


Abstract
Tumors often have an increased uptake of glucose and can be detected by PET imaging using 18F-FDG. 18F-FDG is converted to 18F-FDG-6-phosphate (18F-FDG-6-P), and the usual assumption is that 18F-FDG-6-P is not a substrate for subsequent enzymatic reactions and that tumor hot spots reflect trapping of 18F-FDG-6-P. We recently found, however, that in the pig liver, 18F-FDG is metabolized not only to 18F-FDG-6-P but also to the subsequent oxygenation product 2-18F-fluoro-2-deoxy-6-phospho-d-glucononate (18F-FD-PG1). We therefore wished to characterize the metabolism of 18F-FDG in experimental tumors in mice. Methods: 18F-FDG was given intravenously to mice with either SCCVII squamous cell carcinoma or C3H mammary carcinoma grown on the back. 18F-Labeled metabolites were determined by radio-high-performance liquid chromatography in tumor tissue biopsies, in a time course of 180 min (12 mice of each tumor type), and in liver tissue biopsies 80 min after tracer injection (2 mice of each type). Results: After the tracer injection, not only 18F-FDG and 18F-FDG-6-P but also 18F-FD-PG1 and 2-18F-fluoro-2-deoxy-1,6-biphosphate were detected in both tumors, relatively more in SCCVII carcinoma than in C3H carcinoma. Both tumors accumulated radioactivity throughout the 180-min measurement period, 4-fold more in SCCVII carcinoma than in C3H carcinoma. At 80 min, the radioactivity was approximately 6 and 1.2 times higher in the respective tumors than in liver tissue. Conclusion: Our results agree with the general finding that most malignant tumor tissues accumulate significantly more 18F-radioactivity than do normal tissues, but our results do not support the concept that this increase is caused solely by accumulation of 18F-FDG-6-P. Furthermore, the rate of 18F-FDG metabolism was higher in SCCVII carcinoma than in C3H carcinoma.
Metabolic processes in intact organisms can be imaged and quantified using PET. Because glucose uptake is often increased in tumor tissue (1,2), tumors can be detected by PET through use of the glucose analog 18F-FDG as a radioactive tracer (3–5). 18F-FDG is transported across the cell membrane by the same carrier mechanism as that which transports glucose, and 18F-FDG is subsequently phosphorylated in parallel to glucose by hexokinase to 18F-FDG-6-phosphate (18F-FDG-6-P). The usual assumption is that 18F-FDG-6-P is not a substrate for subsequent enzymatic reactions and that intracellular trapping of 18F-FDG-6-P (6–9) is the reason for the radioactivity accumulation in tumor hot spots on PET images.
We recently found, however, that in pig liver, 18F-FDG not only is metabolized to 18F-FDG-6-P but also is significantly metabolized to the subsequent oxygenation products, 2-18F-fluoro-2-deoxy-6-phospho-d-gluconolactone (18F-FD-PGL) and 2-18F-fluoro-2-deoxy-6-phospho-d-gluconate (18F-FD-PG1) (10). These metabolites started to appear in liver tissue 45 min after the 18F-FDG injection, and their sum reached a level of 40% of the metabolite contents after 180 min. Dienel et al. (11), in studying rat brain, found that 1-14C-2-deoxy-d-glucose was converted not only to 1-14C-2-deoxy-d-glucose-1-phosphate but also to 1-14C-2-deoxy-d-glucose-1,6-biphosphate. To our knowledge, only 1 study (12) has attempted to characterize and quantify the metabolites of 18F-FDG in tumor tissue in vivo. High-performance liquid chromatography (HPLC) on Rous rat sarcoma found that 18F-FDG was metabolized to 18F-FDG-6-P and to several other 18F-labeled metabolites, identified as 18F-FDG nucleotide derivatives of 18F-FDG. Whether the assumption of trapping of 18F-FDG-6-P in tumor tissue is correct, and whether 18F-FDG metabolism is similar in different tumors, are thus open questions. If the metabolism is not similar, what are the consequences for the interpretation of 18F-FDG PET images, for kinetic modeling of 18F-FDG metabolism, and for tumor detection by 18F-FDG PET in oncology? To answer these questions therefore, we wished to conduct a preclinical study that would characterize and quantify 18F-FDG metabolism in experimental carcinomas grown on mice.
MATERIALS AND METHODS
Animals and Tumors
The Danish Animal Experimentation Inspectorate approved the study procedures. Twelve male C3H/KM mice (age range, 12–16 wk; body weight range, 27–41 g) with SCCVII squamous cell carcinoma grown on the back and 12 male CDF1 mice (age range, 12–16 wk; body weight range, 30–40 g) with C3H mammary carcinoma grown on the back were used. Derivation and maintenance was as described previously (13). Average tumor volumes were 790–1,180 mm3 (median, 990 mm3) for SCCVII carcinoma and 600–920 mm3 (median, 760 mm3) for C3H carcinoma. The mice were randomly allocated to the time points of the tumor biopsies.
Radiolabeled Tracer
18F-FDG was produced at the radiochemistry facility of the PET Center of Aarhus University Hospital (Aarhus, Denmark) as previously described (10). The radiochemical purity was >95%, and the specific radioactivity was approximately 72 GBq/μmol. The main impurities were partly hydrolyzed acetylated 18F-FDG (≤3%) and 18F-fluoride (≤1%).
Experimental Setup and Tumor Biopsies
The mice fasted for approximately 4 h. They were restrained in lucid plastic jigs with the tails secured with tape to allow an intravenous line to be inserted into the tail vein. The intravenous line consisted of a butterfly needle (25 gauge) and a polyethylene tube. The volume was approximately 70 μL and adjusted for at the tracer injection by flushing with isotonic saline with heparin. The 18F-FDG was given intravenously in a volume of only 100 μL to minimize changes in the animals’ blood volume and circulation. For the SCCVII carcinoma mice, the 18F-FDG dose was 3.1–5.9 MBq per animal (median, 4.4 MBq per animal). For the C3H carcinoma mice, the 18F-FDG dose was 2.0–2.7 MBq per animal (median, 2.4 MBq per animal). The mice were anesthetized with 0.40 mL intravenous mebumal (4.76 mg/mL saline) 10 min before the biopsy samples were taken. Biopsies were performed using the through-cut technique (cutting biopsy needle, 2.1-mm diameter; Worldwide Medical Technologies, Woodbury, CT). From 2 animals (for double determinations) of each tumor type, tumor biopsy samples were taken 5, 20, 45, 80, 110, and 180 min after the 18F-FDG injection and liver biopsy samples were taken at 80 min. Because of the small size of tumors and livers in mice, 3–4 biopsies, each of 6–8 mg, were taken at each time point from each mouse. The animals were killed by cervical dislocation. The tissue samples were immediately transferred into a precooled (liquid nitrogen) custom-made Teflon (DuPont, Wilmington, DE) container. For homogenization, the container held a tungsten ball and was transferred into a mill (mixer mill MM 200; Retsch Gmbh and Co., KG, Haan, Germany), in which it was shaken vigorously for 60 s to disintegrate the frozen tissue samples into subcellular fractions (14). After 10 min at room temperature, the samples were thawed and mixed with 1,000 μL phosphate buffer (0.01 mol/L K2HPO4/KH2PO4, 1.5 mmol/L K2-ethylenediaminetetraacetic acid disodium salt dihydrate [EDTA], 0.01 mol/L monothioglycinol, 10% glycinol [v/v], 0.01 mol/L Na2MoO4* 2H2O, pH 7.5) by being shaken in the mill for 60 s. The homogenate was divided into 3 samples: 400 μL homogenate were transferred into a V-shaped plastic vial containing 500 μL acetonitrile for protein precipitation, 100 μL were transferred into a vial used for counting radioactivity in a well scintillation counter (Cobra; Canberra Packard, Zurich, Switzerland), and the rest was transferred into a V-shaped plastic vial and stored at −80°C for 1 d, until the protein concentration was measured using the method of Lowry et al. (15). The sample containing acetonitrile was centrifuged (Micromax centrifuge; IEC International Equipment Company, Needham Heights, MA) for 2 min at 13,000 rpm to precipitate the proteins. The supernatant was transferred into a plastic tube, and acetonitrile was evaporated at 20 mm Hg. Finally, the solution was centrifuged for 30 s at 13,000 rpm, and the supernatant was analyzed by radio-HPLC.
In Vitro Enzymatic Preparation of 18F-FDG-6-P, 18F-FD-PGL/18F-FD-PG-1, and 2-18F-Fluoro-2-Deoxy-1,6-Biphosphate (18F-FDG-1,6-P2)
18F-FDG-6-P (12)
Ten milligrams (250–400 units) hexokinase type IV from bakers’ yeast (EC 2.7.1.1; Sigma-Aldrich A/S Denmark, Vallensbaek Strand, Denmark) were dissolved in 1 mL 70 mmol/L NaH2PO4 buffer. Two milligrams (3.6 μmol) adenosine triphosphate (Sigma-Aldrich A/S Denmark) and 1 mg (5 μmol) MgCl2*6H2O (Sigma-Aldrich A/S Denmark) were added. The pH was adjusted to 7.4 using diluted sodium hydroxide solution. Approximately 100 MBq 18F-FDG were added, and the solution was stirred at room temperature (20°C). Samples were taken, and phosphorylation was followed and analyzed by radio-HPLC. The formation of 18F-FDG-6-P was confirmed by coinjection with nonradioactive 19F-FDG-6-P (Sigma-Aldrich A/S Denmark).
18F-FD-PGL/18F-FD-PG-1 (12,16,17)
These products were prepared by enzymatic oxidation of 18F-FDG-6-P. 18F-FDG-6-P was synthesized, and after completion of phosphorylation, 5 mg (6.5 μmol) β-nicotinamide adenine dinucleotide phosphate (β-NADP) (Sigma-Aldrich A/S Denmark) and 35 units glucose-6-phosphate dehydrogenase (EC 1.1.1.49; Boehringer-Mannheim, Mannheim, Germany) were added. After the pH was readjusted with diluted sodium hydroxide solution to 7.2, the solution was stirred at room temperature (20°C). Samples were taken to follow the enzymatic oxidation and were analyzed by radio-HPLC. Because the procedure has been well described in the literature (12,16,17) and enzymatic processes are highly specific, the product was not further characterized.
18F-FDG-1,6-P2 (11,18)
This product was synthesized enzymatically by conversion of 18FDG-6-P using phosphoglucomutase in the presence of glucose-1,6-diphosphate. 18FDG-6-P was synthesized, and after completion of phosphorylation, 1.1 mg (2.7 μmol) EDTA (Sigma-Aldrich A/S Denmark), 1.3 mg bovine serum albumin (Sigma-Aldrich A/S Denmark), 1.1 mg (1.5 μmol) α-d-glucose-1,6-diphosphate cyclohexylammonium salt (Sigma-Aldrich A/S Denmark), and 20 units phosphoglucomutase (EC.5.4.2.2; Boehringer-Mannheim) from rabbit muscle were added. The mixture was stirred at room temperature (20°C) after the pH was adjusted to 7.9 with diluted sodium hydroxide solution. Samples were taken to follow the enzymatic phosphorylation and were analyzed by radio-HPLC. Because the procedure has been well described in the literature (11,18) and enzymatic processes are highly specific, the product was not further characterized.
Radio-HPLC
Radio-HPLC analysis of the tumor and liver tissue samples was performed on Perkin-Elmer Denmark A/S equipment (Allerød, Denmark) as described previously (10), the only difference being that the pH of the mobile phase (20 mmol/L potassium phosphate buffer) was 6.6 instead of 6.9 to adjust for the relatively high pH of the phosphate buffer. The chromatograms were analyzed using commercially available radiochromatography software (Winnie 3.2; raytest Isotopenmessgeraete GmbH, Straubenhardt, Germany). Radio-HPLC identification of the HPLC findings was done by analysis of products from in vitro enzymatic reactions (12). For radio-HPLC of the biopsy samples, an isocratic LC pump 250 (Perkin-Elmer Denmark A/S) equipped with a Rheodyne 7125 injector (Perkin-Elmer Denmark A/S) and 1-mL stainless steel injection loop connected in series with a SphereClone 5-μm strong-anion-exchange (SAX) 250 × 4.6 mm column (Phenomenex, Torrance, CA), an LC 295 UV/Vis detector (Perkin-Elmer Denmark A/S), and a sodium iodide scintillation detector (GABI; raytest) was used. For biopsy samples, only the radioactivity channel was recorded. Chromatograms were analyzed using the Winnie 3.2 software. The mobile phase was 20 mmol/L potassium phosphate buffer, pH 6.6, and the flow rate was 0.8 mL/min. The pH was lowered from the previously reported 6.9 (10) to 6.6 to adjust for the relatively high pH of the phosphate buffer used for tissue extraction. Solutions from the in vitro phosphorylation experiments were analyzed using a binary LC pump 250 equipped with a Rheodyne 7125 injector and a 100-μL stainless steel injection loop connected in series with a SphereClone 5-μm SAX 250 × 4.6 mm anion exchange column, a series 200 refractive index detector (Perkin-Elmer Denmark A/S), and a sodium iodide scintillation detector of in-house design. Chromatograms were analyzed using commercially available chromatography software (Turbochrom 4.1.2; Perkin-Elmer Denmark A/S). The mobile phase was 20 mmol/L potassium phosphate buffer, pH 7.2, and the flow rate was 0.8 mL/min.
Efficiency of Tissue Sample Preparation and Radio-HPLC
For both tumors and liver tissue samples, 95%–98% of the radioactivity in the tissue samples was found in the tissue extracts. Recovery of the injected radioactivity from the HPLC column was 85%–110%, compared with an aliquot sample of standard. The main source of error was the volume determination for aliquot samples.
RESULTS
In Vitro Phosphorylation
18F-FDG-6-P
18F-FDG showed the known high affinity toward hexokinase and was efficiently phosphorylated under the chosen conditions. Ten minutes after the addition of 18F-FDG to the hexokinase/adenosine triphosphate/MgCl2 mixture, >90% of radioactivity was in the form of 18F-FDG-6-P. Remarkably, phosphorylation was also observed in the absence of MgCl2, but with a much slower reaction rate (90% 18F-FDG-6-P 30 min after the addition of 18F-FDG). Obviously, the normal magnesium content of the used glass vials was enough to catalyze the enzymatic phosphorylation. No other radioactive compounds besides 18F-FDG and 18F-FDG-6-P were observed, confirming the specificity toward phosphorylation by hexokinase.
18F-FD-PGL/18F-FD-PG-1
Five minutes after the addition of β-NADP and glucose-6-phosphate dehydrogenase, formation of the oxidation product was already observed. Compared with phosphorylation, enzymatic oxidation occurred at a slower reaction rate. After 180 min, 80% of the analyzed radioactivity was in the form of 18F-FD-PGL/18F-FD-PG-1. No other radioactive products besides 18F-FDG, 18F-FDG-6-P, and 18F-FD-PGL/18F-FD-PG-1 were observed, confirming the specificity of the enzymatic reaction. Gluconolactones are unstable with respect to hydrolysis, and ring opening occurs with a half-life of 90 s at pH values > 6.0 (19,20). Thus, the initially formed 18F-FD-PGL hydrolyzed under the given pH conditions of the enzymatic oxidation (pH 7.2) and HPLC separation (pH 7.2), and the product detected was 18F-FD-PG-1.
18F-FDG-1,6-P2
As reported previously (10,18), formation of the 1,6-diphosphate was observed only in the presence of α-d-glucose-1,6-diphosphate. In the presence of α-d-glucose-1,6-diphosphate as cofactor, 18F-FDG-1,6-P2 formed. Three minutes after the start of the enzymatic reaction, 30% of the analyzed radioactivity was already in the form of 18F-FDG-1,6-P2. After 22 min, the diphosphate was the main product (83%). No other radioactive products besides 18F-FDG, 18F-FDG-6-P, and 18F-FDG-1,6-P2 were observed, confirming the specificity of the enzymatic reaction.
Radio-HPLC
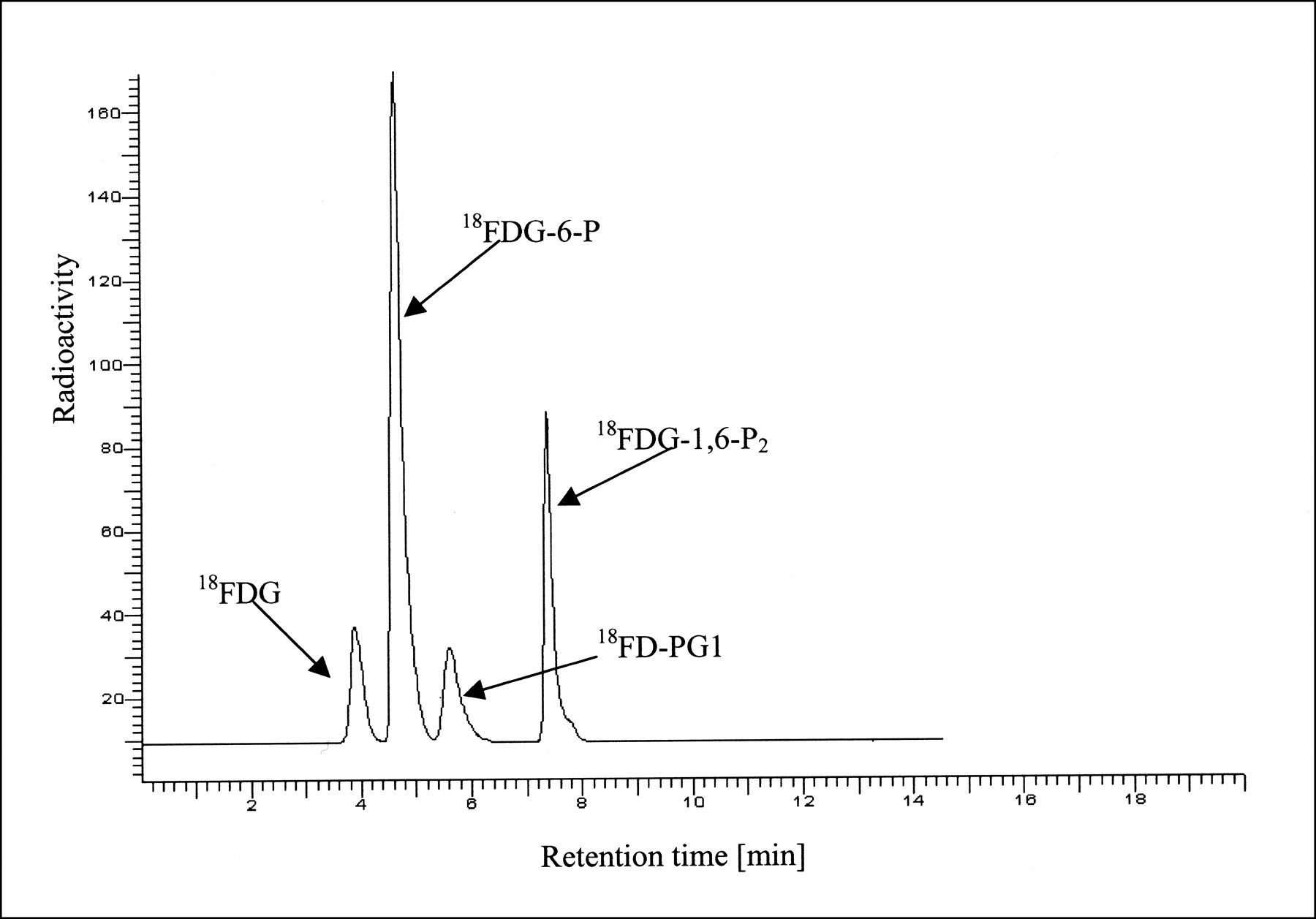
The radio-HPLC method allowed efficient separation of 18F-FDG, 18F-FDG-6-P, 18F-FD-PGL/18F-FD-PG-1, and 18F-FDG-1,6-P2. Because of the findings of 19F nuclear magnetic resonance experiments, 2-18F-fluoro-2-deoxy-mannose (FDM) and 18F-FDG-1-P are possible metabolites. These compounds are present in early stages of the metabolism and are therefore of particular importance. However, separation of these compounds from 18F-FDG and 18F-FDG-6-P is not possible under the given HPLC conditions. 18F-FDM coelutes with 18F-FDG, as confirmed by coinjection of nonradioactive FDG and FDM (reference compounds were purchased from Sigma-Aldrich A/S Denmark). As a test for the separation of 18F-FDG-1-P from 18F-FDG-6-P, a mixture of glucose-1-phosphate and glucose-6-phosphate (reference compounds were purchased from Sigma-Aldrich A/S Denmark) was analyzed under the given conditions. Because separation of glucose-1-phosphate from glucose-6-phosphate was not observed, separation of 18F-FDG-1-P from 18F-FDG-6-P is unlikely. Figure 1 shows the radio-HPLC findings for a mixture of 18F-FDG-6-P, 18F-FD-PGL/18F-FD-PG-1, and 18F-FDG-1,6-P2 obtained by the enzymatic in vitro reaction described above. 18F-FD-PGL/18F-FD-PG-1 and 18F-FDG-1,6-P2 were prepared separately from 18F-FDG-6-P and mixed afterward.

Analytic radio-HPLC of mixture of 18F-FDG, 18F-FDG-6-P, 18F-FD-PG1, and 18F-FDG-1,6-P2. Compounds were prepared individually by in vitro enzymatic reactions and mixed afterward (SphereClone [Phenomenex, Torrance, CA] 5-μm SAX 250 × 4.6 mm column; 20 mmol/L potassium phosphate buffer; pH 7.2; flow rate, 0.8 mL/min).
18F-FDG Purity
The purity of the 18F-FDG used in both in vitro and in vivo experiments was >95%. The main impurities caused by the applied synthetic process were partly hydrolyzed, acetylated 18F-FDG (≤3%) and 18F-fluoride (≤1%). Not being an anion, partly hydrolyzed and acetylated 18F-FDG elutes with 18F-FDG. Furthermore, under the given chromatographic conditions, 18F-fluoride is not separated from 18F-FDG. On injection of the used 18F-FDG, no radioactivity eluted after the 18F-FDG. Thus, the radioactive fractions eluting after 18F-FDG and 18F-FDG-6-P cannot be attributed to impurities arising from the used 18F-FDG formulation and were therefore endogenous products.
18F-FDG Metabolites in SCCVII Carcinoma
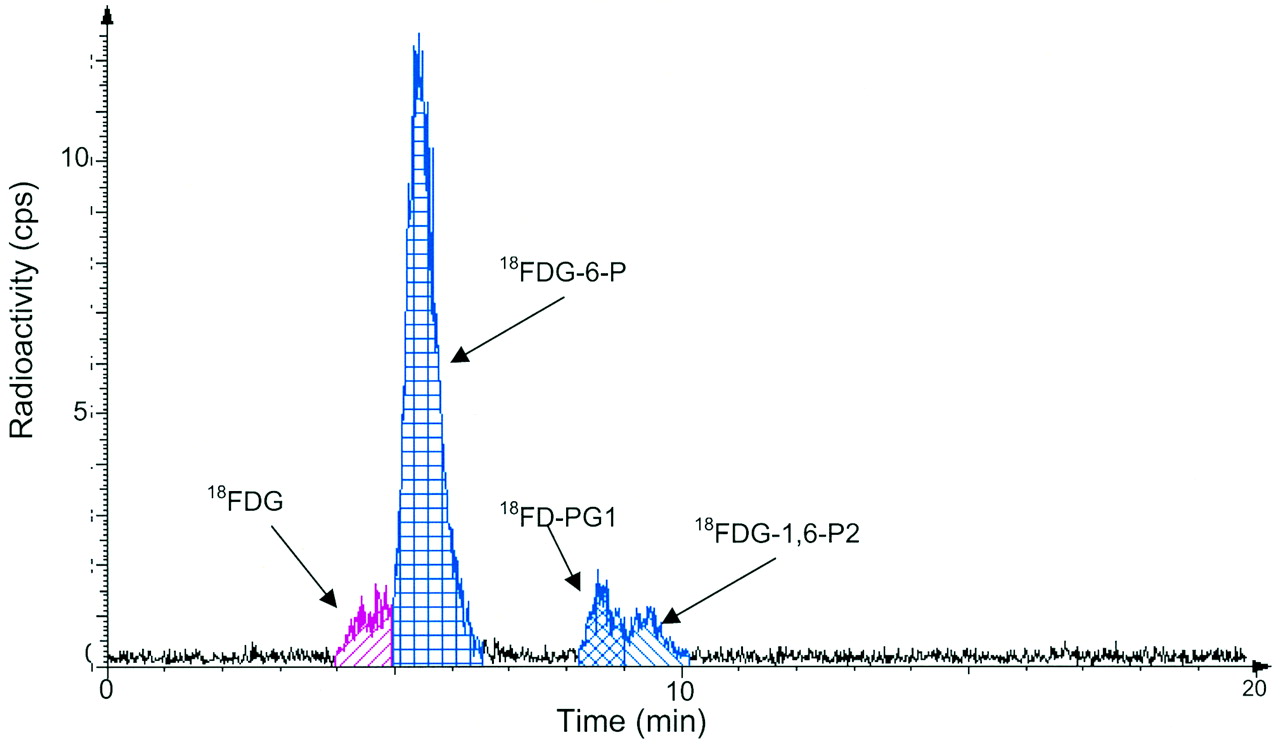
After the 18F-FDG injection, not only 18F-FDG and 18F-FDG-6-P but also 18F-FD-PG1 and 18F-FDG-1,6-P2 were detected in the SCCVII carcinoma tissue samples. An example of the HPLC measurements of the tissue extract is shown in Figure 2. Figure 3 shows the time course of the tissue content of 18F-FDG and the 3 metabolites. 18F-FDG decreased rapidly within the first 20 min and then decreased slowly for the rest of the 180-min measurement period. 18F-FDG-6-P appeared early and accounted for half of the radioactivity after 5 min and three fourths after 20 min. Afterward, it remained at that level. 18F-FD-PG1 had already been seen after 5 min and increased steadily until reaching a level of 15% at 180 min. A third metabolite, 18F-FDG-1,6-P2, appeared after 20 min and increased to 10% after 45 min, followed by a slow decrease throughout the measurement period. The content of other possible metabolites was <0.5% throughout the measurement period. No significant radioactivity was found in the acetonitrile precipitates (which would have contained 18F-labeled glycogen).
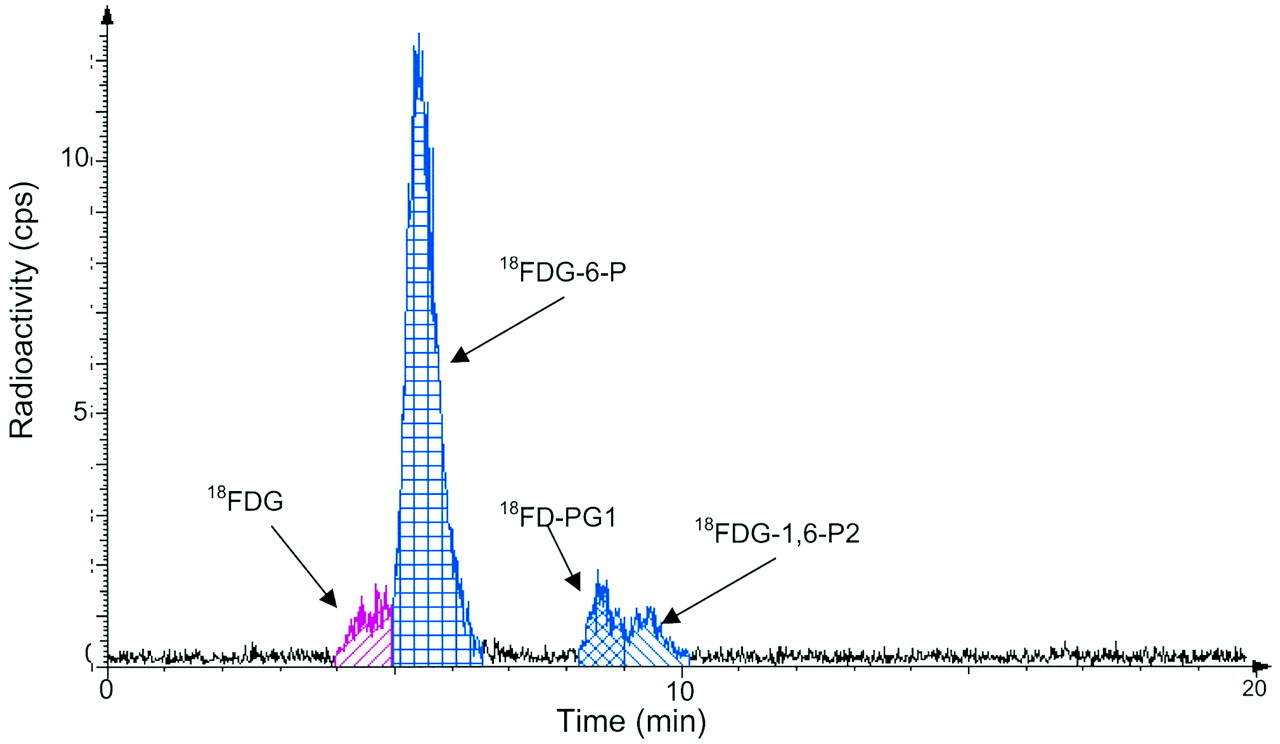
Typical radio-HPLC results for analysis of 18F-FDG in squamous tumor tissue extracts 80 min after 18F-FDG injection.
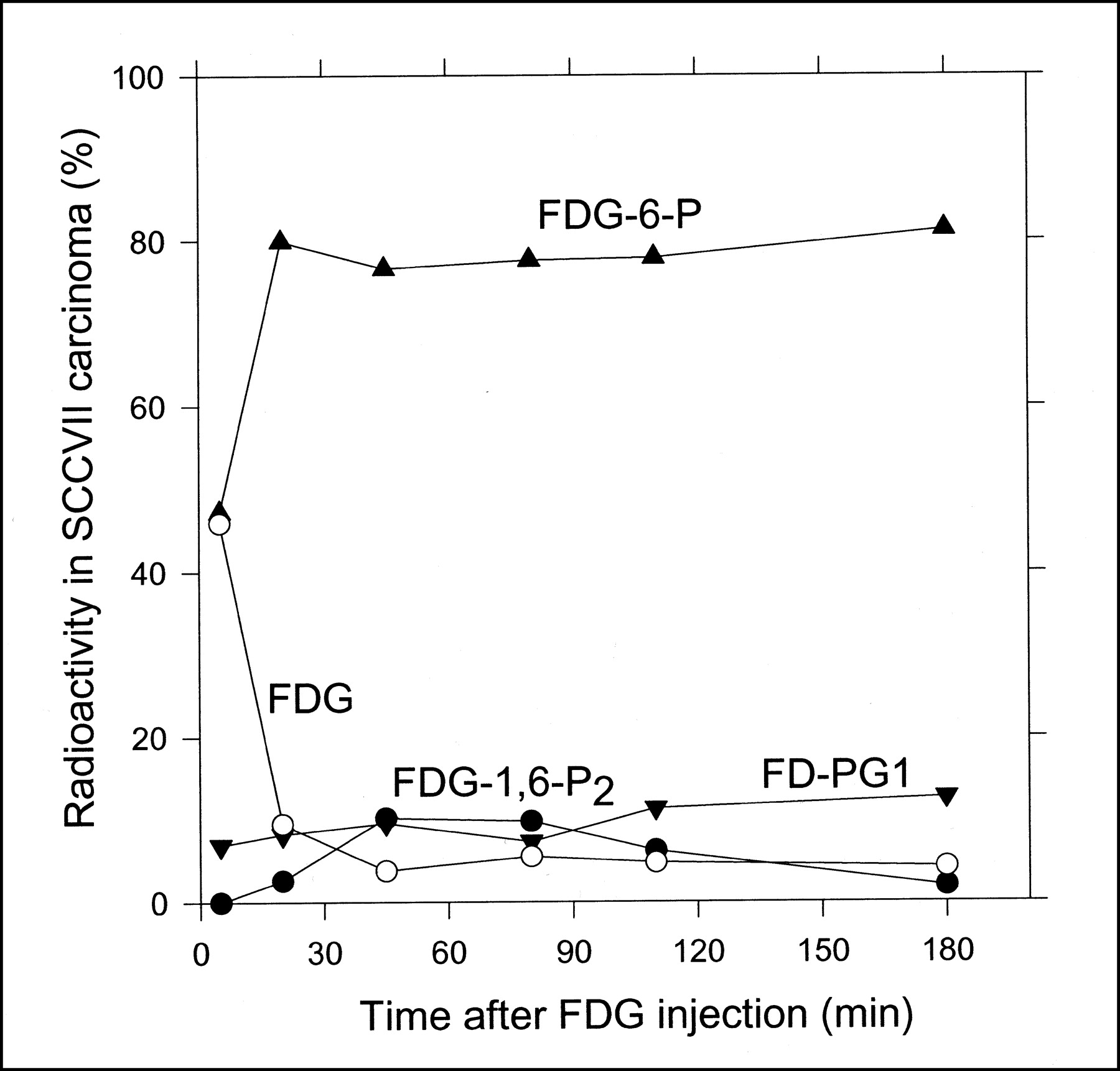
Radio-HPLC-determined time course of percentage tissue content of 18F-FDG (○), 18F-FDG-6-P(▴), 18F-FD-6-PG1 (+18F-FD-6-PGL) (▾), and 18F-FDG-1,6-P2 (•) in SCCVII carcinoma grown on mice.
18F-FDG Metabolites in C3H Carcinoma
Figure 4 shows the time course of the content of 18F-FDG, 18F-FDG-6-P, and 18F-FD-PG1 + 18F-FDG-1,6-P2 in C3H carcinoma tissue after the 18F-FDG injection. In Figure 4, the last 2 metabolites are shown as a sum because of their low content in tissue extracts. The time course of 18F-FDG and the metabolites in C3H carcinoma was similar to that in SCCVII carcinoma (Fig. 3), except for the significantly lower content of 18F-FD-PG1 and 18F-FDG-1,6-P2. The content of other possible metabolites was <0.5% throughout the measurement period. No significant accumulation of radioactivity was found in the acetonitrile precipitates.
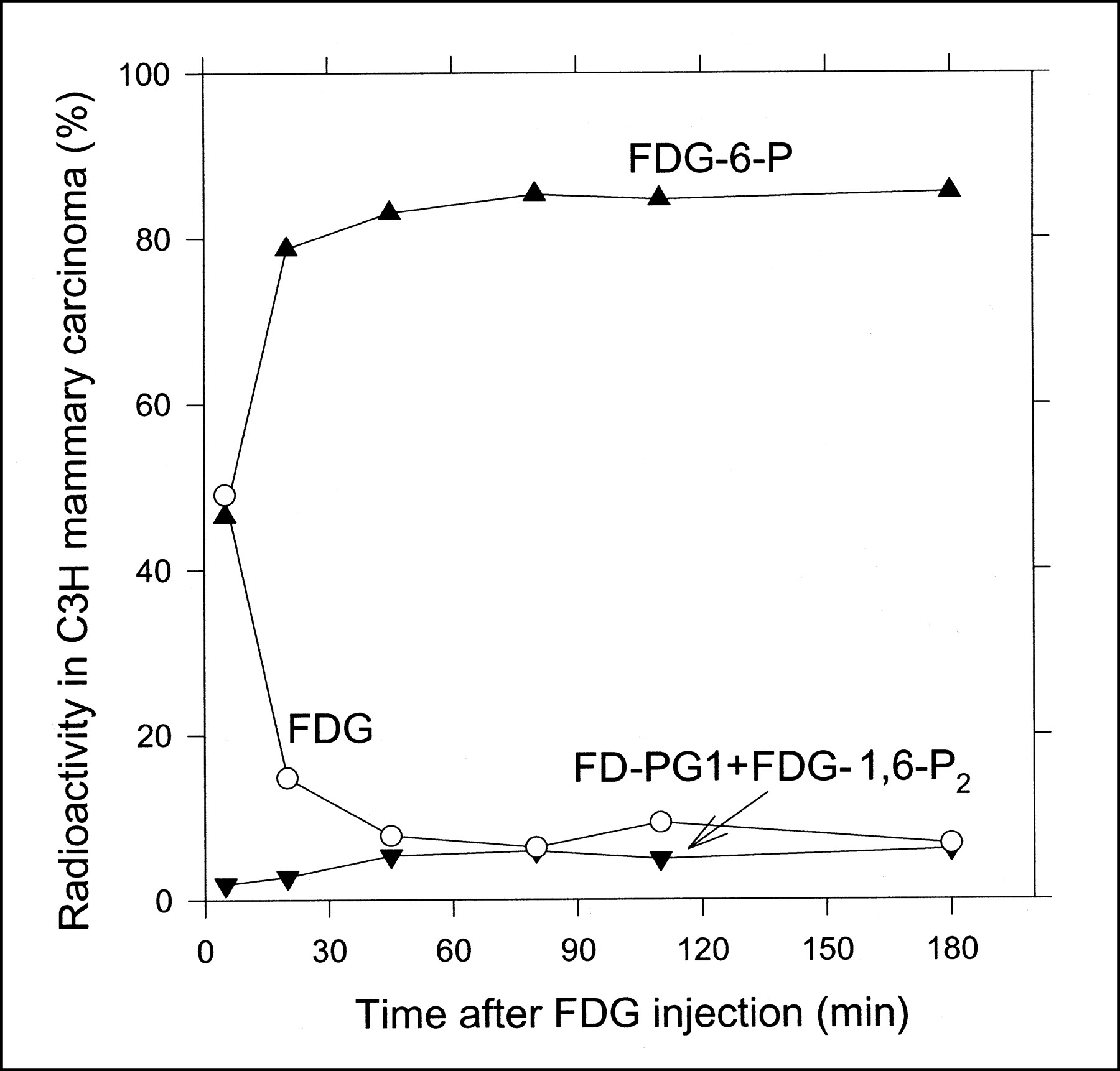
Radio-HPLC-determined time course of percentage tissue content of 18F-FDG (○), 18F-FDG-6-P(▴), and 18F-FD-PG1(+18F-FD-6-PGL) + 18F-FDG-1,6-P2 (▾) in C3H carcinoma grown on mice.
18F-FDG Metabolites in Liver
The liver content of 18F-FDG and metabolites at 80 min was similar in mice bearing either carcinoma, being on average 40% for 18F-FDG, 55% for 18F-FDG-6-P, and 7% for 18F-FD-PG1. No 18F-FDG-1,6-P2 was detectable.
Radioactivity Accumulation in Tumors and Liver
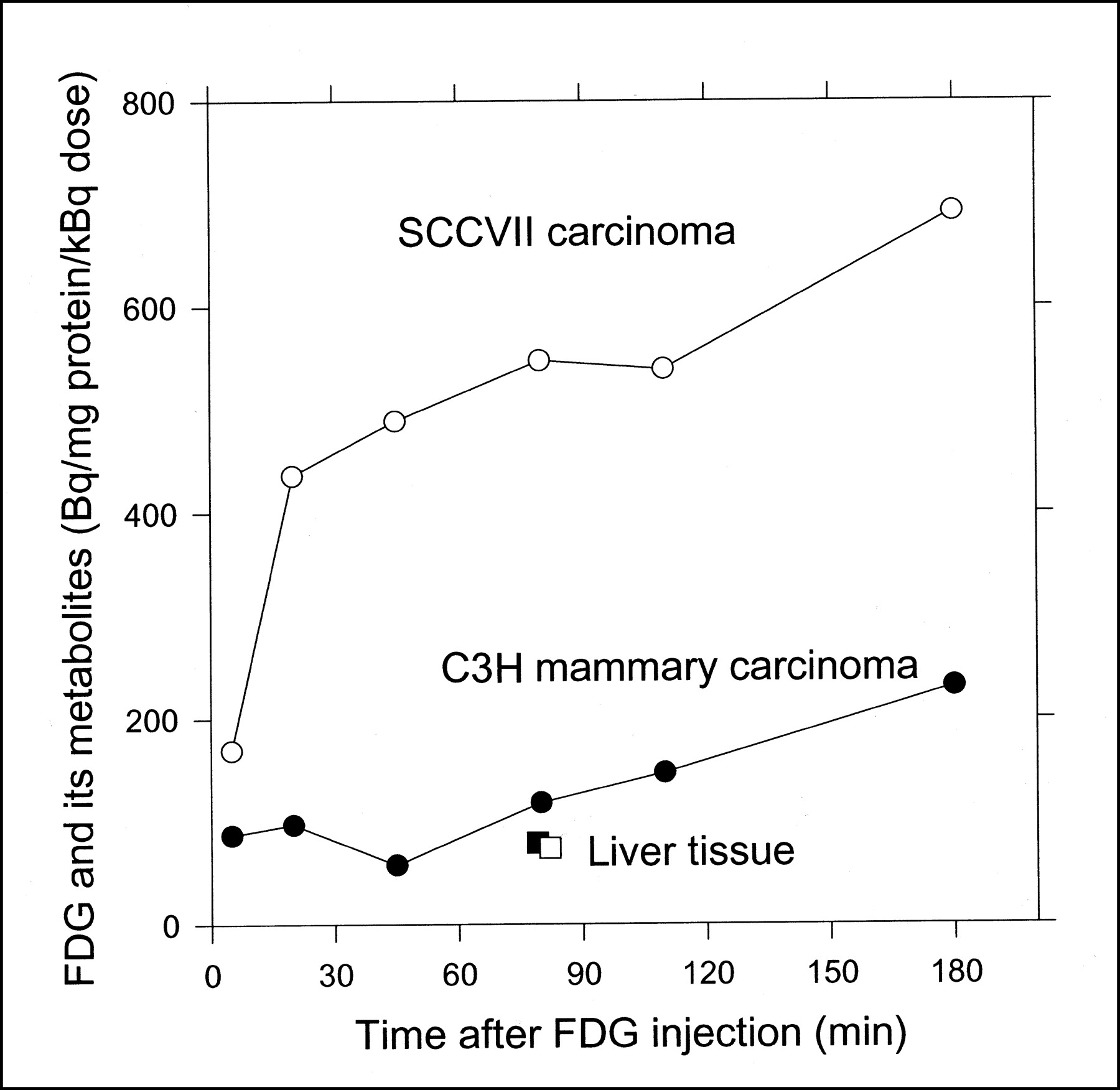
The absolute radioactivity concentrations of 18F-FDG + 18F-metabolites were calculated from the radioactivity concentrations of the tissue homogenates (corrected for radioactivity decay, with a half-life of 109 min for 18F) and the protein concentrations of the homogenates. Figure 5 shows the results normalized for individual 18F-FDG doses. The data were not normalized for body weights or tumor volumes because the ranges of these parameters were narrow. Both tumor types continued to accumulate radioactivity throughout the 180-min measurement period, and SCCVII carcinoma accumulated 4-fold more radioactivity than did C3H carcinoma (normalized for differences in the tracer doses). At 80 min, the radioactivity was approximately 6 and 1.2 times higher in the respective tumors than in the liver tissue.
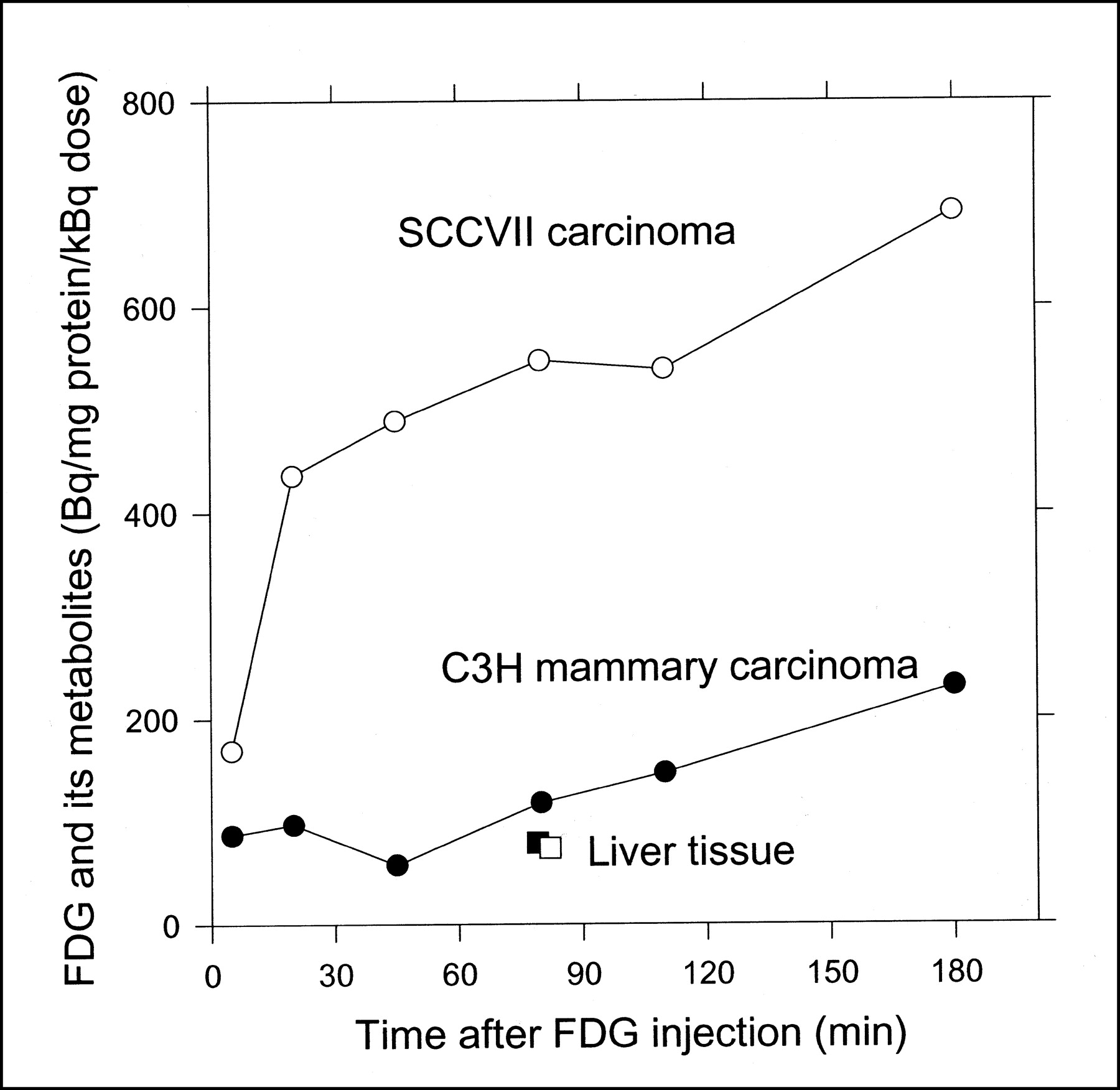
Time course of total radioactivity concentrations (18F-FDG + 18F-metabolites) in SCCVII carcinoma (○), C3H carcinoma (•), and liver tissue from respective mice (□, ▪). Concentrations are normalized for tissue protein concentrations and individual 18F-FDG doses.
DISCUSSION
This study showed, for the first time, the formation of 2 metabolites, 18F-FD-PG1 and 18F-FDG-1,6-P2, besides 18F-FDG-6-P in malignant tumor tissue in vivo after 18F-FDG administration. All 3 metabolites were found in both SCCVII carcinoma and C3H carcinoma. Figure 6 illustrates the possible metabolic pathways for 18F-FDG-6-P. The findings for 18F-FD-PG1 in the tumors showed that 18F-FDG-6-P was oxidized by tumor glucose-6-phosphate dehydrogenase to 18F-FD-PGL, which was further converted to 18F-FD-PG1 by hydrolysis. The oxidation of 18F-FDG-6-P is reversible (21), whereas the hydrolysis is considered irreversible. As we noted in a previous study on pig liver (10), 18F-FD-6-PGL might be an unstable product and might have been converted to 18F-FD-6-PG1 during tissue sample extraction. The measured 18F-FD-6-PG1 therefore also included 18F-FD-6-PGL. In addition, the presence of 18F-FDG-1,6-P2 in tumor tissues showed that 18F-FDG-6-P was converted to 18F-FDG-1-P by the phosphoglucomutase. This finding agreed with findings for 1-14C-2-deoxy-d-glucose (11) and 19F-FDG in rat brain (22,23). The chromatographic conditions of the HPLC method did not allow for separation of 18F-FDG-6-P from 18F-FDG-1-P, and the figures given for 18F-FDG-6-P thus might include some 18F-FDG-1-P. Finally, we did not identify any of the nucleotide derivatives of 18F-FDG-1-P or incorporation of 18F-FDG into glycogen, as has been discussed for 1-14C-2-deoxy-d-glucose (24–27). This finding was in accordance with the results of our previous biopsy experiments on pig liver (10).
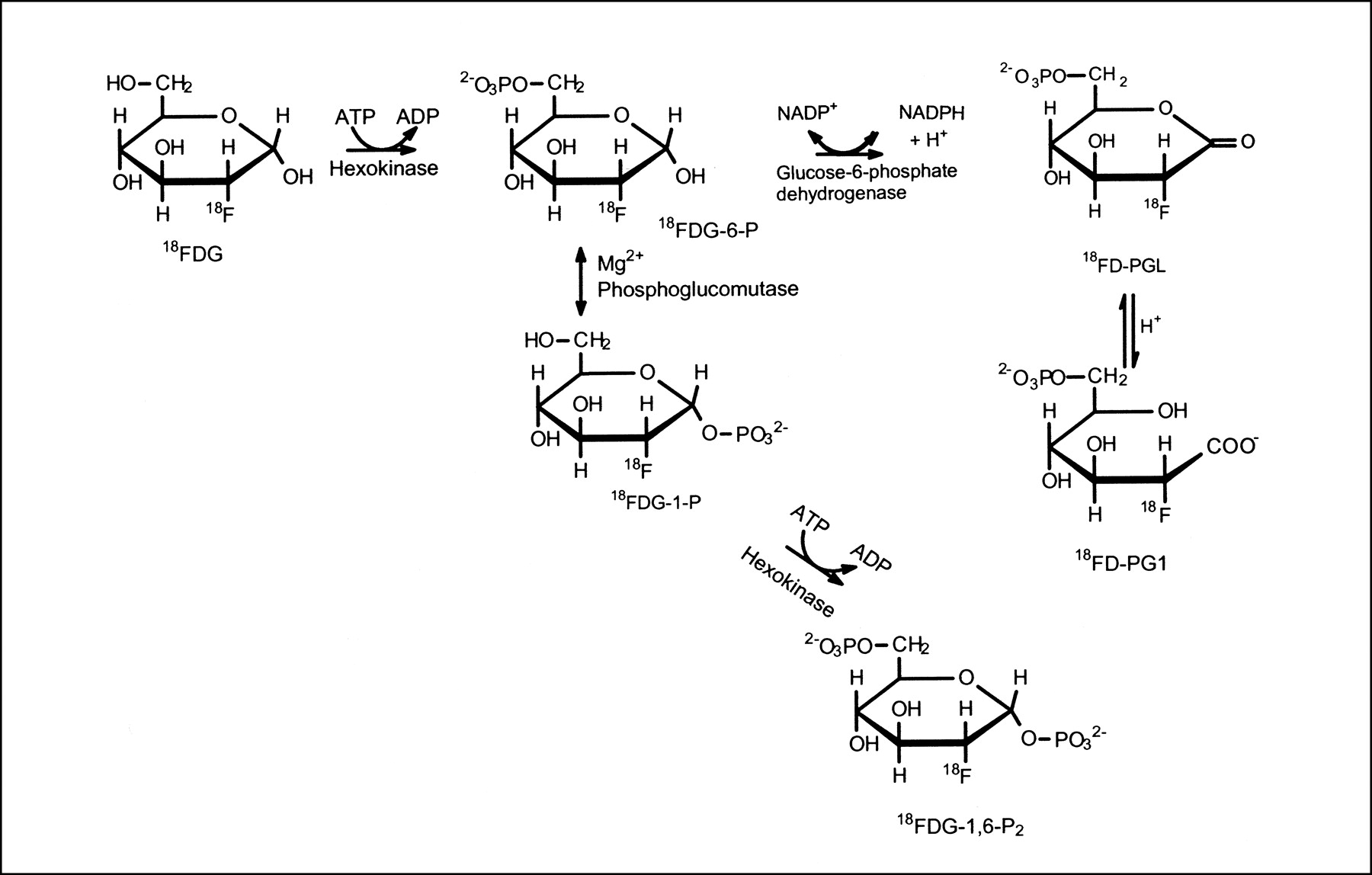
Metabolic pathways for 18F-FDG.
The levels of the secondary metabolites, 18F-FD-PG1 and 18F-FDG-1,6-P2, reached different concentrations in the 2 tumor types, 20% in the SCCVII carcinoma and 8% in the C3H carcinoma (Figs. 3 and 4). In accordance with this finding were the findings of Som et al. (28), that 18F-FDG uptake in tumors depended on both tumor type and animal species, and of Suolinna et al. (12), that only 3% of tumor radioactivity after 18F-FDG injection stemmed from 18F-metabolites other than 18F-FDG-6-P. Accumulation of radioactivity was 4-fold larger in SCCVII carcinoma than in C3H carcinoma (Fig. 5), possibly because of a larger relative number of viable malignant cells in SCCVII carcinoma as judged from the relative content of necrotic tissues (approximately 1% and 30% in SCCVII carcinoma and C3H carcinoma, respectively; M. Horsman, unpublished data, February 2002).
The accumulation of total radioactivity was lower in liver tissue than in either tumor type after 80 min (Fig. 5). Furthermore, the relative amount of 18F-FDG-6-P in liver tissue was low in both mice (50%) and pigs (30% (10)), compared with the high levels seen in tumor tissue (80%, Figs. 3 and 4). In accordance with these findings, Suolinna et al. (12) found that 45 min after 18F-FDG administration, 90% of the 18F-radioactivity in tumor tissue was 18F-FDG-6-P, versus 30% in liver tissue. The low level of 18F-FDG-6-P in liver tissue was probably caused by the reversible transport of 18F-FDG across the liver cell membrane and the degradation of 18F-FDG-6-P in liver tissue (5,29), neither of which occurs in tumor tissue.
The fact that 18F-FDG-metabolites other than 18F-FDG-6-P were found in tumor tissue may have implications for the interpretation of estimated kinetic rate constants in terms of the enzymatic processes as discussed for pig liver (10). However, the current data, with relatively few measurements, do not allow for quantitative evaluations for the rate constants. The finding does not in any way alter the conventional use of 18F-FDG uptake in tumor tissue as an indicator of the rate of glucose use, because the trapped label in tissue still reflects the 18F-FDG phosphorylation rate regardless of whether the label is associated with 18F-FDG-6-P or with the products of 18F-FDG-6-P. It would have been a completely different story if the newly discovered metabolites were products of 18F-FDG directly, that is, in parallel or in competition with the phosphorylation reaction.
CONCLUSION
Our results agree with the general finding that most malignant tumor tissues accumulate significantly more 18F-radioactivity than do normal tissues, but our results do not support the concept that this increase is caused solely by accumulation of 18F-FDG-6-P. These findings raise the question of whether differences in radioactivity accumulation in human tumors after 18F-FDG injection could be caused by differences in 18F-FDG metabolism. It would be interesting to study these phenomena in different human tumors in vivo.
Acknowledgments
This study was supported by grants from the Danish Cancer Society and the Danish Research Agency (Freja Program).
Footnotes
Received Oct. 15, 2001; revision accepted Mar. 25, 2002.
For correspondence or reprints contact: Susanne Keiding, DSc, PET Center, Aarhus University Hospital, DK 8000 Aarhus, Denmark.
E-mail: susanne{at}pet.auh.dk




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}