Article Text

Statistics from Altmetric.com
Preface
Continuous lymphocyte migration to normal gut is a prerequisite for immune homoeostasis in humans. In this review we briefly describe the physiology of lymphocyte recirculation through the bowel. The adhesion molecules mediating the lymphocyte–endothelial interactions in the gut during the multistep extravasation cascade will be presented in the light of their ability to confer mucosal selectivity of lymphocyte trafficking. We will also discuss the relevance of leucocyte recirculation in respect to bowel inflammation and mucosal vaccination, and its potential in the anti-inflammatory treatment of gastrointestinal diseases.
Lymphocyte recirculation—the general theme
The gut is the main portal of antigen entry into the body and lymphocytes are responsible for mounting an adequate immune response against harmful antigens. As each lymphocyte only carries an antigen receptor for a single antigen, a huge number of lymphocytes, each specific for a different antigen, are produced in the bone marrow and thymus each day. These naive cells must then be able to sample freely all different tissues of the body in search of their cognate antigens. To maximise the likelyhood of the rare possibility that a given lymphocyte would find its cognate antigen introduced anywhere in the body, a sophisticated system of lymphocyte recirculation has evolved.1-4
In this process, lymphocytes continuously patrol between the blood and different tissues of the body. Initially blood borne lymphocytes leave the circulation via secondary lymphoid tissues like lymph nodes. The exit from the blood mainly takes place in distinct postcapillary vessels called high endothelial venules (HEV), which display several unique structural and functional modifications to facilitate extravasation.5 Foreign antigens are retrieved and concentrated from distal epithelial surfaces via the afferent lymphatic system into the secondary lymphoid organs.
When an extravasated lymphocyte finds its antigen in the supportive context of the secondary lymphoid tissues, it starts to proliferate and differentiate and gives rise to numerous effector cells. These progeny cells then leave the lymphoid tissues via efferent lymphatics and re-enter the systemic circulation. During this process the activated effector lymphocytes have become imprinted in a way which allows them to migrate preferentially back (home) to the sites where they became activated (the reactive secondary lymphoid organ) and to the sites from which the inciting antigen was originally drained (the challenged epithelial surfaces). At these target organs the antigen specific effector cells then effectively clear the antigenic threat. After the acute inflammation has resolved, some of these cells revert into a less activated stage and survive for years as memory cells. These cells patrol at low levels the epithelial organs all the time and afford a brisk, antigen specific secondary immune response should the same antigenic challenge recur.
Lymphocyte migration into the gut
Intestinal antigens are sampled from the gut lumen by M cells which are positioned between the enterocytes. The antigens are translocated into underlying Peyer’s patches (the organised secondary lymphoid organs in the gut wall) where they can be efficiently processed and introduced to lymphocytes.6 Naive lymphocytes carried by the blood enter the Peyer’s patches by binding to the specialised HEV. If the lymphocyte does not find its antigen it leaves the Peyer’s patch and returns into the systemic circulation via the efferent lymphatics. If, however, the B or T lymphocyte recognises its cognate antigen it starts to divide and differentiate. After some time the immunoblasts are transported via the lymphatics into mesenteric lymph nodes. Ultimately, the effector cells return into the blood and are randomly dispersed all over the body. When those progeny cells sooner or later re-enter the bowel vasculature they can return to the Peyer’s patch via HEV. More importantly, the immunoblasts now can very efficiently bind to flat-walled venules of lamina propria and immigrate into this non-organised subepithelial lymphoid tissue of the gut, which is constantly subjected to immunostimulation. The different nature of these two lymphoid compartments is also highlighted by the fact that the molecular mechanisms directing lymphocyte migration into Peyer’s patches and lamina propria are somewhat distinct although overlapping (see later).7
The mucosal recirculation system opposes to that of peripheral lymph nodes (PLN) which drains antigens mainly from the skin. The whole concept of tissue selective lymphocyte homing was, in fact, worked out in the 1960s and 1970s by following the fate of transferred mucosal immunoblasts.8-11 In those in vivo animal experiments, gut derived immunoblasts showed 5–20-fold better localisation back to bowel than to PLN. It is traditionally thought that oral, intestinal, respiratory, mammary, and urogenital surfaces form a common mucosal lymphatic system which also constitutes a functionally unified recirculation system.12 More recently this view has been challenged.13 Furthermore, there are clearly some interconnections between the gut and peripheral tissues like joint synovium and pancreas as gut derived lymphocytes have a propensity for selective homing to these organs14 ,15 when compared with PLN or skin.
Multistep nature of the extravasation cascade
Lymphocyte adhesion to vascular endothelium is much more than just a simple juxtaposition of two stationary cells. The shear force introduced by the blood flow and the rapid change of a lymphocyte from a non-adhesive blood cell into an adhesive cell in the extravascular compartment necessitates a complex cascade of successive but separable phases of cell–cell interaction.16 ,17
Initially, the blood borne lymphocyte is displaced from the central flow of the vessel and collides with the vascular endothelium in HEV (fig 1).18 ,19 These tethering contacts lead the cells to adopt a rolling behaviour—that is, they start to roll down the vascular lining at a velocity (10–50 μm/s) considerably slower than that of erythrocytes (typically >4000 μm/s). If the rolling cell locally receives appropriate triggering signals from the tissue, the lymphocyte can become activated. Then the cell can firmly bind to the endothelium, resist the shear stress and actually halt in the vessel. The stably bound cell finally seeks interendothelial junctions, and penetrates through them and the underlying basement membrane into the tissue. There it perceives other directional signals which help it to move within the tissue stroma.

Multistep model of lymphocyte homing into the gut. The five separate steps in lymphocyte trafficking from the blood into gut are shown. The molecules mediating different stages of binding of naive lymphocytes to Peyer’s patch (PP) high endothelial venules (HEV) and adhesion of activated immunoblasts to Peyer’s patch HEV or flat walled venules in lamina propria are shown. In the three first steps only molecules which have been shown to function in vivo under flow conditions in the gut are included. MAdCAM, mucosal addressin cell adhesion molecule; ICAM, intercellular adhesion molecule; LFA, lymphocyte function associated antigen; MAdCAMcho, MAdCAM with special carbohydrate structures.
Lymphocyte binding to endothelial cells involves adhesion molecules expressed on the surface of the two opposing cells types.1-3 ,20 During the multistep cascade molecules from many adhesion molecule superfamilies are successively brought into play (figs 1 and 2). The rapid and readily reversible contacts moderating tethering and rolling are mediated by selectins and their glycoprotein counterparts.21 These molecules are constitutively active and are presented on the tip of microvillous projections well above the planar cell surface, which renders them highly suitable for making initial contacts. The activation step is generally thought to involve binding of chemoattractants and other chemotactic cytokines secreted or presented by the endothelium to their seven-pass transmembrane receptors on lymphocytes.22 The ensuing signalling cascade triggers a change in the affinity and/or avidity of constitutively expressed inactive integrin molecules on the lymphocytes.23 The interaction of the activated integrins with their endothelial ligands, mainly belonging into a immunoglobulin superfamily, ultimately secures the contacts between the lymphocyte and the vessel wall. The molecular basis of the transmigration is poorly understood,24 but it may involve some of the same molecules that are used for stable arrest together with directional clues from chemoattractants and the cellular machinery that permits cell movement and penetration through the matrix.

{kind=link}
{kind=link}
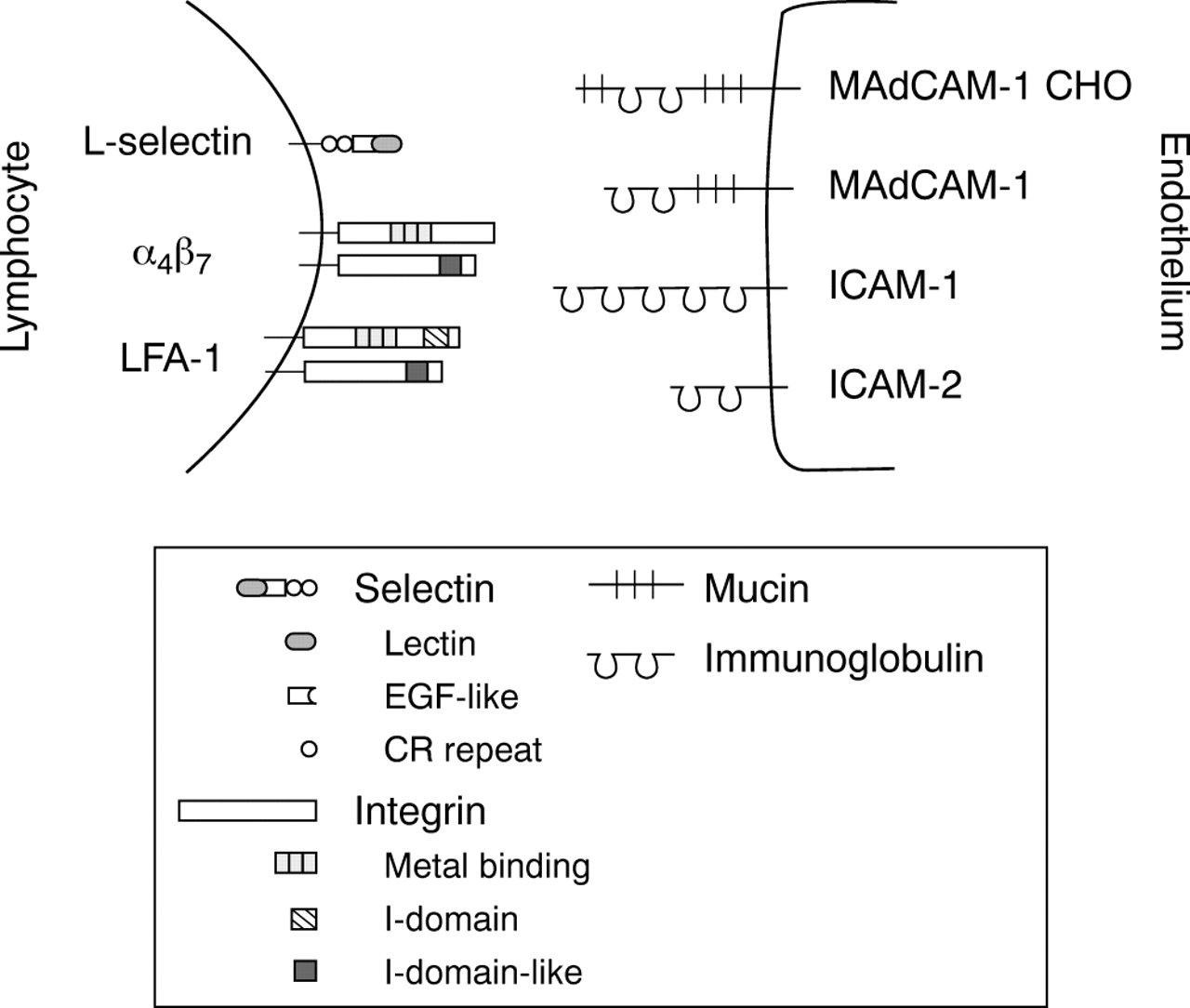
Structure of the mucosal adhesion antigens. The mosaic nature of L-selectin, mucosal addressin cell adhesion molecule (MAdCAM) 1 and special carbohydrate containing forms of MAdCAM-1 (MAdCAM-1 CHO) is shown together with the prototypic structure of integrins and intercellular adhesion molecules (ICAM). Lectin, C-type (calcium dependent) lectin domain; EGF-like, epidermal growth factor-like domain; and CR, complement regulatory protein-like motif in selectins. Metal binding = divalent cation binding repeats in α subunits of integrins; I-domain = inserted domain in αL subunit of lymphocyte function associated antigen (LFA) 1 and I-domain-like, a putative inserted domain-like area in β subunits of integrins.
The selectivity of the migration is determined by combinations of decisions at different steps of the emigration cascade.1 ,17 Thus, there are homing receptors relatively specifically expressed on gut seeking lymphocyte subpopulations and certain endothelial addressins are also almost exclusively synthesised in the intestine. Moreover, gut specific chemokines are likely to be found, although none of the 50 species characterised so far has been directly shown to function as a bowel specific leucocyte activator and chemoattractant. Firm adhesion and the later steps in the cascade are mediated by molecules which display little tissue selectivity as such, but nevertheless are absolutely required for a successful emigration cascade to be executed. The gut also serves as an example of a site that has been particularly thoroughly studied in respect of the overlapping function of distinct adhesion molecules (see later).18 ,25
Next we will present those adhesion molecules which are most relevant to lymphocyte homing into the gut. The description of the structure and function of many other molecules involved in leucocyte–endothelial adhesion may be found in recent review articles and in the original references therein.1-3 ,20
Molecular control of lymphocyte traffic in mucosal tissues under physiological conditions
LYMPHOCYTE RECEPTORS
L-selectin
Naive cells enter the mucosal sites via organised mucosal lymphoid tissues. They express high levels of L-selectin (CD62L), the principal homing receptor for PLN, and use it for tethering to mucosal endothelium. L-selectin interacts with a special form of mucosal addressin cell adhesion molecule-1 (MAdCAM-1) that is adorned with carbohydrate epitopes normally present in peripheral lymph node addressins (PNAd).26 These carbohydrate moieties function as ligands for L-selectin. The importance of L-selectin in mucosal homing of naive lymphocytes has been shown in studies in which anti-L-selectin antibody treatment partially inhibits homing of lymphocytes to Peyer’s patches.27 Moreover, L-selectin knockout mice show diminished trafficking of lymphocytes to Peyer’s patches,28 thus confirming the conclusions based on the antibody treatment studies.
L-selectin belongs to the family of selectins that all characteristically have a lectin domain, an epidermal growth factor (EGF) like area and variable numbers of repeats similar to those found in complement regulatory proteins (fig 2). The lectin domain is the most central structure of the molecule in ligand binding because it interacts efficiently with molecules containing certain sialylated, fucosylated, and sulfated carbohydrate structures.21
α4β7
Unlike naive lymphocytes, gut homing blasts and memory cells express very low levels of L-selectin or lack it completely.29 They do not need L-selectin to home to Peyer’s patches or the appendix.30 Instead, as a result of their maturation stage they can skip the selectin mediated phase in the multistep adhesion cascade and bind directly to endothelium via an activated integrin, α4β7.18α4β7 is also the key receptor for the entrance of lymphocytes into the lamina propria.18Interestingly, α4β7, the principal mucosal homing receptor, functions at several steps of the adhesion cascade by interacting with MAdCAM-1.25 It is able to act as a rolling receptor as well as a receptor mediating firm binding to endothelium. The central role of α4β7 in mucosal homing is unequivocally illustrated by the gene targeted mice lacking α4β7. In these animals, formation of mucosa associated lymphatic tissues is severly impaired and β7 negative lymphocytes fail to adhere to Peyer’s patch HEV.31
α4β7 is a member of the large family of integrins. As all integrins, it is a heterodimeric molecule consisting of non-covalently associated α and β chains. Naive lymphocytes express low levels of α4β7 but upon activation a significant amount of α4β7appears on the lymphocyte surface in a functionally active form.32
Lymphocyte function associated antigen
Naive lymphocytes and most likely also immunoblasts and memory cells utilise lymphocyte function associated antigen (LFA) 1 (CD11a/CD18, αLβ2) in the later steps of the adhesion cascade to arrest and transmigrate into mucosal sites.25 LFA-1 is a multifunctional adhesion molecule involved in various adhesive interactions in the immune system including antigen presentation and T cell mediated killing.33 Patients completely lacking LFA-1 and other β2 integrins die at an early age because their granulocytes are not able to extravasate to sites of inflammation.34However, lymphocyte homing is not severely impaired indicating that other molecules, especially α4 integrins, can compensate for the lack of LFA-1 in lymphocyte homing.
LFA-1 is a β2 integrin which are also known as leucocyte integrins. It shares the β chain with three other members of the leucocyte integrin family. LFA-1 needs to be either in the high affinity stage or clustered to interact efficiently with its endothelial cell ligands, intercellular adhesion molecules (ICAMs), via its inserted domain (I domain).23
αEβ7
It is obvious that following the transmigration step various intercellular and cell–matrix interactions occur when the lymphocyte travels and finds its final destination within the mucosa associated lymphatic tissues. One molecule thought be important in determining the destination of intraepithelial lymphocytes is the αEβ7integrin. It is prominently expressed on intraepithelial lymphocytes and is able to use E-cadherin present on epithelial cells as its ligand.35 Thus, this receptor–ligand pair may be important in retaining the intraepithelial lymphocytes in their anatomical location.
Other molecules
CD44 is a multifunctional adhesion proteoglycan molecule able to mediate lymphocyte rolling on hyaluronate and to activate LFA-1.36 ,37 Lymphocyte binding to mucosal endothelium can be partially blocked by anti-CD44 antibodies in vitro.38As CD44 deficient mice have not been reported to have abnormalities in their mucosa associated lymphatic tissues,39 the role of CD44 may not, however, be central in the homing of lymphocytes to normal mucosal sites.
In vivo inhibition studies have shown that blockade of the sialomucin CD43 results in diminished homing of lymphocytes into several organs including Peyer’s patches.40 In contrast, knockout animals show increased homing capacity of lymphocytes indicating that CD43 may regulate the function of other homing associated molecules.41
ENDOTHELIAL LIGANDS FOR LYMPHOCYTE RECEPTORS
MAdCAM-1
MAdCAM-1 is preferentially expressed on vascular endothelium at mucosal sites both in organised lymphoid tissues and in the lamina propria.42 In flat walled vessels in lamina propria MAdCAM-1 is not decked with carbohydrates functioning as L-selectin ligands and therefore in these vessels it mediates the entrance of blasts and memory cells, namely the cells in which α4β7 has already been activated.18 The importance of MAdCAM-1 in lymphocyte homing to mucosal sites is clearly shown by the significantly reduced homing of lymphocytes to mucosa associated lymphatic tissues after the treatment with anti-MAdCAM-1 antibody.43 Moreover, long term antibody treatment results in severe hypoplasia of Peyer’s patches.14
MAdCAM-1 belongs to the immunoglobulin (Ig) superfamily but has also a mucin-like domain that is heavilyO-glycosylated (fig 2). The Ig domains are crucial for binding to α4β7, whereas the mucin-like structure has been thought to act as a stalk facilitating the presentation of the Ig domains to α4β7.44
ICAMs
ICAM-1 (CD54) and ICAM-2 (CD102), members of the immunoglobulin superfamily (fig 2), are widely present in vasculature throughout the body. In the context of mucosal homing, they act as LFA-1 ligands mediating later steps in the extravasation cascade.25 The most N-terminal domain of ICAM-1 and -2 is the counter-structure for LFA-1, and dimerisation of ICAM-1 is reported to be important for the functional activity of the molecule.45
CHEMOKINES
Chemokines are thought to be required to trigger a rolling lymphocyte to become an adherent one by activating the integrins. However, most chemokines that are capable of attracting lymphocytes most likely exert their action within lymphoid tissues and thus are central in determining the travel routes and final destination of the lymphocytes within the organ. In contrast, secondary lymphoid-tissue chemokine (SLC) (also known as 6Ckine, Exodus-2, and thymus derived chemotactic agent 4) is strategically located in HEV both in Peyer’s patches and PLN.46 SLC increases α4β7 mediated rolling and firm arrest on recombinant MAdCAM-1 in vitro.47 Moreover, SLC augments LFA-1 mediated attachment of lymphocytes to ICAM-1.46 SLC is also expressed in T cell areas in the lymph nodes and may thus additionally help to guide lymphocytes within the lymphoid tissues.46
Chemokines are small soluble heparin binding proteins (excluding fractalkine which is larger and also has a transmembrane form) that function as chemoattractants for different subsets of leucocytes.22 They can be divided into four families that are defined by a cysteine signature motif: CXC (e.g. interleukin-8), CC (e.g. SLC), C (lymphotactin and CX3C (fractalkine)), where C is a cysteine and X any amino acid residue. Seven transmembrane domain molecules coupled to G-proteins serve as their ligands. Ligand binding leads to modulation of the activation of intracellular signal transduction enzymes (adenylate cyclase and phospholipase C) with subsequent signalling effects. Proteoglycan molecules are thought to present soluble chemokines to lymphocytes at the endothelial cell surface.22 ,48
Inflammation induced changes in the traffic of mucosal lymphocytes
During inflammation the same principles apply to the extravasation cascade as during physiological recirculation.49 Together with the haemodynamic changes the net effect of inflammatory activation of endothelial cells and leucocytes is an increased influx of both lymphocytes and granulocytes to sites of inflammation. Expression of existing endothelial cell molecules such as MAdCAM-1 and ICAM-1 is upregulated, and some inflammation inducible molecules such as E-selectin (CD62E) and P-selectin (CD62P), practically absent under normal circumstances, appear on the endothelium.29 ,42 ,50Kinetic studies on the expression of ICAM-1 and E- and P-selectin in mouse small intestine have been performed after experimental administration of cytokines and endotoxin. Tumour necrosis factor (TNF) α causes upregulation of ICAM-1 that reaches its maximum by six hours. Relatively rapid (<1 h) induction of P-selectin can be caused by histamine and more prolonged expression of P-selectin can be induced by endotoxin. Endotoxin induced E-selectin expression is maximal at three hours and thereafter it rapidly disappears from the endothelial cell surface in gut.50 E- and P-selectin, instead of controlling lymphocyte entrance may, however, rather facilitate the extravasation of granulocytes to inflammatory foci.
Vascular adhesion protein-1 (VAP-1) is one of those molecules that is expressed in low levels in normal intestine but is translocated from the intracellular granules to the endothelial cell surface upon elicition of inflammation. Significant induction of PNAd, which normally mediates lymphocyte traffic to PLN, also takes place during gut inflammation. PNAd together with VAP-1 also allow the entrance of the lymphocytes belonging to the PLN recirculatory pool to inflamed mucosal sites.29 ,51 Aberrant homing of these cells to mucosal sites may be a central element in the pathogenesis of inflammatory bowel disease (IBD), because these cells most likely actively participate in the process resulting in tissue damage.
Mucosal recirculation and vaccine development
Organ specific lymphocyte trafficking routes have direct implications for the development of new vaccines, especially in connection to selection of the site of vaccination. As lymphocytes activated in a certain organ system tend to home back to sites of their activation, immunisation protocols that evoke the response in a subset of lymphocytes bearing high levels of mucosal homing receptor, α4β7, will presumably provide the best protection against mucosal pathogens. The importance of the vaccination site to the nature of the immune response has been confirmed experimentally by vaccinating volunteers orally or parenterally with the same antigen. The antigen given orally induces significantly more antibody producing cells with a α4β7phenotype than the same antigen given parenterally.52 ,53In an analogous way, T cells activated via mucosal routes tend to bear more α4β7 than those activated systemically.54
An exciting possibility is the regulation of the expression of homing associated molecules of the effector cell populations by cytokines and other factors or drugs, and thus redirect the homing of the protection providing lymphocyte populations in the body to sites where the elimination of a pathogen is needed. However, much basic and applied work needs to be performed to elucidate the feasibility of this approach.
Therapeutic intervention in mucosal homing
As the inflammatory component is a major feature in a vast number of gastrointestinal diseases, the advent of molecular understanding of leucocyte trafficking in mucosa has fuelled enthusiasm for the development of novel anti-inflammatory strategies. The basic idea is that, regardless of the aetiology, excessive, prolonged or otherwise inappropriate gut inflammation can be terminated by blunting the multistep adhesion cascade at any step. Most studies testing this concept have used mouse monoclonal antibodies or their humanised versions, but in future receptor or ligand analogs and small molecular drugs are the likely formulas to be used in clinics.
Inflammatory bowel disease has been succesfully targeted using anti-adhesion therapy. In initial trials anti-α4 monoclonal antibodies (recognizing both α4β1 and α4β7) noticeably relieved gut inflammation in a cotton-top tamarin model.55 An antibody specific for the principal mucosal homing receptor, α4β7, has also been used in the same model. In these chronically colitic animals the stool consistency improved within a day, the number of cells in the inflammatory infiltrate decreased after intravenous treatment with the antibody and the favourable effects were long lasting.56 Similar findings have been produced in a mouse model of IBD, both after blocking α4β7 and after targeting MAdCAM-1.57 In another approach, antisense oligonucleotides preventing synthesis of ICAM-1 were administred to patients with IBD.58 Although the number of patients enrolled in the study was small, the authors concluded that the treatment was safe. However, it did not seem to alleviate significantly most symptoms and signs of the disease.
Adhesion molecules also play a central role in the pathogenesis of radiation enteropathy. Blocking of LFA-1 and ICAM-1 has been shown to be benefical in treating this postirradiation syndrome in rats.59 Ischemia–reperfusion injury of the gut is also notably attenuated if the function of P-selectin is neutralised with monoclonal antibodies.60 The leucocytic infiltration caused by Clostridium difficile toxin A can also be abrogated to a large extent if the function of the leucocyte–endothelial adhesion molecules (e.g. P-selectin and LFA-1), induced indirectly by this toxin, is blocked.61 These examples have been mentioned just to illustrate the concept that anti-adhesive therapy can be a broad spectrum and at the same time a precise anti-inflammatory device, only affecting the involved gut area if the target molecules are selected appropriately. Suprisingly, very few, if any, side effects have been reported in these trials. This suggests that short term interference in mucosal recirculation can be well tolerated without jeopardising the vital need for mucosal immune homoeostasis.
Concluding remarks
Molecular understanding of the mechanisms which guide lymphocyte trafficking into the gut is rapidly increasing. At the moment we know that in this anatomical compartment lymphocyte α4β7 and endothelial MAdCAM-1 are the most critical elements, which nevertheless need the help of other molecules for successful extravasation to occur. Changes in lymphocyte homing into the gut may cast light on the pathogenesis of intestinal inflammation. Moreover, there are realistic expectations that molecular dissection of this recirculation pathway will help to design rational vaccination regimens and new anti-inflammatory strategies.
Abbreviations used in this article
- HEV
- high endothelial venules
- PLN
- peripheral lymph nodes
- MAdCAM
- mucosal addressin cell adhesion molecule
- PNAd
- peripheral lymph node addressins
- EGF
- epidermal growth factor
- ICAM
- intercellular adhesion molecule
- SLC
- secondary lymphoid-tissue chemokine
- TNF
- tumour necrosis factor
- VAP
- vascular adhesion protein
- LFA
- lymphocyte function associated antigen