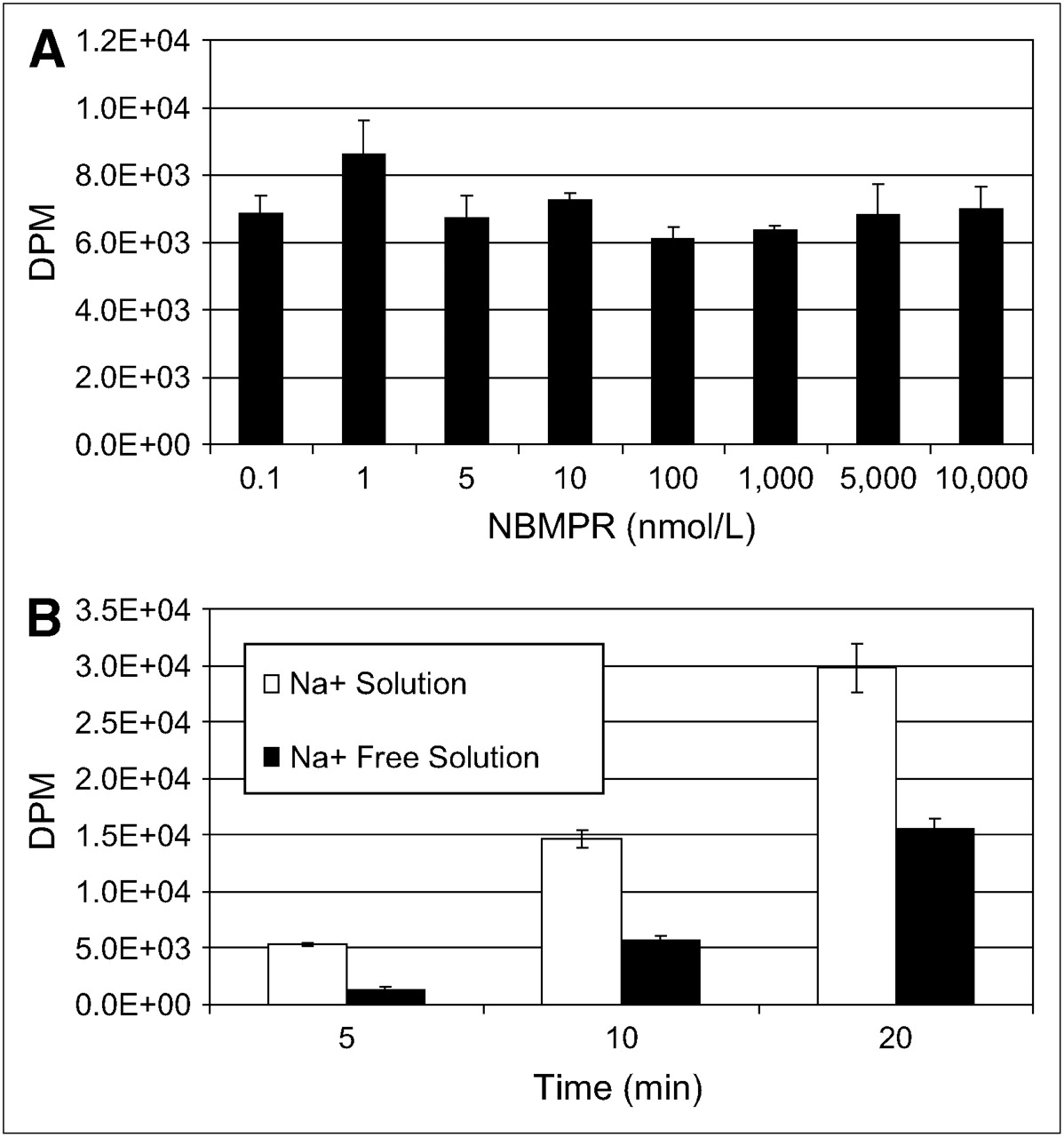
To identify the nucleoside transporter subtypes present in SKOV-3 and U87 cells, we functionally characterized the intracellular uptake of 3H-adenosine using 2 different experiments. First, we evaluated the inhibition of sodium-independent uptake of 3H-adenosine by ENTs with varying concentrations of NBMPR in physiologic buffer that was sodium-free (with NMG replacing NaCl). ENT1 is inhibited by low nanomolar concentrations of NBMPR, and ENT2 is unaffected by less than 1 μmol/L concentrations of NBMPR. Second, we evaluated the uptake of 3H-adenosine in physiologic and sodium-free physiologic buffers to determine whether nucleoside transport was Na+-dependent. In SKOV-3 cells, 3H-adenosine uptake was inhibited by low nanomolar concentrations of NBMPR (Fig. 5A) and there was no evidence of a Na+-dependent nucleoside transport (Fig. 5B), indicating a predominant ENT1 uptake mechanism. In U87 cells, 3H-adenosine uptake was not significantly inhibited by either nanomolar or micromolar concentrations (up to 10 μmol/L) of NBMPR (Fig. 6A), and about half the nucleoside uptake was by Na+-dependent nucleoside transport (Fig. 6B). This finding indicates the presence of both ENTs and CNTs in U87 cells. The subtype of ENT is unclear and may be due to a non-ENT1 or a non-ENT2 or a mutation of ENT1 or ENT2 resulting in resistance to NBMPR.
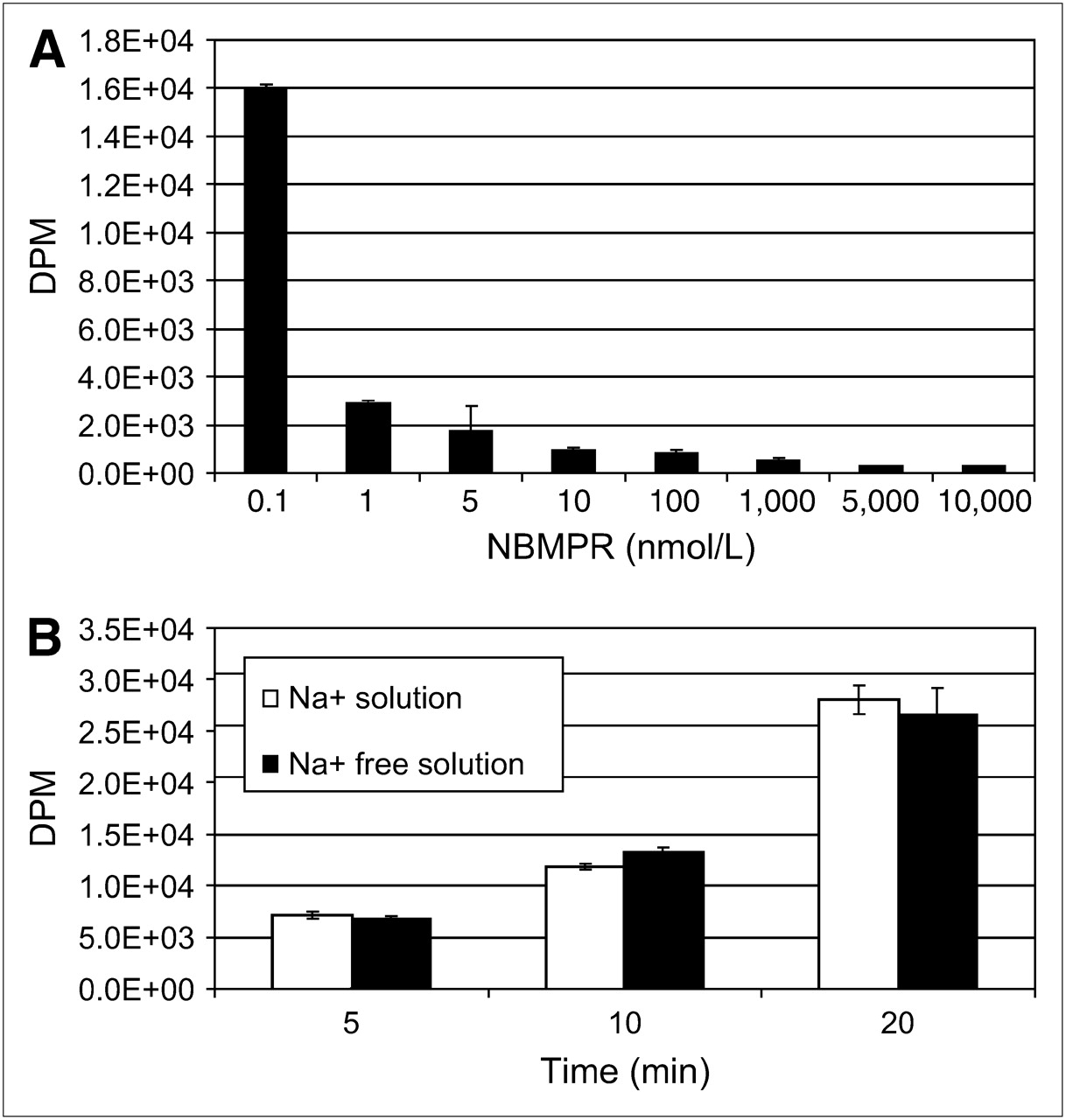
Nucleoside transporter characterization reveals that SKOV-3 cells possess primarily hENT1s. (A) 3H-Adenosine uptake is inhibited by low nanomolar concentrations of NBMPR in Na+-free physiologic solution. (B) No evidence is seen of concentrative Na+-dependent 3H-adenosine transport in SKOV-3 cells in physiologic or Na+-free physiologic buffer.
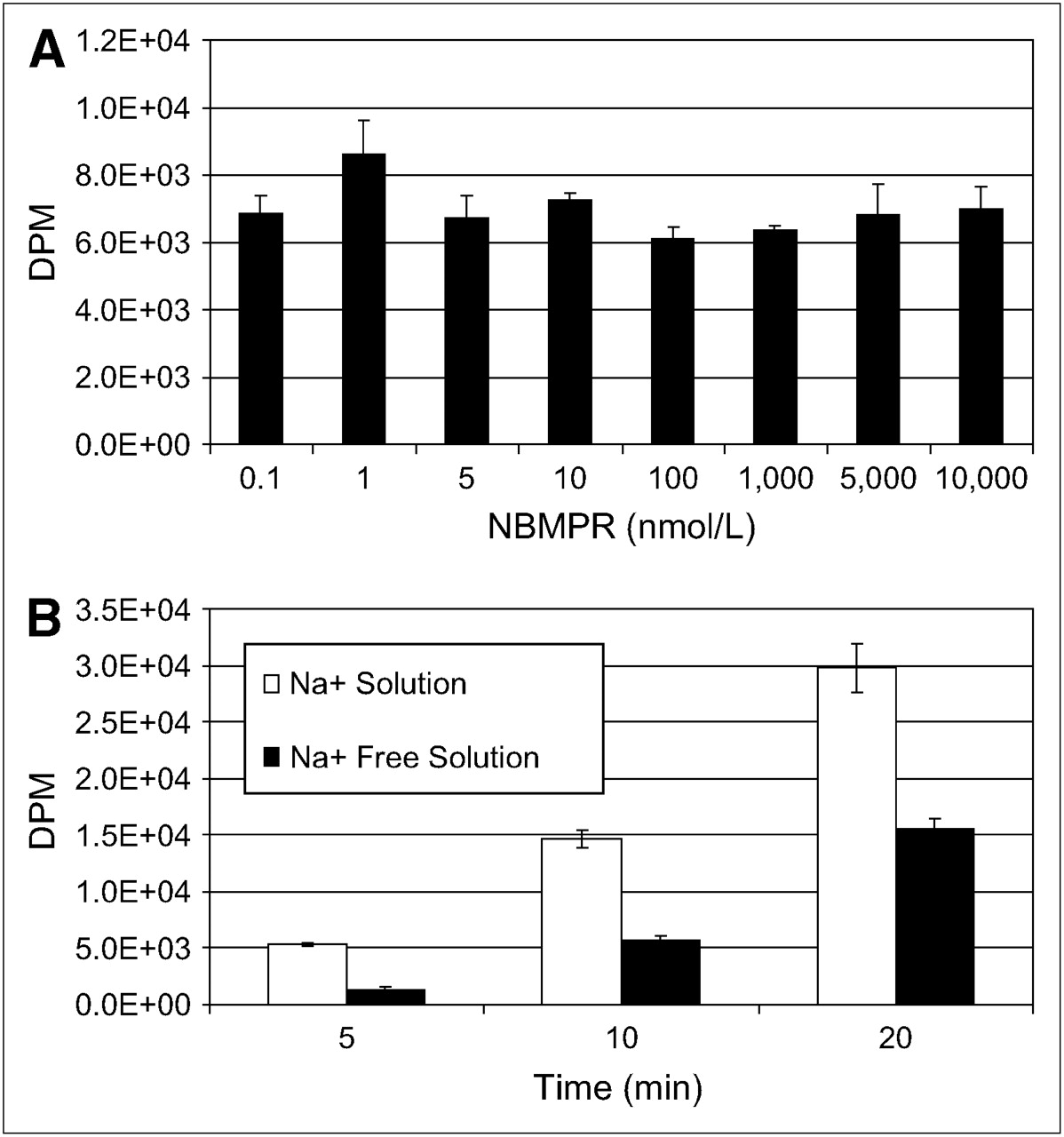
Nucleoside transporter characterization of U87 cells. U87 cells possess both ENTs and CNTs. ENT appears to be NBMPR resistant. (A) 3H-Adenosine uptake is not inhibited by nanomolar or micromolar concentrations of NBMPR in Na+-free physiologic solution. (B) Evidence is seen of concentrative Na+-dependent 3H-adenosine transport in SKOV-3 cells in physiologic or Na+-free physiologic buffer.
Intracellular Retention of 3H-AMP
SKOV-3 cells were exposed to 3H-AMP to determine the 1-h uptake after removal of the radiotracer medium and exposure to a 3H-free medium. The purpose of this was to stop further influx of 3H-AMP and allow for efflux of intracellular 3H-AMP. Intracellular retention of 3H was significant—comparable to the initial uptake even after 30 min to 2 h of “washout” (Table 1). Intracellular acid extracts of cells exposed to 3H-AMP were analyzed by high-performance liquid chromatography, which demonstrated that most of the intracellular 3H pool consisted of ATP and ADP, low levels of inosine, and minimal AMP species (Table 2). 3H-Adenosine was not found in the intracellular 3H pool.
3H-AMP Retention in SKOV3 Cells After 1 Hour of Uptake Followed by Washout with 3H-AMP–Free Medium
High-Performance Liquid Chromatography Determination of Intracellular Adenylate and Inosine Species in SKOV-3 Cells
DISCUSSION
Adenosine is a ubiquitous intra- and extracellular compound involved in various physiologic and pathologic processes. Recent studies have implicated adenosine and adenylates as important mediators of tumor biology, and these compounds are involved in tumor metabolism and tumor immunosuppressive effects. Adenosine receptors are possible antitumor targets. We evaluated the potential utility of 11C-AMP for PET tumor imaging by using 3H-AMP to study the capacity and mechanisms of in vitro AMP uptake in various human tumor cell lines.
We found that various tumor cell lines (SKOV-3, U251, U87, SCC-15, and MN-1), when exposed to 3H-AMP in vitro, have a marked increase in intracellular uptake compared with that after exposure to 3H-FDG, a tritiated analog of 18F-FDG, which is currently the radiotracer usually used for imaging tumor metabolism. The level of intracellular uptake of 3H-AMP was more than 10–100 times the uptake of 3H-FDG in corresponding cells in vitro, depending on the particular tumor cell line—a highly incremental uptake over 3H-FDG. This high uptake, if it translates into in vivo results, may allow for very sensitive PET tumor imaging using 11C-AMP.
In order to study the mechanism of adenosine accumulation in tumor cells with high uptake of 3H-AMP, chemical inhibitors were used to evaluate specific pathways in cellular adenosine metabolism. Caffeine, a competitive inhibitor of adenosine receptor (A1, A2, and A3) binding, was used to show that competitive inhibition of the adenosine receptor did not affect the uptake of 3H-AMP in SKOV-3 cells. Adenosine receptor binding therefore does not appear to be an important mechanism of total cellular 3H-AMP uptake. However, pretreatment of the cells with dipyridamole, an inhibitor of ENTs, dramatically decreased 3H-AMP uptake in a dose-dependent fashion in SKOV-3, SCC-15, and U251 cells. This finding indicates that ENTs were responsible for most of the intracellular uptake of 3H-AMP in these particular cells. Interestingly, U87 cells did not show an appreciable inhibition of 3H-AMP uptake with dipyridamole, possibly because of an altered ENT phenotype. Expression of ENTs and CNTs in normal and neoplastic cells can be of various patterns, and these patterns may predict the effectiveness of tumor treatment (12–14).
CD73, an ecto-5′-nucleotidase, is inhibited by APCP. We were able to use APCP to demonstrate that in most of our cell lines, CD73 was necessary to dephosphorylate 3H-AMP to 3H-adenosine before intracellular uptake by a nucleoside transporter. CD73 was important for cell uptake in SKOV-3, SCC-15, and U251 cells, in which uptake of 3H-AMP was dramatically and significantly inhibited when CD73 was inhibited. However, in U87 cells, APCP only partially inhibited uptake of 3H-AMP, indicating the expression of other enzymes able to dephosphorylate 3H-AMP. CD73 has been reported to be associated with various physiologic processes, including coagulation and endothelial function, and a recent report indicated that CD73 may be involved in tumor metastases and aggressiveness.
The B-cell lymphoma cell lines (Raji and Daudi) did not take up significant 3H-AMP in our in vitro experiments. Although intracellular uptake of 3H-AMP was very low in these lymphoma cell lines, uptake of 3H-adenosine was significantly increased and was inhibited by dipyridamole and high levels of adenosine. This finding is consistent with the expression of ENTs. As would be expected, uptake of 3H-adenosine was unaffected by APCP inhibition of CD73. We were able to further demonstrate by flow cytometry using a fluorescently labeled anti-CD73 antibody that these cells had no appreciable CD73 protein expression. SKOV-3, as expected, showed significant expression of CD73 by flow cytometry. These experiments demonstrated that a lack of CD73 activity was responsible for the low 3H-AMP intracellular uptake in the Raji and Daudi cells.
Functional characterization of nucleoside transporters using 3H-adenosine indicates primarily ENT1s in SKOV-3 cells and a relative absence of Na+-dependent concentrative transporter expression in this cell type. In U87 cells, inhibition of 3H-adenosine uptake at low nanomolar and micromolar concentrations of NBMPR was minimal, indicating the presence of an ENT- and sodium-dependent uptake. This indication is consistent with the presence of a CNT. However, partial inhibition of 3H-adenosine uptake in U87 cells occurred at a 10 μmol/L concentration of dipyridamole. This finding may indicate expression of a non-ENT1 and an ENT2 subtype sensitive to dipyridamole but resistant to 0.1 nmol/L to 10 μmol/L concentrations of NBMPR inhibition because of mutation of ENT1 or ENT2 or possibly expression of another ENT subtype. These experiments further demonstrate that ENTs and CNTs can have varying patterns of expression in different tumors and that both types of transporters are involved in intracellular uptake of 3H-adenosine.
It is theorized that under specific physiologic circumstances, a coordinate regulation of enzymes may be responsible for adenosine metabolism or production resulting in an adenosine-generating or adenosine-removing cellular phenotype, with resultant high or low adenosine concentrations (3,6,7). Interestingly, B-cell lymphomas and myeloid leukemias were thought to be adenosine metabolizing whereas epithelial neoplasms were thought to be adenosine producing. Our 3H-AMP cellular uptake experiments support this hypothesis. Although adenosine-removing or -producing cancer cell phenotypes are intriguing, their significance has yet to be fully validated. We were able to show persistent intracellular uptake and retention of 3H-adenosine after it had been metabolized to predominantly 3H-ATP and 3H-ADP in our SKOV-3 cell lines. Our in vitro uptake experiments appeared to show that cancer cells may exhibit intracellular uptake and retention of 3H-adenosine metabolites.
These preliminary in vitro results show that in addition to the glucose analog, FDG, other molecules involved in cellular metabolism may be used as a marker of tumor metabolism. FDG is thought to be especially powerful because it is a downstream and later marker of tumor cell adaptation to hypoxia and of cellular transformation to a more aggressive phenotype. Nucleoside and nucleotide metabolic pathways have been studied extensively during the past 20–30 y, primarily because of their importance in tumor biology and because of their role in the development of nucleoside analogs as chemotherapeutic drugs in oncology. Nucleosides and nucleotides may prove to be a fertile area in which to develop new radiotracers for metabolic tumor imaging.
CONCLUSION
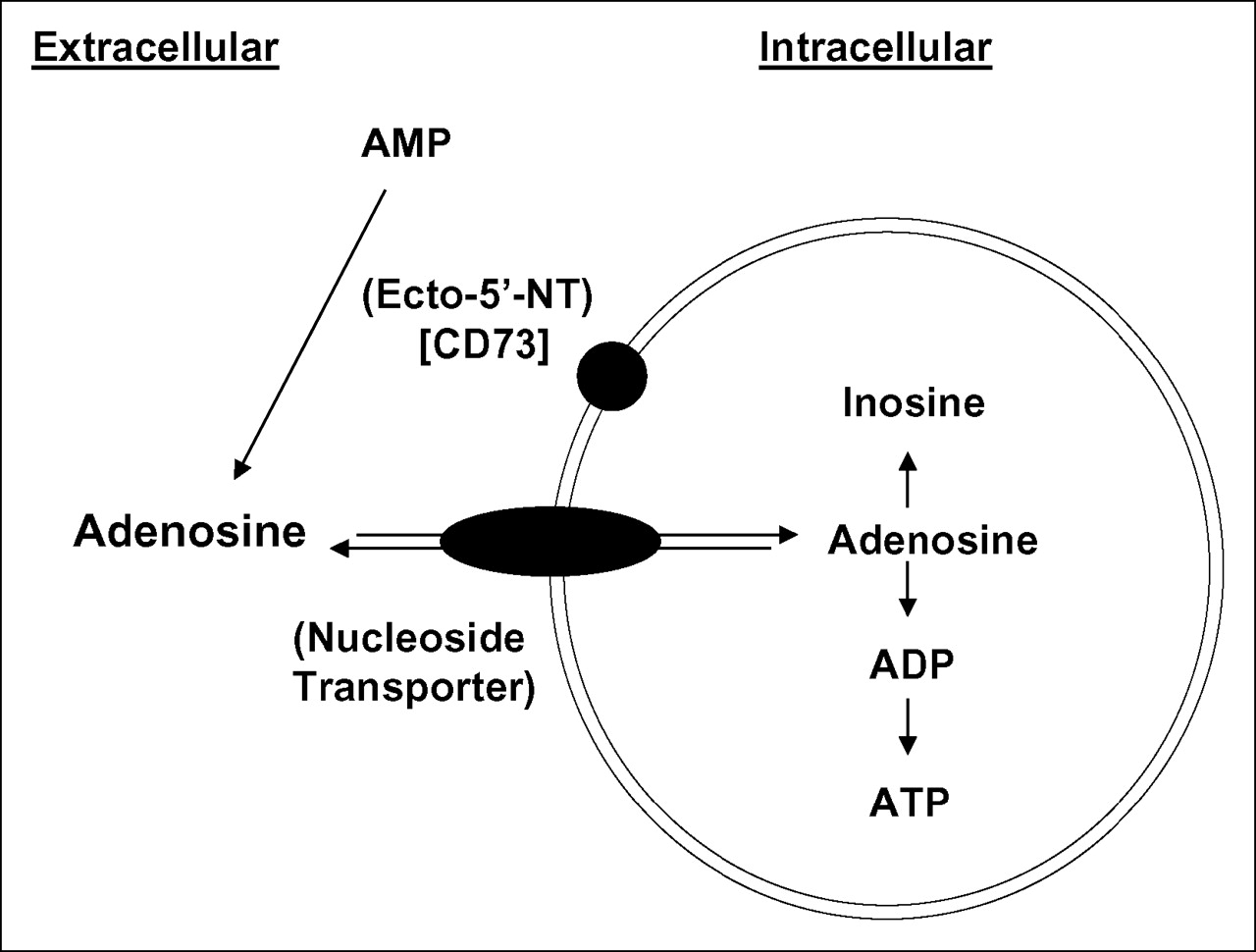
In this study, several human cancer cell lines (SKOV-3, SCC-15, U251, and U87) were evaluated in vitro and demonstrated marked and significantly greater uptake of 3H-AMP than of 3H-FDG. Radiolabeled adenosine showed that the mechanism of intracellular uptake involved dephosphorylation of AMP by ENTs or CNTs. 3H-AMP uptake was dependent predominantly on ENTs after conversion of AMP to adenosine by CD73 in SKOV-3, SCC-15, and U251 cells and specifically on ENT1 in SKOV-3 cells (Fig. 7). In U87 cells, uptake of radiolabeled AMP appeared to be via both an equilibrative and a Na+-dependent CNT mechanism. Raji and Daudi B-cell lymphoma cell lines, however, showed low uptake of 3H-AMP because of a lack of CD73 expression, with the resulting inability to dephosphorylate AMP to adenosine, but these cells had high intracellular uptake of 3H-adenosine. Uptake experiments demonstrated persistent retention of 3H-adenosine metabolites after influx into the cell. High-performance liquid chromatography of the intracellular 3H pool showed conversion to 3H-ATP and 3H-ADP. Our in vitro work demonstrates the potential of 11C-AMP for use in metabolic tumor imaging with PET. Studies are currently under way to further investigate the in vivo applicability of 11C-AMP.
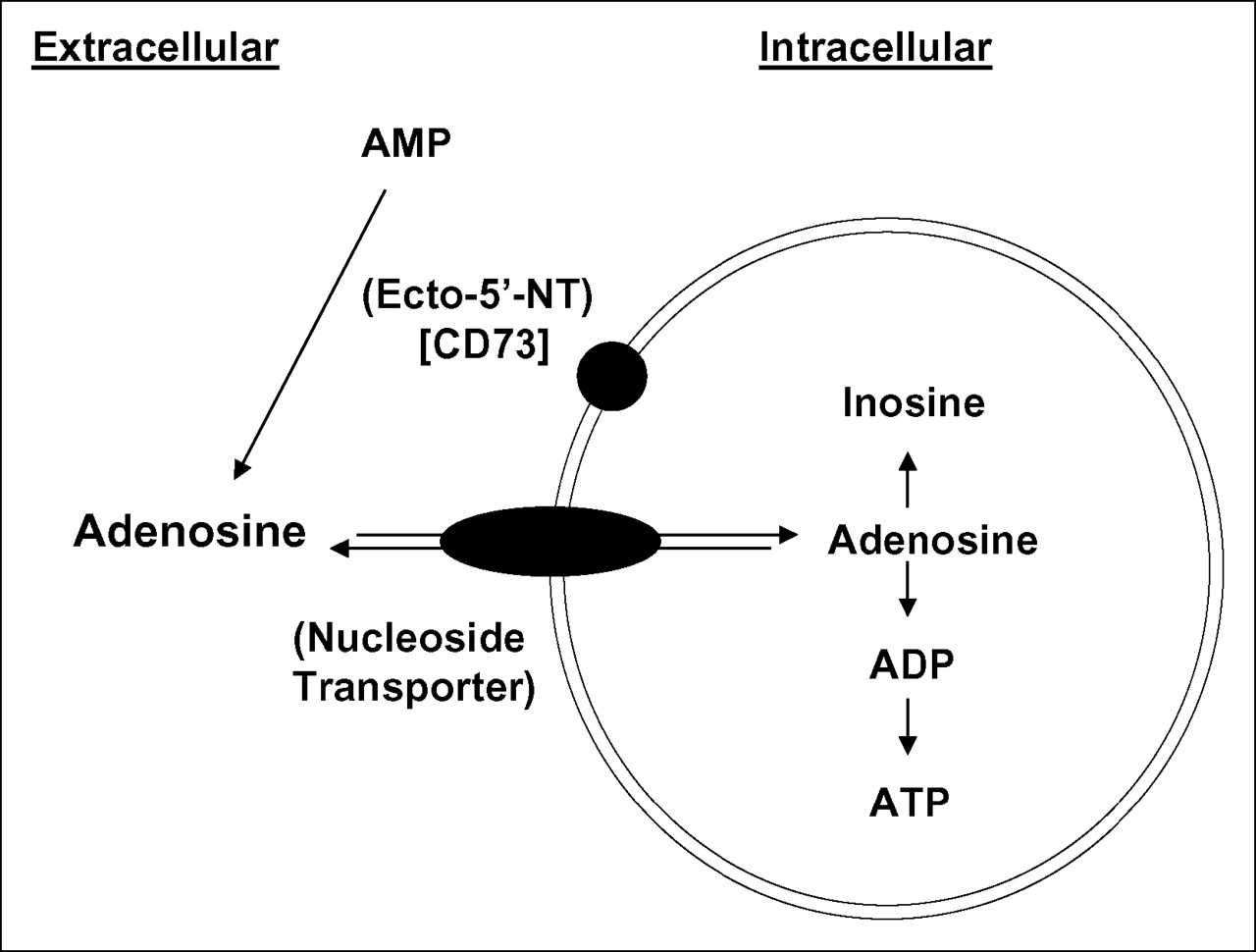
Schematic diagram of proposed mechanism of intracellular uptake of 3H-AMP based on the in vitro experiments performed in this project.
Acknowledgments
This study was supported in part by NCI grant CA 52880 and by an NIH extramural loan repayment grant. We thank Nicole Benoit for her excellent help and expertise with the CD73 flow cytometry experiment and Judy Buchanan for her editorial help.
References
- Received for publication September 20, 2005.
- Accepted for publication January 30, 2006.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}