


Abstract
Auger electrons can create breaks in nucleic acids, giving them possible therapeutic utility. We investigated the therapeutic effect of Auger electrons emitted by 111In-labeled phosphorothioate antisense oligonucleotides on human neuroblastoma cells in which N-myc was overexpressed. Methods: Human SK-N-DZ neuroblastoma cells (5 × 106 cells) were treated with cationic reverse-phase evaporation vesicles (REVs) encapsulating 111In-labeled antisense (40 MBq/2 nmol of oligonucleotides/μmol of total phospholipids) that had an average diameter of 250 nm. Hybridization of the radiolabeled oligonucleotides with N-myc messenger RNA (mRNA), N-myc expression, and cell proliferation were investigated. The tumorigenicity of treated cells was analyzed in nude mice. Nonradiolabeled antisense, 111In-labeled sense, or empty cationic REVs were used as controls. Results: 111In-Labeled antisense, which hybridized with N-myc mRNA, was detected in cells at 12 and 24 h after the initiation of treatment. Reduced N-myc expression and inhibited cell proliferation were shown in the same cells at 48 h after the completion of treatment. N-myc expression–suppressed cells produced intraperitoneal tumors in nude mice, but the average weight of the tumors was lower than that of tumors in control mice. Conclusion: Auger electrons emitted from 111In in close proximity to their target N-myc mRNA may prolong the time to cell proliferation in human neuroblastoma cells due to inhibition of the translation of N-myc. Auger electron therapy therefore has potential as an internally delivered molecular radiotherapy targeting the mRNA of a tumor cell.
Deregulation of members of the family of myc genes (the most common of which are c-myc, N-myc, and L-myc) that encode transcription factors is found in a variety of human and animal tumors (1). Association of the N-myc gene with human tumor may be striking in neuroblastoma, a tumor of neural crest origin that occurs predominantly in early childhood, in which the N-myc gene is amplified in about 20%–25% of human neuroblastoma (2). The N-myc overexpression as well as the age of patients with neuroblastoma, clinical stage of disease, and regional lymph node involvement at the diagnosis are associated with poor prognosis (3–5). It has been hypothesized that deregulation of the N-myc gene is causative in the formation of neuroblastoma; evidence in support of this hypothesis was found in a study of transgenic mice, in which targeted expression of N-myc in vivo caused neuroblastoma to develop (6). Conversely, modulation of N-myc expression has been shown to have biologic effects—for instance, reducing the cell growth rate and inducing differentiation in human neuroblastoma cell lines. Targeted inhibitors of N-myc are being sought so that they can be used in the development of specific therapeutic agents for neuroblastoma with N-myc overexpression. To date, the biologic effects of N-myc modulators have been achieved by antisense oligonucleotides targeted against N-myc messenger RNA (mRNA) (7–9), expression vectors designed to generate N-myc antisense RNA (7,10), and antisense peptide nucleic acids (PNAs) against N-myc mRNA (PNA is an analog of DNA in which the backbone is a pseudopeptide rather than a sugar) (11). Each of these modulators has caused the translation of N-myc to be inhibited.
Auger electrons are a widely recognized potential candidate for use in targeted cancer therapies. DNA damage caused by Auger electrons emitted by an atom undergoing electron capture (EC) (12–16) could directly affect the oncogenes involved in the cancer's development or growth. The commercially available atoms used as Auger electron emitters are 125I and 111In. The yield of DNA strand breaks by 125I has been demonstrated to strongly depend on the distance between the decay site and the DNA (13), with greater damage being inflicted when the radionuclide is close to the gene site than when it is further away. Therefore, to estimate the extent of DNA damage in terms of strand breaks, not only the average energy of the Auger electrons but also the distance from the radionuclide to the DNA strands should be considered (17). Presumably, N-myc mRNA would also be broken by Auger electrons as internal radiation would be expected to cause RNA as well as DNA damage (18). Such damage similarly could result in suppressed N-myc expression.
In the present study, we investigated the therapeutic effect of Auger electrons on human neuroblastoma cells in which N-myc was overexpressed. Antisense oligonucleotides designed to bind to N-myc mRNA through Watson–Crick hybridization were used as a vehicle, with the assistance of cationic reverse-phase evaporation vesicles (REVs), to ensure the delivery of the Auger electrons emitted by 111In into the human neuroblastoma cells.
MATERIALS AND METHODS
Preparation of 111In-Labeled Oligonucleotides
The sequence of the N-myc 15-mer phosphorothioate sense oligonucleotide was 5′-ATG CCG GGC ATG ATC-3′; it corresponded to the region of the N-myc mRNA beginning with the ATG start codon at position 1,650 (19). The sequence of the N-myc 15-mer phosphorothioate antisense oligonucleotide was 5′-GAT CAT GCC CGG CAT-3′, with replacement of C-5–substituted deoxyuridine with T in the T position to conjugate isothiocyanobenzyl ethylenediaminetetraacetic acid (SCN-Bn-EDTA) (Fig. 1). The substituent at the C-5 position of T was N-1-acetylhexamethylenediamine (20). Oligonucleotides were assembled using an ABI 381-A DNA synthesizer (Perkin-Elmer) by means of the solid-phase phosphoramidite method, and Bn-EDTA–conjugated oligonucleotides were prepared as described by Watanabe et al. (21).
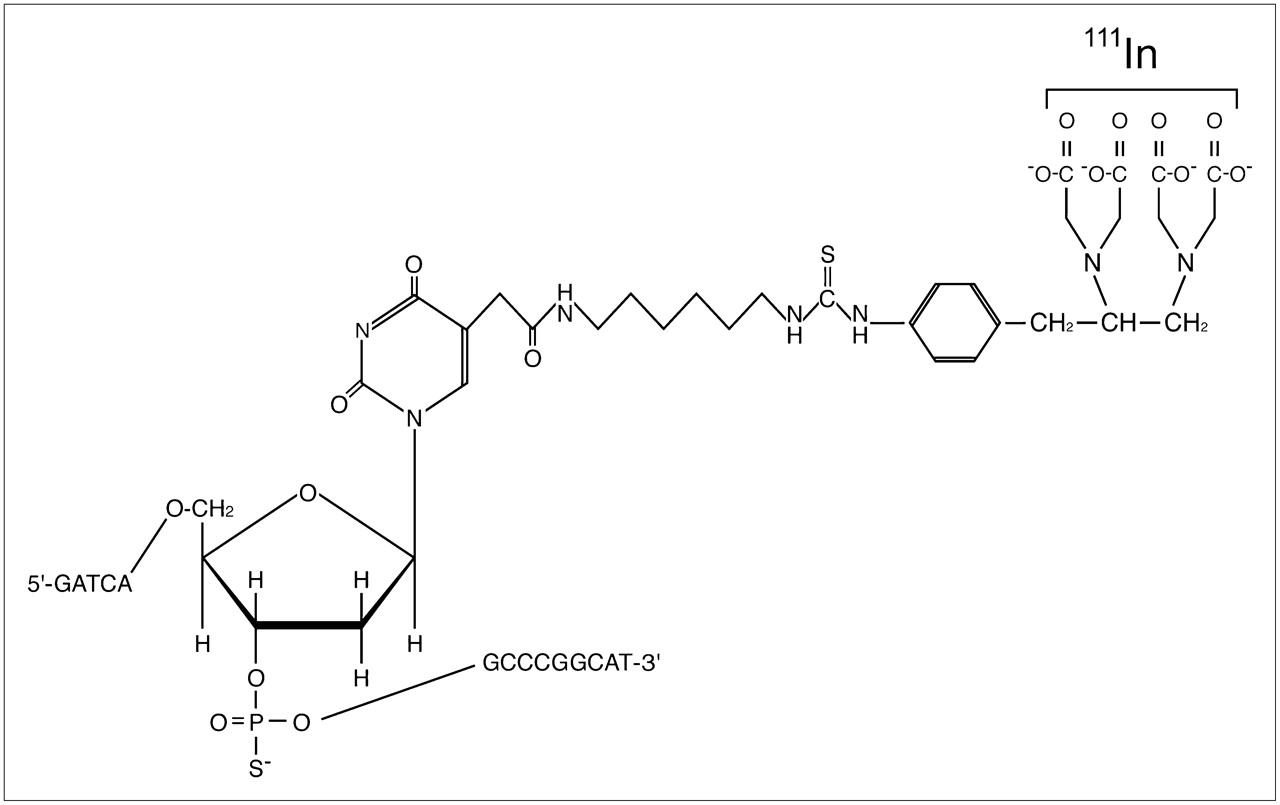
Structure of 111In-Bn-EDTA-phosphorotioate antisense oligonucleotides.
The Bn-EDTA–conjugated oligonucleotides in 100 mmol/L citrate buffer (pH 6.0) were mixed with 111InCl3 (Nihon Medi-Physics Co., Ltd.) at a 15:1 molar ratio for 30 min at room temperature. Disodium salt EDTA (Na2EDTA) (Dojin Chemical Co., Ltd.) to a final concentration of 5 mmol/L was added to the reaction solutions to chelate unbound 111In. The samples were subjected to 3 rounds of Sephadex G-25 (Amersham Biosciences) gel filtration in phosphate-buffered saline (PBS) without magnesium and calcium. The appropriate fractions were applied to a centrifugal concentrator (CC-105; Tomy Seiko Co., Ltd.), and the concentration of 111In-Bn-EDTA–conjugated oligonucleotides (111In-labeled oligonucleotides) was adjusted to 410 μmol/L in PBS. The labeling efficiency was approximately 85%. The range of specific radioactivities of 111In-labeled oligonucleotides was 19.7∼20.3 MBq/nmol. The radioactivity was measured with a γ- and β-counting calibrator (Capintec), and the concentration of oligonucleotides was calculated from their ultraviolet absorbance at 260 mm.
The ability of the 111In-labeled antisense and sense and their precursors to hybridize with the synthesized 15-mer phosphorodiester sense (5′-ATG CCG GGC ATG ATC-3′) and antisense (5′-GAT CAT GCC CGG CAT-3′) oligonucleotides was assessed by measuring the melting temperature (Tm) with a DU-650Tm programmable spectrophotometer (Beckman Coulter Inc.). The Tm was 54.4°C ± 0.3°C (mean ± SD of 3 independent experiments) of 111In-labeled antisense and 54.5°C ± 0.2°C of 111In-labeled sense, whereas the Tm was 54.7°C ± 0.3°C of phosphorothioate antisense and 54.6°C ± 0.3°C of phosphorothioate sense. 111In-Labeled oligonucleotides to a final concentration of 2.3 μmol/L were incubated with fresh human normal serum or PBS for 96 h at 37°C in a humidified 5% CO2/95% air atmosphere, and the stability was assessed by means of reverse-phase high-performance liquid chromatography (HPLC). As free 111In from 111In-labeled antisense and sense, 2.2% ± 0.2% and 2.1% ± 0.2% of the total radioactivity, respectively, was found in serum and 1.2% ± 0.2% and 1.3% ± 0.2%, respectively, was found in PBS. The half-lives (t1/2) of 111In-labeled antisense and sense were 12.2 ± 0.3 h and 12.1 ± 0.2 h in serum, respectively.
Preparation of Cationic REVs Encapsulating 111In-Labeled Oligonucleotides
N-(α-Trimethylammoniumacetyl)didodecyl-d-glutamate chloride (Sogo Pharmaceutical Co., Ltd.), dilauroyl phosphatidylcholine (Sigma Chemical Co.), l-α-diarachidoyl phosphatidylcholine (Avanti Polar-Lipids, Inc.), and dioleoyl phosphatidylethanolamine (Avanti Polar-Lipids) in a molar ratio of 1:1:1:2 were dissolved in chloroform. Positively charged REVs were prepared according to the method described by Szoka and Papahadjopoulos (22). The lipid mixture was added to a 5-mL round-bottom flask with a long extension neck, and the solvent was removed under reduced pressure by a rotary evaporator. The system was then purged with nitrogen, and lipids were redissolved in isopropyl ether and ethanol at a volume ratio of 9:1 to obtain a final lipid concentration of approximately 10 mmol/L. 111In-Labeled oligonucleotides in PBS (3.6 μmol/L) were added at a volume ratio of 0.25 to the organic phase at this point, and the system was kept continuously under nitrogen. The resulting 2-phase system was sonicated intermittently and briefly (10–15 s) in a bath-type sonicator until the mixture became a 1-phase dispersion. The mixture was then placed on the rotary evaporator, and the solvents were removed under reduced pressure (generated by a water aspirator) at 25°C, with rotation at approximately 200 rpm.
The REV preparation was extruded 10 times through 2 stacked Nucleopore polycarbonate filters (0.2 μm) (Whatman Inc.) in a 10-mL capacity thermobarrel extrusion (Lipex Biomembranes).
The REV preparation was then mixed with 30% (w/v) Ficoll in PBS and transferred to a sterile cellulose nitrate centrifuge tube. Ten percent (w/v) Ficoll and PBS (in a 3:1 volume ratio) were gently layered consecutively onto the sample, and the step gradient was centrifuged at 100,000g for 30 min (Optima XL-80K centrifuge and SW55Ti rotor; Beckman Coulter Inc.) to separate cationic REVs encapsulating 111In-labeled oligonucleotides from unencapsulated radiolabeled oligonucleotides. Cationic REVs encapsulating 111In-labeled oligonucleotides with 39.4∼40.6 MBq/μmol of total phospholipid were obtained in PBS.
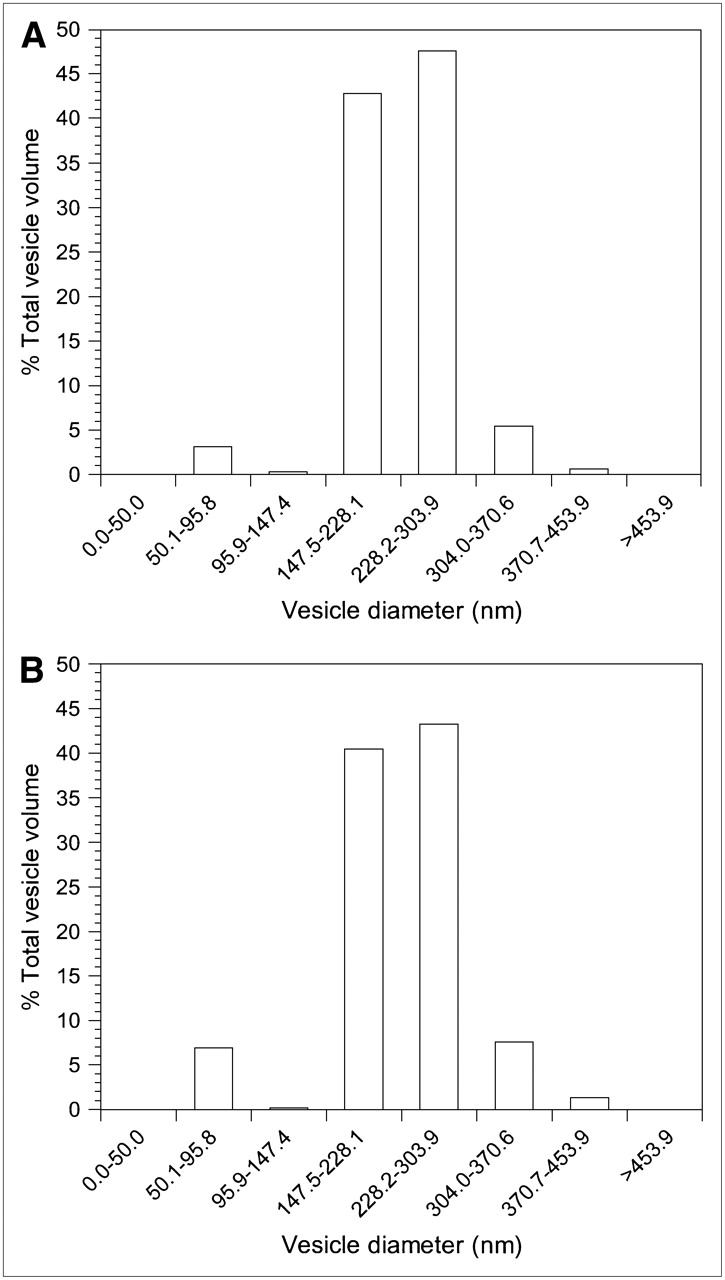
The average diameters of the cationic REVs encapsulating 111In-labeled antisense and sense were measured as 253.8 ± 10.5 nm and 247.1 ± 11.4 nm, respectively, by light scattering spectrophotometry (DLS-700 spectrophotometer; Otsuka Electronics Co., Ltd.) (Fig. 2). Cationic REVs encapsulating 111In-labeled oligonucleotides to a final concentration of 1.14 μmol/L of total phospholipid were incubated with fresh human normal serum or PBS for up to 24 h at 37°C, and the stability was assessed by means of a Sepharose CL-4B column (250 mm × 8 mm; Pharmacia). The cationic REVs retained 90.0% ± 0.5% and 90.4% ± 0.5% of the encapsulated 111In-labeled antisense and sense in serum, respectively, over 24 h and 96.9% ± 0.2% and 96.6% ± 0.2% in PBS, respectively.
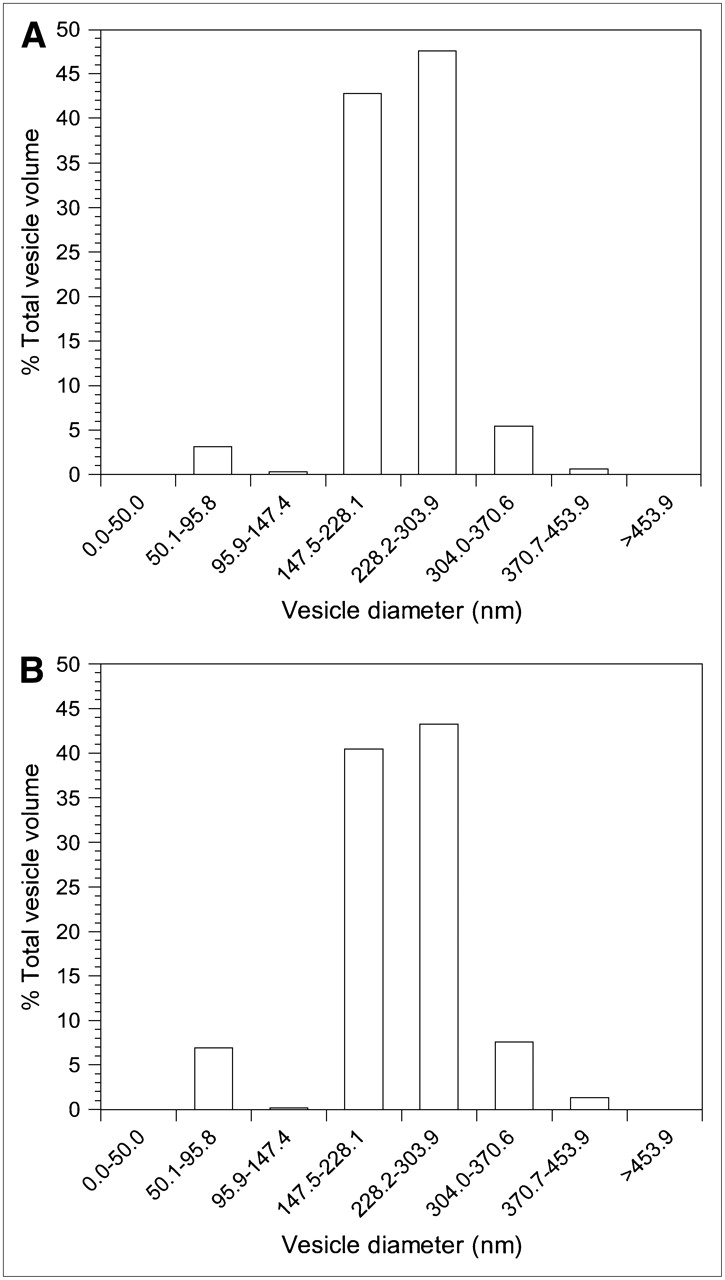
Size distribution of cationic REVs encapsulating 111In-labeled antisense (A) and 111In-labeled sense (B) determined by light-scattering spectrophotometer.
Detection of 111In-Labeled Oligonucleotides Hybridized with N-myc mRNA of Neuroblastoma Cells
A human neuroblastoma cell line (SK-N-DZ) was obtained from the American Type Culture Collection and maintained at 37°C in a humidified 5% CO2/95% air atmosphere in Dulbecco's modified Eagle medium (DMEM) supplemented with 4.5 mg/L glucose, 0.1 mmol/L nonessential amino acids, and 10% fetal calf serum inactivated for 30 min at 65°C. When the SK-N-DZ cells reached logarithmic-phase growth, they were seeded in 145-mm dishes at a density of 5 × 106 cells per dish and maintained in supplemented DMEM at 37°C in a 5% CO2 atmosphere for 24 h. Cationic REVs encapsulating 111In-labeled oligonucleotides (1 μmol of total phospholipid) were added to the dishes and incubated at 37°C in a 5% CO2 atmosphere for 20 h. After REVs were then removed by washing in PBS containing 0.25% (w/v) trypsin/0.53 mmol/L EDTA, the cells were maintained in fresh supplemented DMEM at 37°C in a 5% CO2 atmosphere for a further 28 h.
At 0, 12, 24, and 48 h after their exposure to cationic REVs encapsulating 111In-labeled oligonucleotides, cell samples were washed with 0.25% (w/v) trypsin/0.53 mmol/L EDTA in PBS and all cells in a dish were collected in a tube. The level of radioactivity was determined using a γ-counter (Aloka). Total RNA, including mRNA hybridized with radiolabeled oligonucleotides, was then extracted from the cells using the acid guanidinium thiocyanate/phenol/chloroform method (23). The nucleic acids were incubated in S1 nuclease buffer (280 mmol/L NaCl, 30 mmol/L sodium acetate [pH 4.4], and 4.5 mmol/L zinc acetate) with 20 μg/mL sonicated salmon sperm DNA, containing 200 U of S1 nuclease (Takara Bio Inc.) at 37°C for 30 min. The digestion was terminated by adding 2.5 mol/L ammonium acetate and 50 mmol/L EDTA, and the sample was then fractionated by alkaline agarose gel electrophoresis (5.0% NuSieve GTG gel [FMC BioProducts], 50 mmol/L NaOH, and 1 mmol/L EDTA). Intact 111In-labeled antisense and sense were also loaded as controls. 111In-Oligonucleotides were transferred in alkaline transfer buffer (0.4N NaOH and 1 mol/L NaCl) to a Hybond-N+ nylon membrane (Amersham) by means of the capillary transfer method. The membrane was reacted with 32P-labeled phosphorodiester sense oligonucleotides (2 × 106 cpm/mL) in hybridization buffer (prehybridization buffer and 50% dextran sulfate in a 5:1 ratio) at 42°C for 12 h. Phosphorodiester sense (5′-ATG CCG GGC ATG ATC-3′) and antisense (5′-GAT CAT GCC CGG CAT-3′) oligonucleotides were assembled by the synthesizer, and 5′-end labeling with [γ-32P]ATP (111 TBq/mmol; Perkin-Elmer Life & Analytic Sciences, Inc.) was conducted with T4 polynucleotide kinase (Takara Bio Inc.). The radioactivity on the membrane was detected and quantified using an imaging phosphor plate system (BAS-1800II; Fuji Fikm Co., Ltd.) after sufficient decay of the radioindium activity of the samples. Quantitative linearity between the dose (pmol) of 111In-labeled oligonucleotides and the value of photostimulated luminescence was maintained. The recovery ratio of 111In-labeled antisense that underwent in vitro hybridization with a 60-mer RNA probe corresponding to the region of N-myc mRNA beginning with the ATG start codon at position 1,650) was 9.14% ± 0.16%. Dose corrected by the recovery ratio was expressed as the mean ± SD of 3 independent experiments.
Effects of 111In-Labeled Oligonucletides on N-myc Expression in Cells and Cell Proliferation
SK-N-DZ cells in logarithmic-phase growth were seeded in 145-mm dishes at a density of 5 × 106 cells per dish and maintained in supplemented DMEM at 37°C in a 5% CO2 atmosphere for 24 h. Cationic REVs encapsulating 111In-labeled oligonucleotides (1 μmol total phospholipid) or cationic REVs encapsulating nonradiolabeled oligonucleotides (1 μmol total phospholipid) that had been prepared using the methods described above were added to the dishes, which then were incubated at 37°C in a 5% CO2 atmosphere for 20 h. After REVs were removed by washing with 0.25% (w/v) trypsin/0.53 mmol/L EDTA in PBS, the cells were maintained in fresh supplemented DMEM at 37°C in a 5% CO2 atmosphere. As a control, treatment with empty cationic REVs was performed.
At 24, 48, and 72 h after the completion of treatment with cationic REVs encapsulating 111In-labeled oligonucleotides or nonradiolabeled oligonucleotides, 5 × 106 cells per dish were collected into a microtube on ice and lysed in ice-cold TNE buffer (10 mmol/L Tris-HCl [pH 7.8], 1% Nonidet P-40, 150 mmol/L NaCl, and 1 mmol/L EDTA) containing 10 μg/mL protease inhibitor aprotinin and 200 nmol/L phosphatase inhibitor sodium orthovanadate. Protein concentrations were determined by dye-binding assay (Bio-Rad Laboratories). The volume of protein solution (100 μg of total cell protein) adjusted with TNE buffer was immunoprecipitated with anti-N-myc mouse monoclonal antibody (clone NCM II 100; Calbiochem) to a concentration of 5 μmol/L, and it was subjected to 10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis. Proteins were transferred onto a polyvinylidene difluoride membrane for 1 h in transfer buffer (100 mmol/L Tris [pH 8.3] and 200 mmol/L glycine) with a protein-blotting semidry system (BE-300; Bio Craft). The membrane was incubated with the anti-N-myc monoclonal antibody, and the antibodies were detected with rabbit antimouse IgG (Fc specific) (Fluka Chemie AG) labeled with 125I (Amersham) by means of the chloramine-T method (24). The radioactivity was detected using the imaging phosphor plate system. The expression rate (given as a percentage) was calculated by dividing the 125I activity of N-myc in treated cells by that of control cells and was expressed as the mean ± SD of 3 independent experiments. Cell count and viability were determined by trypan blue exclusion at 24, 48, and 72 h after treatment. Each experiment was performed in quadruplicate: One pair of samples was tested to determine the effect of the radionuclide on N-myc expression and the other pair was tested for the radionuclide's effect on cell proliferation.
Intraperitoneal Tumorigenicity of Human Neuroblastoma Cells with Cationic REVs Encapsulating 111In-Labeled Antisense Oligonucleotides in Nude Mice
Six-week-old female athymic nude mice (n = 45) with a BALB/c/nu/nu background (Clea Japan, Inc.) were used as an animal model for neuroblastoma in which to test the effects of 111In-labeled oligonucleotides. This animal study was performed in compliance with the institutional guidelines and rules governing animal care and use.
Each mouse was injected intraperitoneally with 500 μL of 2,6,10,14-tetramethyl pentadecane (Pristan; Aldrich Chemical Co.) and was inoculated intraperitoneally with 5 × 106 SK-N-DZ cells 1 wk later. The mice were divided equally into 3 groups, and then the cells treated with cationic REVs encapsulating 111In-labeled antisense and nonradiolabeled anisense were inoculated intraperitoneally into the first 2 groups, respectively, whereas the cells treated with empty cationic REVs formed the third group as a control.
Seven, 14, and 21 d after inoculation, 5 mice in each group were killed while under general anesthesia with diethylether, and tumors on the peritoneum were collected and weighed. Data obtained for each group at each time point were expressed as the mean ± SD for 5 mice.
Data Analysis
Statistical comparisons were made using the Student t test, and differences between groups were considered statistically significant when the P value was <0.01.
RESULTS
Detection of 111In-Labeled Oligonucleotides Hybridized with N-myc RNA in Human Neuroblastoma Cells
111In-Labeled antisense was detectable at 12 and 24 h after the initiation of treatment (Fig. 3). For 111In-labeled antisense, the levels of hybridization with N-myc mRNA in the tumor cells were 135.7 ± 6.6 pmol at 12 h and 142.2 ± 8.5 pmol at 24 h; 111In-labeled sense was undetectable at all time points (Fig. 3).

Detection of 111In-labeled oligonucleotides hybridized with N-myc mRNA in human neuroblastoma cells. 111In-Labeled antisense (111In-AS) hybridized with N-myc mRNA at 12 and 24 h after they were exposed to cells by means of transport in cationic REVs; 111In-labeled sense (111In-S) did not hybridize with N-myc mRNA. Arrow indicates radiolabeled oligonucleotides.
Effects of 111In-Labeled Oligonucleotides on N-myc Expression in Human Neuroblastoma Cells and Cell Proliferation
As shown in Figure 4, N-myc expression was suppressed in human neuroblastoma cells at 48 h after treatment with cationic REVs encapsulating 111In-labeled antisense, but it gradually recovered. However, no change in N-myc expression was demonstrated in the cells treated with cationic REVs encapsulating nonradiolabeled antisense at 72 h after treatment. In addition, no difference in N-myc expression was noted between the cells treated with cationic REVs encapsulating nonradiolabeled antisense and the control cells (data not shown). Furthermore, no significant difference in N-myc expression was demonstrated between the cells treated with cationic REVs encapsulating 111In-labeled sense and those exposed to nonradiolabeled antisense (Fig. 5).
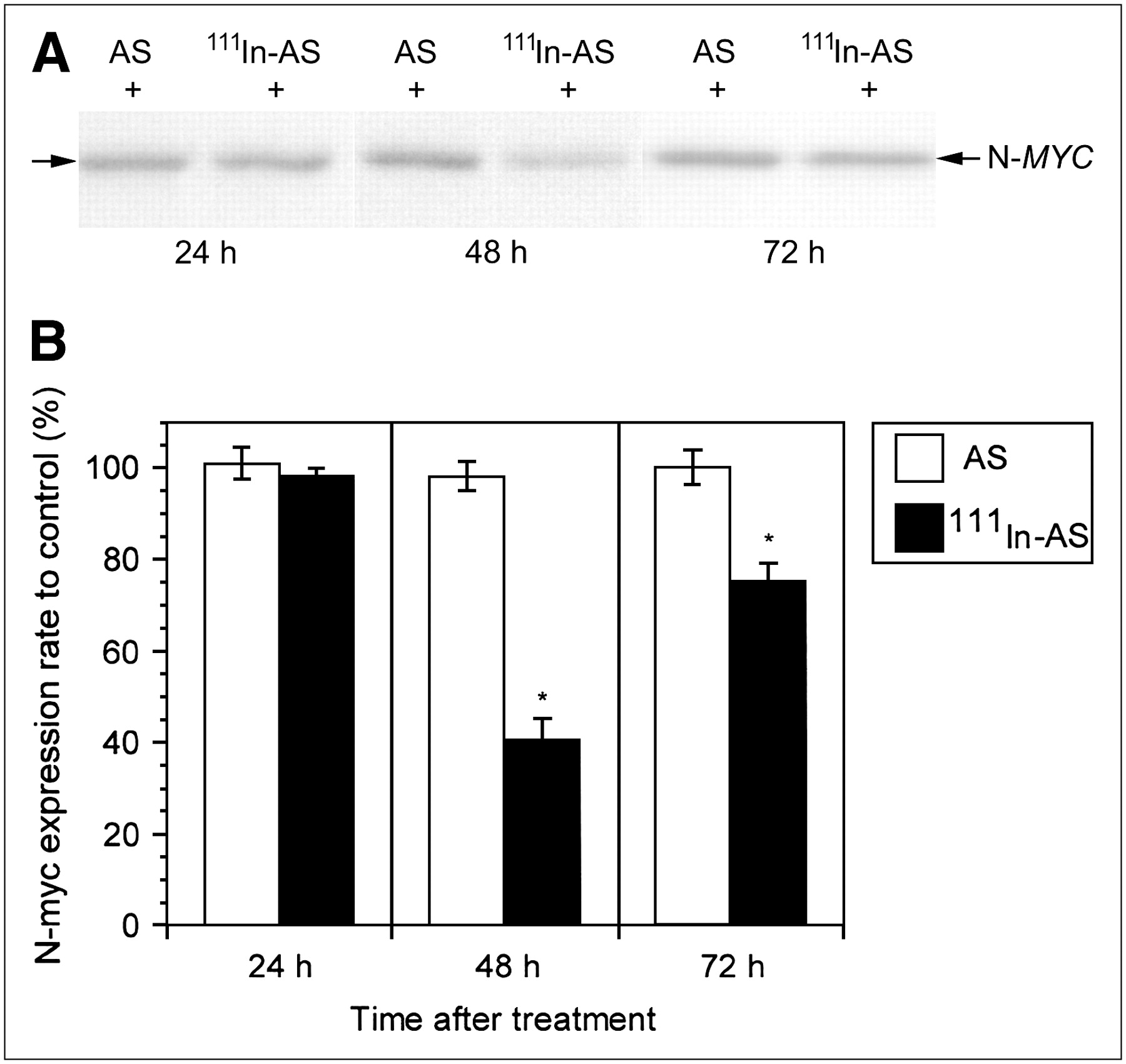
N-myc expression (N-MYC) in human neuroblastoma cells treated with cationic REVs encapsulating 111In-labeled antisense (111In-AS) and nonradiolabeled antisense (AS). (A) Western blot analysis. (B) N-myc expression ratio of treated cells to control. Reduction in N-myc expression was observed in cells treated with cationic REVs encapsulating 111In-labeled antisense at 48 h after completion of treatment. However, nonradiolabeled antisense did not confer a significant change in N-myc expression in cells. *P < 0.01 compared with ratio of nonradiolabeled antisense.
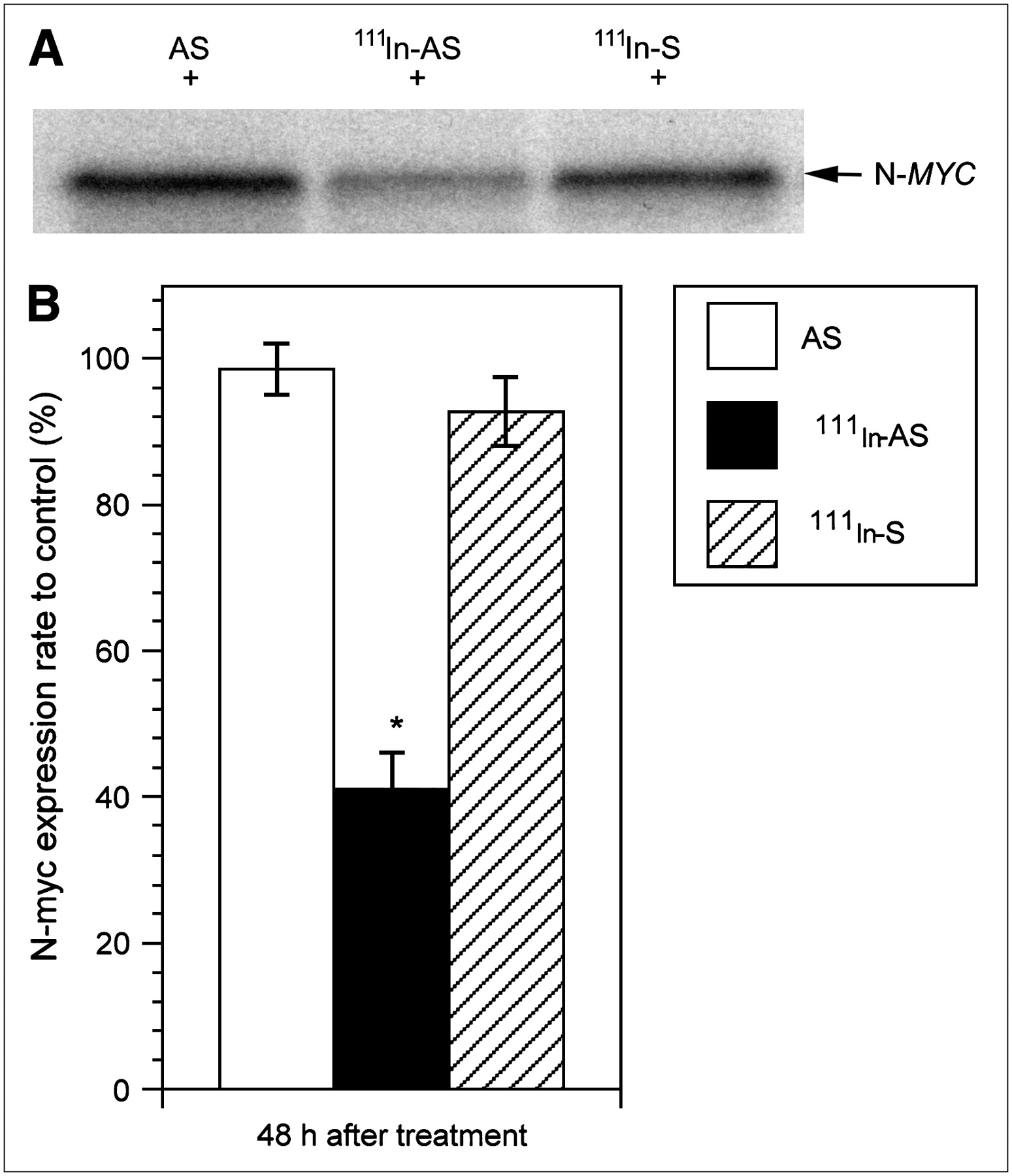
N-myc expression (N-MYC) in human neuroblastoma cells treated with cationic REVs encapsulating 111In-labeled sense (111In-S). (A) Western blot analysis. (B) N-myc expression ratio of treated cells to control. No significant difference in N-myc expression was observed between these cells and those treated with cationic REVs encapsulating nonradiolabeled antisense (AS), whereas 111In-labeled antisense (111In-AS) had shown effects at 48 h after treatment. *P < 0.01 compared with ratio of nonradiolabeled antisense.
Proliferation of the cells treated with cationic REVs encapsulating 111In-labeled antisense was reduced at 48 h after treatment (Fig. 6). No difference in cell proliferation was observed between the cells treated with cationic REVs encapsulating nonradiolabeled antisense and the control cells (data not shown). The viability of the treated cells remained at >96% at 72 h after treatment.
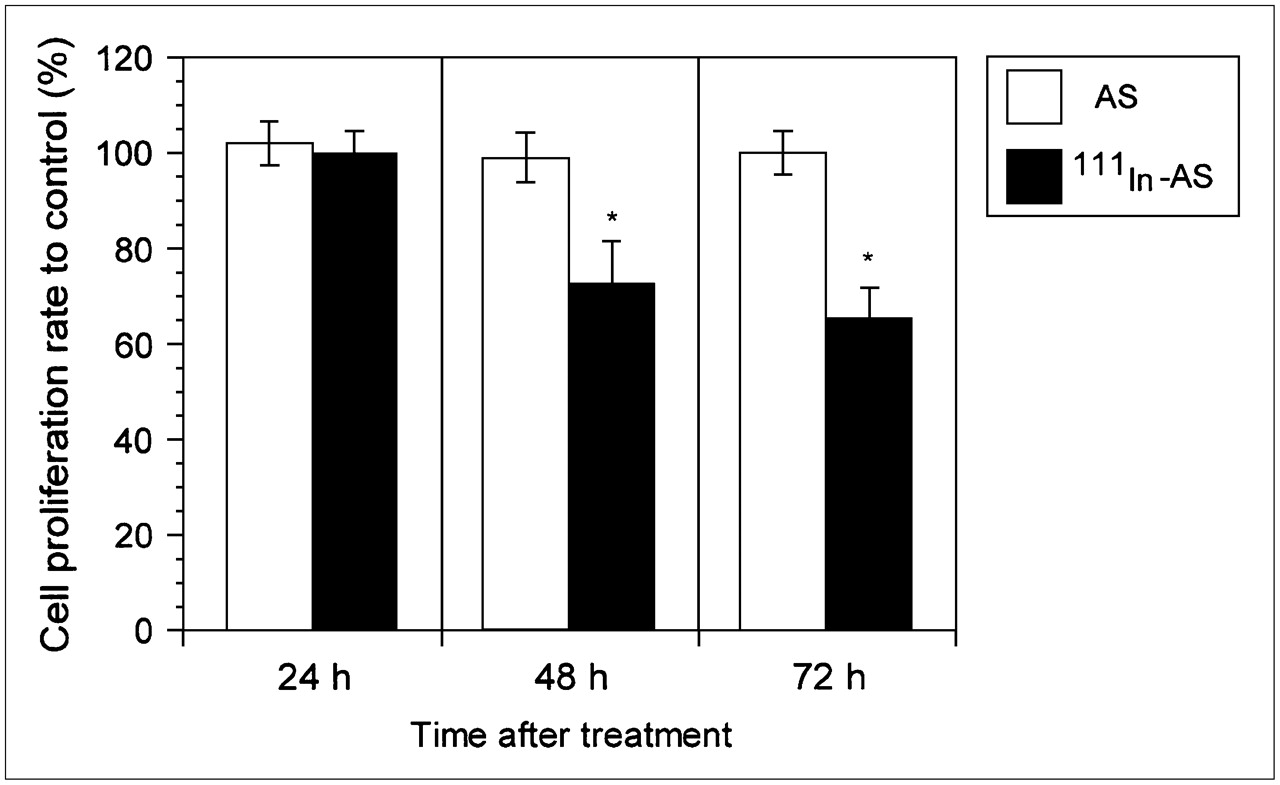
Cell growth in human neuroblastoma cells treated with cationic REVs encapsulating 111In-labeled antisense (111In-AS) and nonradiolabeled antisense (AS). Cell proliferation was inhibited at 48 h after treatment of the cells with cationic REVs encapsulating radiolabeled antisense, whereas cationic REVs encapsulating nonradiolabeled antisense had no effect on cells. *P < 0.01 compared with ratio of nonradiolabeled antisense.
Intraperitoneal Tumorigenicity of Human Neuroblastoma Cells
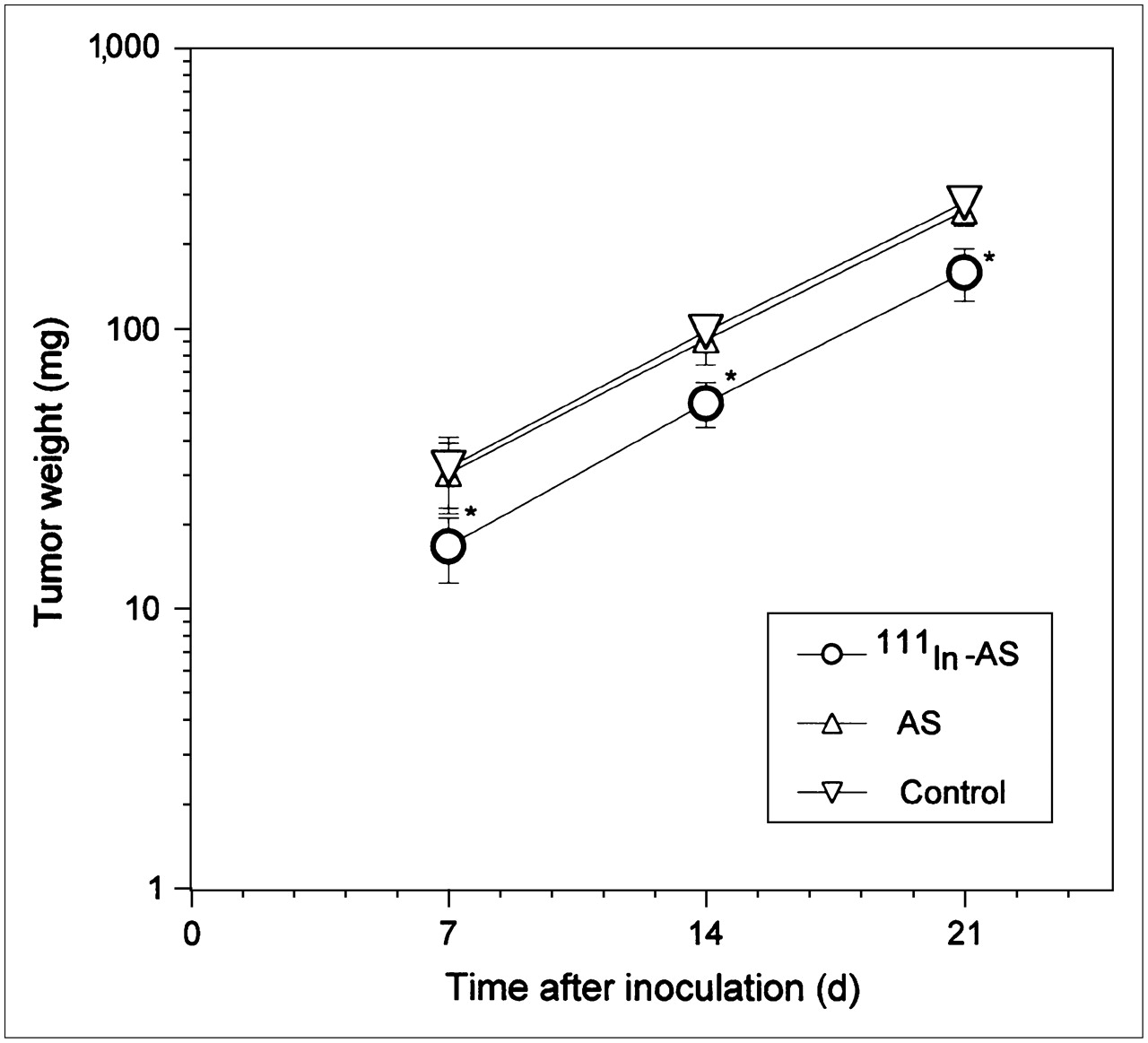
Figure 7 shows that the cationic REVs encapsulating 111In-labeled antisense caused the average weight of intraperitoneal tumors produced by neuroblastoma cells to decline to 48% of control or nonradiolabeled antisense treatments, although the treated cells could produce tumors in the mice. No differences in tumorigenicity were found between the cells treated with cationic REVs encapsulating nonradiolabeled antisense and empty cationic REVs as a control.

Tumorigenicity of human neuroblastoma cells treated with cationic REVs encapsulating 111In-labeled antisense (111In-AS) and nonradiolabeled antisense (AS). Cells treated with cationic REVs encapsulating radiolabeled antisense produced tumors in mice. However, average weight of tumors was lower than that of tumors produced by cells treated with cationic REVs encapsulating nonradiolabeled antisense and empty cationic REVs as a control. *P < 0.01 compared with control.
DISCUSSION
In this study, we used Auger electrons emitted from 111In-labeled antisense that had been encapsulated in cationic REVs to investigate the therapeutic effects on a human neuroblastoma cell line in which N-myc was overexpressed. 111In-labeled antisense reduced N-myc expression in the cells and slowed cell proliferation, whereas neither nonradiolabeled antisense nor 111In-labeled sense did. Thus, Auger electrons that were released in close proximity to their mRNA target did cause a therapeutic effect on human neuroblastoma cells.
Because the reverse-phase evaporation method was devised for the preparation of large unilamellar vesicles (LUVs) into which DNA can be encapsulated, LUVs have been explored as a carrier for gene transfer (25–28). Modification of the positive charge of liposomes, considering that both DNA and cell surfaces are negatively charged, has enhanced gene transfer in cells in vitro (29–31). In this study, cationic REVs were prepared to (a) effectively transfer 111In-oligonucleotides, (b) enhance cellular uptake of 111In-oligonucleotides, and (c) minimize extracellular degradation of 111In-oligonucleotides. Moreover, these cationic REVs may result in high tumor uptake in various tumor animal models (data not shown). Unilamellar liposome vesicles can be characterized in terms of vesicle size, as well as lipid composition, dose, and vesicle surface characteristics, such as charge and hydrophilicity (32). Small unilamellar vesicles (SUVs) are normally <0.1 μm in diameter, but the trapping efficiency of these materials is generally lower than that of LUVs, which range from 0.1 to 1 μm in diameter. Reverse-phase vesicle preparation can produce LUVs that are widely heterogeneous in terms of size. This heterogeneity may dilute the potential in vivo application of LUVs because their effects could vary. To ensure reproducible effects, liposome vesicles of defined size and homogeneity should be prepared. After the LUV size was defined and homogenized by extrusion, the cationic REVs prepared in this study had a trapping volume of about 2.2 μL, including 2.02 nmol of 111In-labeled oligonucleotides. The trapping efficiency was 45%, and the transfection efficiency was 13.6%.
Antisense oligonucleotides generally break targeted RNA by activation of endogenous cellular nucleases, such as ribonuclease H or the nuclease associated with the RNA interference mechanism (33). Oligonucleotides that inhibit expression of the target gene by noncatalytic mechanisms, such as modulation of splicing or arrest of translation, can also be potent and selective modulators of gene function (33). Such an antisense mechanism, however, was not expected in this study. Antisense did play a role in conveying the Auger electrons emitted by 111In close to the target mRNA. Although 111In-phosphorothioate antisense could be broken apart in cell culture medium–supplemented serum, little free 111In was observed in this study.
DNA strand breaks may be caused by Auger electron emitters in close proximity to DNA in vitro (12–16). One study found that during the first 30 d after treatment, most breaks caused by 125I were located within 10 base pairs around the decay site and that they were due to the direct action of the radioisotope rather than diffusible free radicals (13). A hydroxyl radical scavenger, dimethyl sulfoxide (DMSO), protected against damage by free 125I in solution, but the minimal effect on the damage was noted even in the presence of DMSO by groove-bound 125I (14). Another study found that decay of 111In produced highly localized DNA breaks over 12 d, with efficiency nearly comparable with that of 125I (16). A study on the effects of 111In decay, using diethylenetriaminepentaacetic dianhydride to restrict the intracellular kinetics of 111In binding to DNA, demonstrated that the average 11-fold increased effectiveness of Auger electrons of 111In when in proximity to DNA appears to be due mainly to the higher yield of single-strand breaks (15). Decay of the Auger electron emitters causes DNA breaks in the target gene, leading—under certain conditions—to its inactivation (34–36). We could detect 111In-labeled antisense that hybridized with the target N-myc mRNA in tumor cells at 12 and 24 h after the cells were exposed to cationic REVs encapsulating the antisense, but this was not detected at 48 h. Reduction in N-myc expression, however, was observed at 48 h after treatment. These findings suggest that N-myc mRNA breaks were caused by Auger electrons of 111In in close proximity to the mRNA and that N-myc translation was then arrested. The effect of intracellular Auger electrons emitted by 111In has been demonstrated in mammalian cells in vitro (37). As little as 4.4 mBq per cell (about 4.4 MBq/L of culture) of intracellular 111In were able to affect the progression of cells through their first division cycle, whereas extracellular 111In at 1,150 MBq/mL of culture had little effect (37). About 7.5% of intracellular 111In was associated with DNA (37). On the other hand, in our study, the radioactivity of the 111In-labeled antisense integrated into the target of N-myc mRNA of a tumor cell at 24 h after the initiation of treatment was calculated to be about 556 mBq. Specific reduction in N-myc expression was caused by Auger electrons from 111In-labeled antisense. 111In-Labeled sense was incorporated into cells as much as the radiolabeled antisense, but it did not cause a significant biologic effect. These results emphasize that Auger electrons should be emitted in close proximity to their target N-myc mRNA.
111In and 125I have been investigated extensively as Auger electron emitters. The fact that 111In breaks DNA only half as effectively as 125I is in good agreement with the isotopes' calculated total energies and numbers of Auger electrons (16). The model of the mean absorbed dose-rate ratio of the tumor to normal tissue (TND)—a simple model that takes into account both the electron energy and the photon contribution in the evaluation of suitable radionuclides for therapy—demonstrates the importance of a low photon-to-electron energy ratio (p/e) in obtaining a high TND (17). The high abundance of photons associated with 111In (p/e ≈ 10) is predicted to reduce TND >5-fold compared with 125I (p/e ≈ 2) (17). However, 111In is still useful therapeutically because it is commercially and clinically available and has reliable radiochemical purity, particularly in vivo, compared with 125I.
Our cell proliferation study showed >96% cell viability during the observation period, and about 27% of the cell proliferation was suppressed 48 h after treatment with the radionuclide. In the tumorigenicity portion of the study, the tumor produced by the human neuroblastoma cells treated with cationic REVs encapsulating 111In-labeled antisense showed about 48% weight inhibition of the control. During the observation period, however, no significant differences in tumor growth rate were detected among cells treated with radiolabeled antisense, nonradiolabeled antisense, and control (empty) REVs. Cell proliferation was suppressed after N-myc mRNA was damaged by Auger electrons, but it recovered with the replenishment of N-myc mRNA because the N-myc gene itself remained intact. Thus, it can be speculated that treatment with cationic REVs encapsulating 111In-labeled antisense may prolong the time to tumor cell proliferation but does not kill the tumor cells. The duration of this effect may depend on the half-life of N-myc mRNA in the tumor cells. However, higher doses of radiolabeled antisense might cause more damage of the tumor cells. Prolongation of the time to cell proliferation might be applicable as a tumor dormancy therapy (38) by repeating the treatment. The intraperitoneal administration of cationic REVs encapsulating 111In-labeled antisense could be a potential approach to molecular therapy of small peritoneal metastases of neuroblastoma.
Many phosphorothioate oligodeoxynucleotides have been evaluated for their toxicologic effects in animals (39). In rodents, the most prominent toxic effects appear to be related to cytokine release induced by these agents. In monkeys, complement activation, abnormalities in clotting, and hypotensive events have been reported. However, no significant toxic effects have been observed in humans. The most probable mechanism of the observed toxic effects in laboratory animals is the binding of phosphorothioate oligodeoxynucleotides to protein (33). Different patterns of toxicity may exist between species (39). The technique used in this study, which involved use of a tiny dose of antisense labeled with 111In—too small to have a biologic effect on cells—and the encapsulation of antisense by cationic REVs to ensure delivery into the cells, might minimize the potential for toxicologic effects.
CONCLUSION
Auger electrons emitted from 111In reduced the proliferation of human neuroblastoma cells with overexpressed N-myc by inhibiting N-myc expression. Auger electron therapy, therefore, has potential as an internal radiotherapy targeting oncogenes at the level of the mRNA.
Acknowledgments
We thank Prof. Tadashi Yamamoto (Department of Oncology, Institute of Medical Science, The University of Tokyo, Tokyo, Japan) for his kind guidance in the technical aspects of molecular techniques.
Footnotes
-
COPYRIGHT © 2006 by the Society of Nuclear Medicine, Inc.
References
- Received for publication March 9, 2006.
- Accepted for publication July 25, 2006.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}