Article Text

Abstract
Background and aim: Somatostatin analogues may have antifibrotic properties in the lung. The aim of this study was to evaluate the expression of the five somatostatin receptors sst1 to sst5 in normal and fibrotic mouse lung and the action of SOM230 (pasireotide), a new somatostatin analogue with a long half-life, in bleomycin induced lung fibrosis and in human lung fibroblasts in vitro.
Methods: After intratracheal injection of bleomycin, C57Bl6 male mice received one daily subcutaneous injection of SOM230 or saline. The lungs were evaluated on days 3, 7 and 14 after administration of bleomycin.
Results: We found that all somatostatin receptors were expressed in the normal mouse lung. The sst2 receptor mRNA expression was increased after bleomycin. SOM230 improved mice survival (69% vs 44%; p = 0.024), reduced lung collagen content at day 14 and decreased lung collagen-1 mRNA at day 7. SOM230 reduced bronchoalveolar lavage inflammatory cell influx at day 3, decreased lung connective tissue growth factor mRNA and transforming growth factor (TGF) β mRNA and increased lung hepatocyte growth factor and keratinocyte growth factor mRNA. The sst2 receptor was strongly expressed in the human lung (normal or fibrotic), particularly by fibroblasts. In vitro, SOM230 reduced BrdU incorporation by control human lung fibroblasts cultured under basal conditions or with TGFβ, and reduced alpha-1 collagen-1 mRNA expression in TGFβ stimulated fibroblasts.
Conclusion: We conclude that SOM230 attenuates bleomycin induced pulmonary fibrosis in mice and human lung fibroblasts activation. This study points to a potential new approach for treating pulmonary fibrotic disorders.
Statistics from Altmetric.com
Idiopathic pulmonary fibrosis (IPF) is a chronic lung disease which carries a poor prognosis. To date, no treatment has been demonstrated to prevent the evolution of fibrosis in patients with IPF and new therapeutic opportunities are needed. While the pathophysiology of pulmonary fibrosis is poorly elucidated, it is characterised at the microscopic level by fibroblast accumulation and extracellular matrix deposition in the lung, leading to alveolar wall thickening and loss of elasticity. Apart from chronic lung inflammation, which probably participates in the pathophysiology of the disease,1 the recruitment, activation and proliferation of fibroblasts are now considered key features during the development of pulmonary fibrosis, and therapeutic agents targeting fibroblasts are being evaluated in IPF.2
Somatostatin is an endogenous cyclic peptide initially identified as a regulator of growth hormone secretion. Five somatostatin receptors—sst1, sst2A, sst3, sst4 and sst5—have been described in humans, that bind somatostatin equally. Although the relative expression and localisation of these receptors vary within organs, their expression in the lung has been poorly studied. In a previous study, our group has shown that human lung fibroblasts, from normal and fibrotic lung, bound radiolabelled somatostatin in vitro.3 We also evidenced an increased uptake of radiolabelled octreotide (a somatostatin agonist) in the lung in patients with IPF compared with control patients.3 Most importantly, the octreotide uptake correlated with the severity of lung fibrosis.3 Taken together, these data suggested that somatostatin receptors were increased in IPF lungs.
Somatostatin and its pharmacological analogues display antifibrotic activities in vitro and in vivo in different experimental settings.4 The pathways involved in this antifibrotic effect are not completely elucidated but inhibition of transforming growth factor β1 (TGFβ1) production5 6 as well as a degree of anti-inflammatory actions7 may contribute to the protective effect. However, the antifibrotic effect of somatostatin and its analogues has never been evaluated in the lung.
SOM230 (pasireotide) is a new somatostatin analogue that binds four out of the five sst receptors (sst1, sst2A, sst3 and sst5) and has a long half-life which makes it suitable for therapeutic use.8 In this study, we evaluated the antifibrotic effect of SOM230 in a bleomycin induced lung fibrosis model in mice. We quantified the expression and localisation of somatostatin receptors (sst1 to sst5) in the normal and fibrotic lung, and we evaluated whether SOM230 modulated the TGFβ and connective tissue growth factor (CTGF) profibrogenic pathways. Furthermore, we characterised the effect of SOM230 on human lung fibroblasts in vitro.
MATERIAL AND METHODS
Bleomycin animal model
Male C57BL/6J mice (R Janvier, Le Genest St Isle, France), aged 6–7 weeks, were kept in accordance with INSERM rules. On day 0, mice were administered 80 μg of bleomycin hydrochloride (Bleomycine Bellon; Aventis, Paris, France) intratracheally, and were randomly assigned to receive either SOM230 25 μg/kg/day (bleo+SOM mice) or 2.5 μg/kg/day, or an equal volume of saline (bleo+vehicle mice). SOM230 was diluted in 100 μl of saline and subcutaneously administered daily, beginning immediately after injection of bleomycin. Mortality was assessed daily over a 14 day period. For further analysis, animals were sacrificed on day 3, 7 or 14 after administration of bleomycin.
To evaluate the effect of late treatment, SOM230 (25 μg/kg/day) or vehicle was given from day 5 to day 13. The animals were killed on day 14. Naïve mice were used as controls. The doses of SOM230 used in this study were similar to those used in previous experiments.9
In further experiments, controls animals received SOM230 (25 μg/kg/day) or vehicle for 3, 7 or 14 days, and were sacrificed at each time point (n = 6 per time).
Tissue sampling, lung histology, TGFβ1 assay and mRNA analysis
At specified time points, the animals were killed and the lungs were freed of blood by perfusion. Bronchoalveolar lavage (BAL) was performed with 1 ml of saline. Total and differential cell counts were estimated. BAL supernatant was stored at −80°C until TGFβ1 assay.
Total TGFβ1 was measured after acidification of the samples according to the manufacturer’s instructions (Quantikine; R&D System Europe, Lille, France).
Semiquantitative assessment of lung injury used the grading system described by Inoshima and colleagues.10
Lung collagen was quantified in snap frozen right lungs with the Sircol collagen assay (Biocolor Ltd, Newtownabbey, UK).
RNA extraction and real time quantitative PCR were performed on left mouse lungs using standard protocols (see online supplement). Expression of the gene of interest was expressed as a ratio to the ribosomal protein L13 (RPL13) gene, as previously described.11
Immunohistochemical detection of somatostatin receptors
The sst2 receptor was detected by immunohistochemistry on mice samples with a rabbit polyclonal anti-mouse sst2A antibody (ref SS-800; Biotrend, Cologne, Germany) (1/400 dilution). Immunodetection was performed on frozen human lung tissue samples (from three controls and five patients with IPF, diagnosed according to the American Thoracic Society/European Respiratory Society consensus criteria12—the origin of lung samples has been previously described11), and cultured human fibroblasts, using a polyclonal rabbit antibody to human somatostatin receptor type 2B (ref SS-860; Biotrend) (1/1000 dilution). Standard protocols were used (see online supplement).
Data analysis
Data are expressed as median (extremes values). Data analysis was performed with the GraphPad Prism 4.0 software (GraphPad software, San Diego, California, USA). Survival between experimental and control groups was studied with a Kaplan–Meier analysis using the log rank test. Experimental groups were compared first with a non-parametric analysis of variance equivalent (the Kruskall–Wallis test) and, if significant, pairwise comparisons were performed with the Mann–Whitney U test. Comparison of histological scores on day 14 was performed with the Fisher exact test after grouping the scores 0 and 1 and the scores 2 and 3. p Values below 0.05 were considered significant.
RESULTS
Somatostatin receptors are present in the lung and their expression is modulated after bleomycin instillation
In naive mice, all (sst1 to sst5) somatostatin receptor mRNA was detected in the lung, with very low levels of expression of sst5 (fig 1). After bleomycin instillation, expression of sst2A and sst5 mRNA was strongly increased (maximum on day 7) whereas expression of sst1 and sst4 mRNA was decreased (fig 1).

Treatment with SOM230 of bleomycin injected mice temporarily increased expression of sst1 (sst1/RPL13 mRNA ratio = 2.9 (2.5 to 24) on day 7 in bleo+SOM mice vs 0.9 (0.4 to 3.5) for bleo+vehicle mice, n = 4; p = 0.015), and partly inhibited the increase in expression of sst2A (sst2/RPL13 mRNA ratio = 7.4 (2.8 to 10.6) on day 7 in bleo+SOM mice vs 19 (14.1 to 36.3) for bleo+vehicle mice, n = 4; p = 0.015) (fig 1).
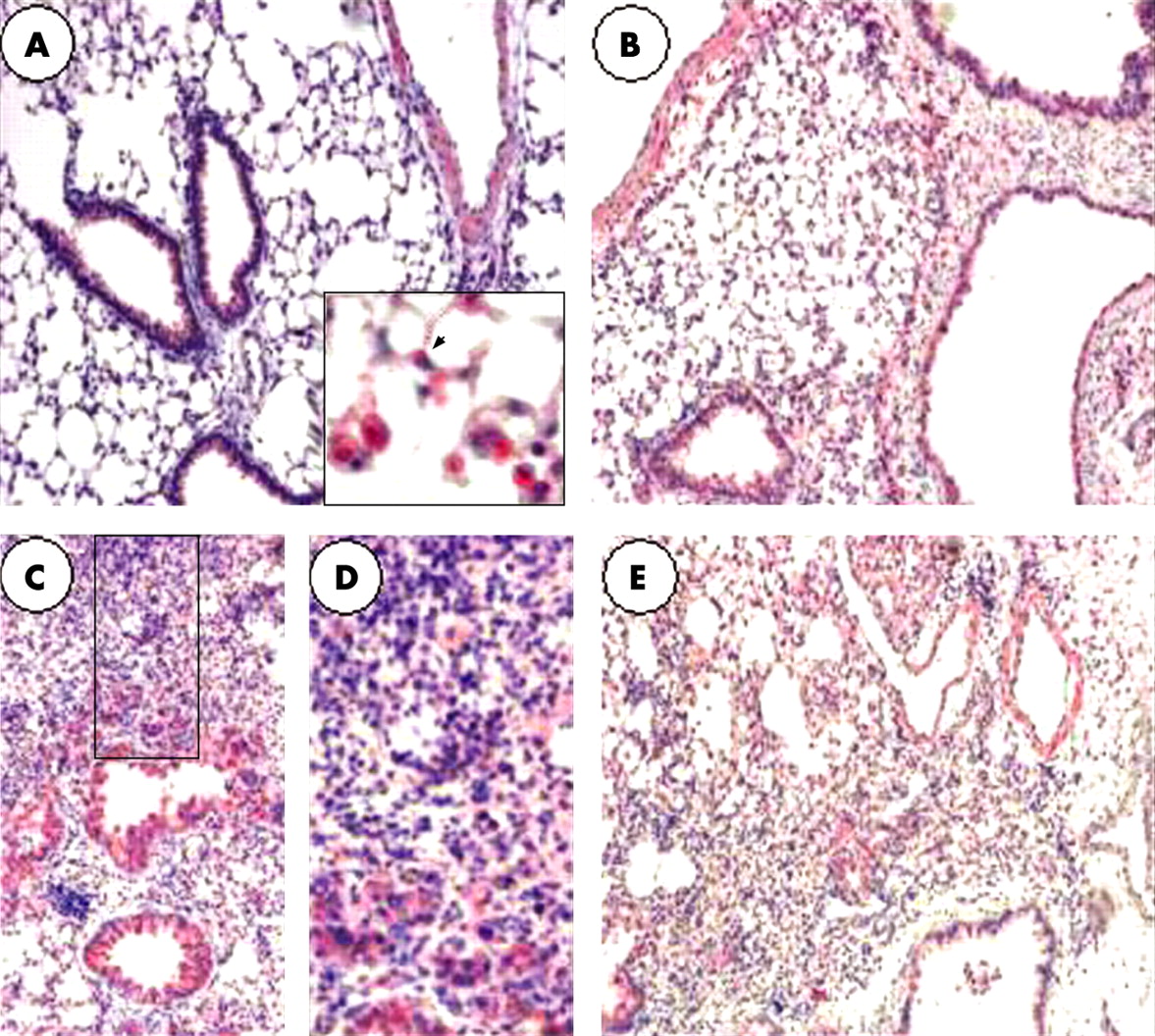
We characterised the cellular localisation of the sst2A receptor by immunohistochemistry. In the normal murine lung, sst2A was localised to alveolar type 2 pneumocytes, bronchial epithelial cells, alveolar macrophages, arterial smooth muscle cells and some endothelial cells. After bleomycin instillation, the number of sst2A positive cells increased, with a maximum on day 7, essentially because of increased infiltration of sst2A positive cells (macrophages and monocytes) in inflammatory areas (fig 2).
Treatment with SOM230 improved mice survival after bleomycin instillation
Intratracheal injection of bleomycin induced significant mortality among mice, which typically occurred from day 7. Treatment with SOM230 (25 μg/kg/day) increased the 14 day survival of mice instilled with bleomycin to 69% (n = 36) compared with 44% in the bleo+vehicle group (n = 41, p = 0.02). Treatment with SOM230 at a lower dose (2.5 μg/kg/day) did not improve survival (14 day survival 32%, n = 22, p = 0.46) (fig 3).

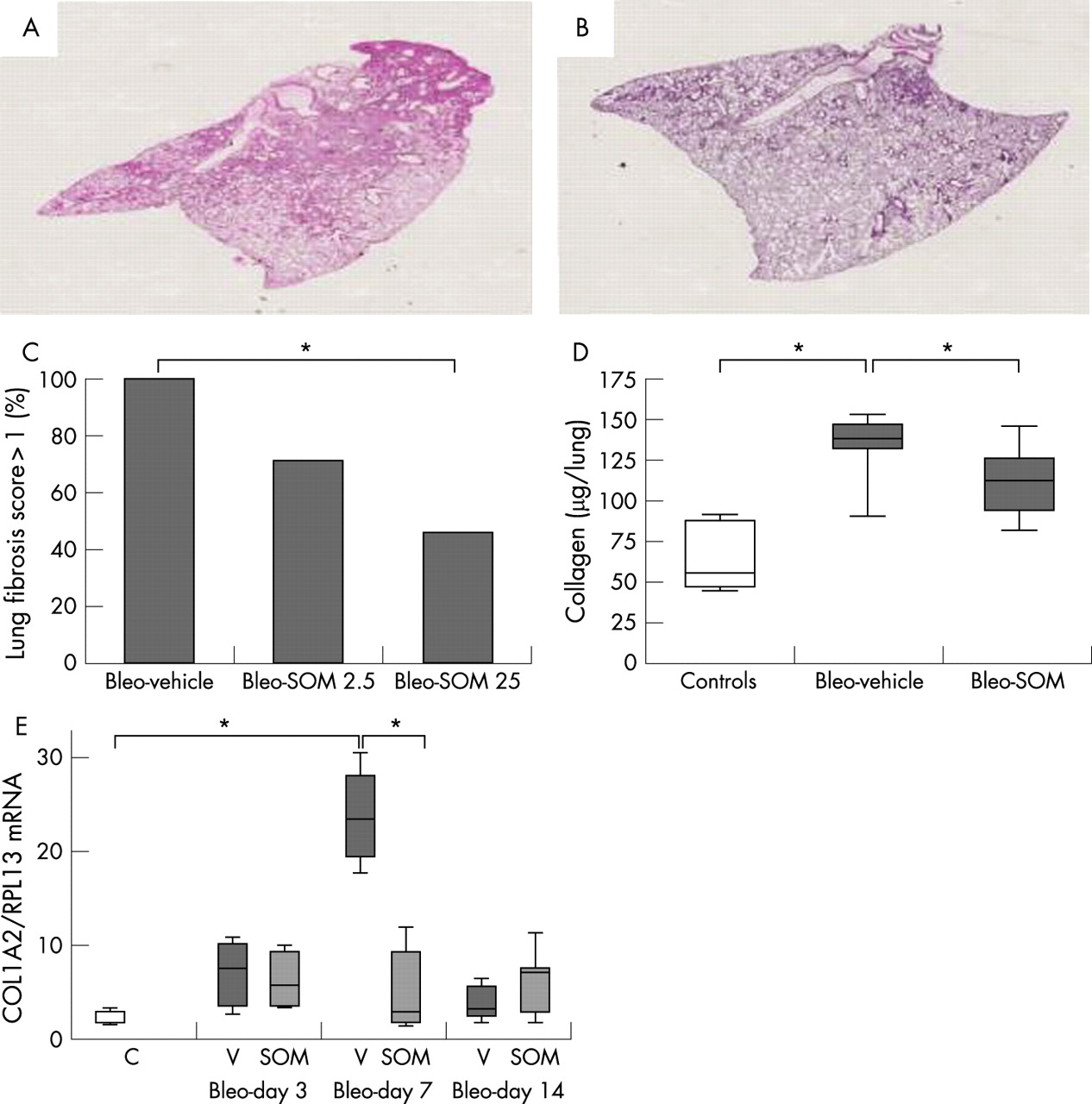
Treatment with SOM230 attenuated bleomycin induced pulmonary fibrosis
We analysed the histological changes in the lung of surviving mice at day 14 following bleomycin instillation in bleo+SOM mice (25 μg/kg/day, n = 11; 2.5 μg/kg/day, n = 7) compared with bleo+vehicle mice. As shown in fig 4, treatment with SOM230 25 μg/kg/day improved the pathological score at day 14 compared with bleo+vehicle mice (p = 0.03) whereas the lower dose (2.5 μg/kg/day) had no effect. In view of these favourable results, the 25 μg/kg/day dose of SOM230 was used in further experiments.
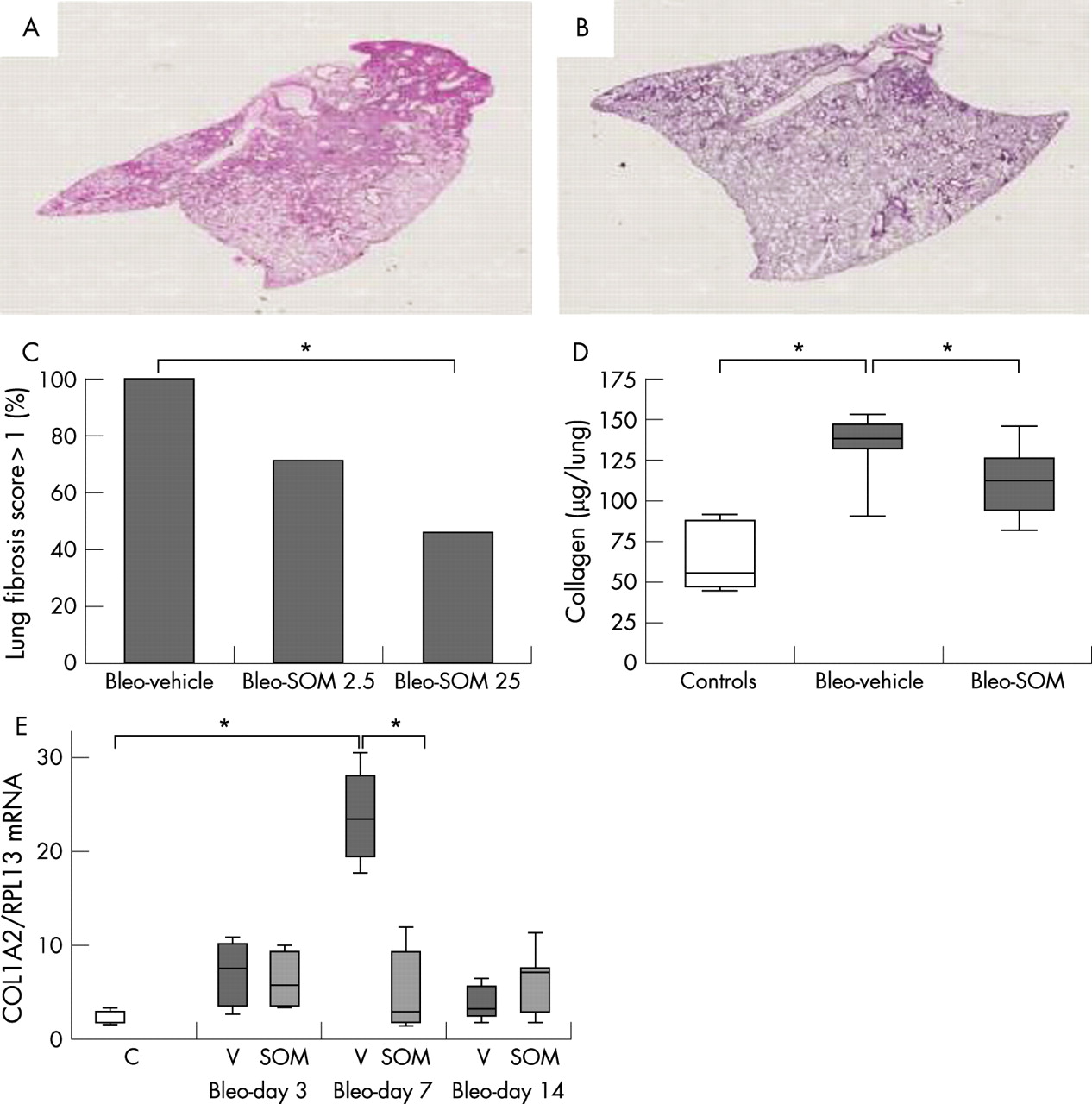
SOM230 reduced lung collagen concentration
Treatment with SOM230 reduced lung collagen content on day 14 (112 (21 to 146) μg/lung in bleo+SOM mice versus 139 (90 to 153) μg/lung in the bleo+vehicle mice, p = 0.01) compared with 55 μg/lung (45–90) in naive mice (p = 0.007 and p = 0.003 vs bleo+SOM mice and bleo+vehicle, mice respectively) (fig 4).
In bleo+vehicle mice, alpha-2 collagen-1 (COL1A2) mRNA lung content was increased on day 3 after bleomycin instillation, peaked on day 7 and then decreased on day 14 (fig 4). SOM230 treatment strongly inhibited the peak of COL1A2 mRNA expression in the lung on day 7 (COL1A2/RPL13 mRNA ratio = 2.6 in the bleo+SOM mice vs 23 in the bleo+vehicle mice; p = 0.01). SOM230 had no effect on COL1A2 mRNA contents at other time points. These results show that the reduction in collagen content on day 14 was preceded by a reduction in COL1A2 mRNA on day 7.
Treatment with SOM 230 reduced lung inflammation and inhibited the increase in TGFβ1 and CTGF
Treatment with SOM230 reduced the alveolar inflammatory cell influx at early times, as assessed by the reduction in total BAL cellularity, BAL macrophages and BAL lymphocyte counts on day 3 (table 1). On day 7 and day 14, BAL cellularity was similar in bleo+SOM and bleo+vehicle mice (data not shown).
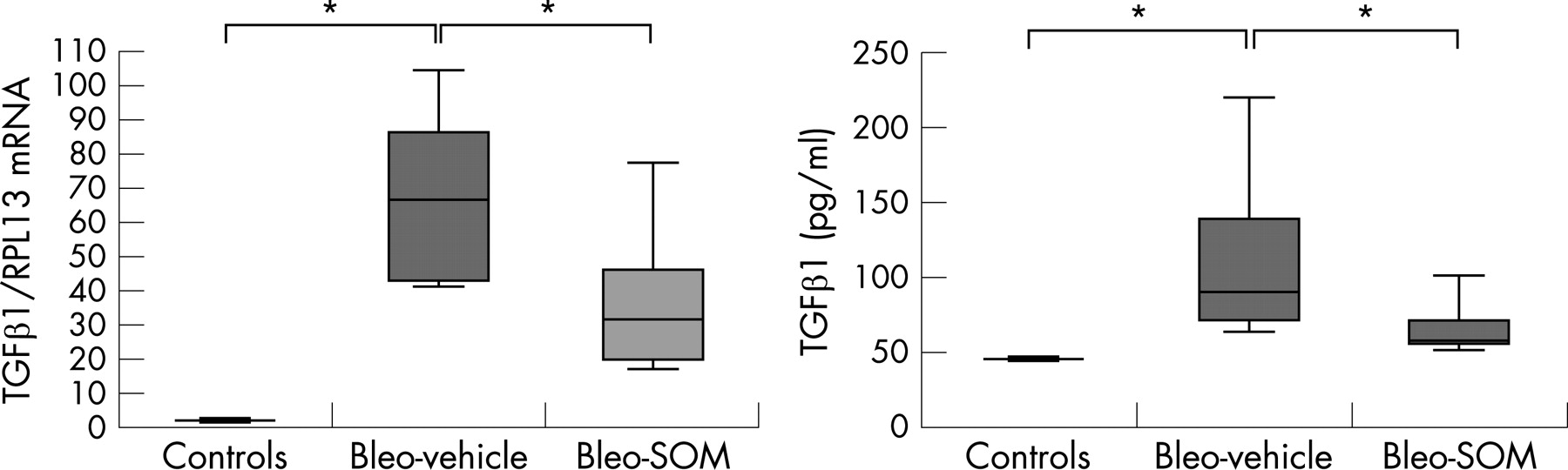
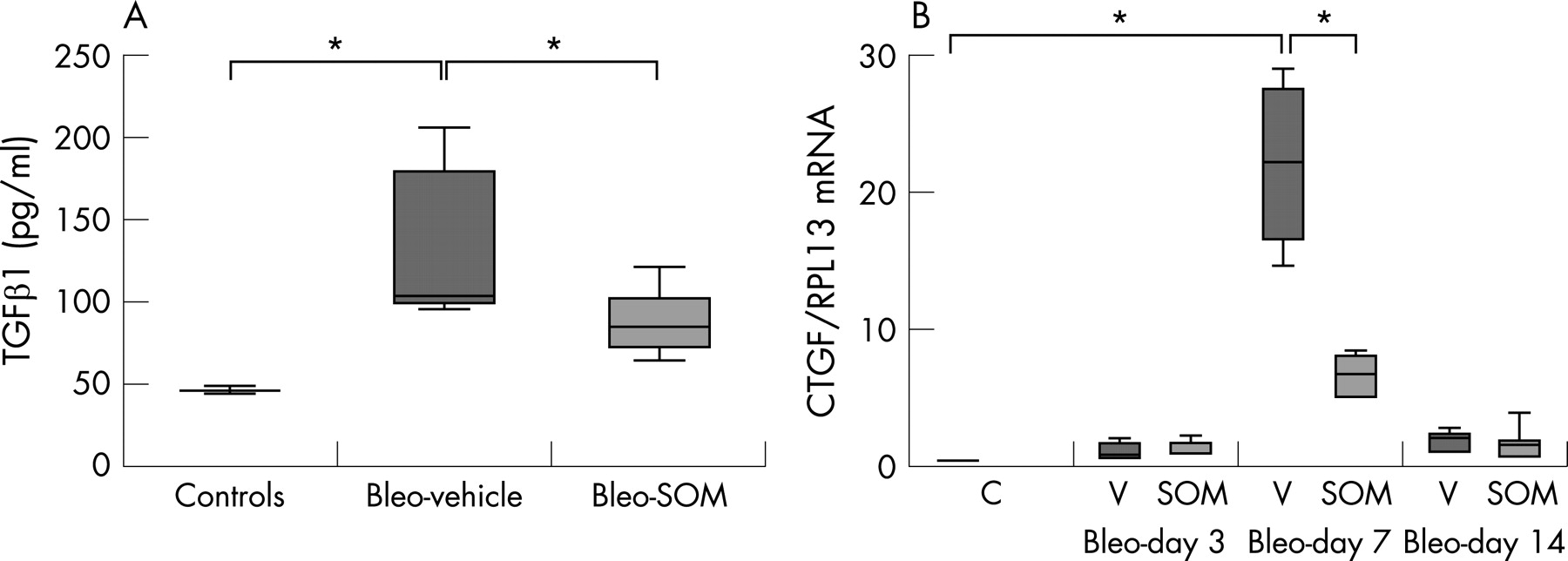
TGFβ and CTGF are two potent profibrogenic factors. Lung TGFβ mRNA content tended to be lower in bleo+SOM mice on day 7 after bleomycin administration compared with bleo+vehicle mice (TGFβ/RPL13 mRNA ratio 2.6 (0.9 to 3.2) in bleo+SOM mice vs 4.8 (2.3–16.1) in bleo+vehicle mice; p = 0.11). TGFβ1 concentration in BAL fluid was lower in bleo+SOM mice on day 14 (85 pg/ml (64 to 121)) compared with bleo+vehicle mice (103 pg/ml (96 to 206), n = 9; p = 0.01) (fig 5).

Similarly, SOM230 strongly inhibited the increase in CTGF mRNA induced by bleomycin. After bleomycin instillation, CTGF mRNA content was increased by day 3, with a maximum by day 7. Treatment with SOM230 reduced by 70% the CTGF/RPL13 mRNA ratio on day 7 (ratio = 6.4 (4.7–8.3), n = 5) compared with bleo+vehicle mice (ratio = 22.2 (14.6–29.2), n = 4; p = 0.01) (fig 5).
Furthermore, SOM230 promoted the expression of hepatocyte growth factor (HGF) and keratinocyte growth factor (KGF) (two antifibrotic mediators) after bleomycin, and reduced lung CTGF mRNA content by 30% when given to naive mice (see online data supplement).
Effect of late treatment with SOM230
In a further set of experiments, we assessed the protective effect of SOM230 given daily after bleomycin instillation. Mice were started with SOM230 (25 μg/kg/day) from day 5 after bleomycin instillation until day 13.
In these experiments, the pathological score (score >1: 5/9 bleo+vehicle animals vs 3/8 bleo+SOM animals) and lung collagen content (79.4 (61 to 128) μg/lung in bleo+SOM mice vs 93.4 (56 to 130) μg/lung in bleo+vehicle mice) tended to be lower in bleo+SOM mice (NS). TGFβ mRNA and TGFβ BAL concentrations were strongly reduced in bleo+SOM mice on day 14 (fig 6). There was no significant effect on lung CTGF mRNA content on day 14.
Immunohistochemical detection of the sst2 receptor in human lung (fig 7)
Expression of the sst2 receptor was very low in the normal lung, and was essentially detected in alveolar macrophages and in isolated alveolar cells primarily located in the corners of the alveoli, representing most probably type 2 pneumocytes. The receptor was not detected in endothelial cells. In the fibrotic lung, we observed strong expression of this receptor in hyperplastic alveolar epithelial cells, endothelial cells and fibroblasts as well as a predominance of the expression of the sst2 receptor in areas of dense fibrosis.
In vitro, fibroblasts cultured from normal lung and fibrotic lung strongly expressed the sst2 receptor, as assessed by immunohistochemistry (fig 7). The intensity of labelling was similar in the two groups.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
To our knowledge, this is the first study describing the antifibrotic action of a somatostatin analogue in pulmonary fibrosis. Our results demonstrate (1) that somatostatin receptors are expressed in the lung and are increased during bleomycin induced lung fibrosis in mice and in IPF in humans and (2) that SOM230, a new pharmacological analogue of somatostatin, has a protective effect in bleomycin induced pulmonary fibrosis in mice, as daily treatment with SOM230 improved survival, reduced the lung pathological score and lung inflammation, reduced lung collagen content and lung collagen mRNA expression, inhibited expression of profibrotic mediators (TGFβ and CTGF) and increased expression of two antifibrotic mediators (HGF and KGF). Although less marked, the antifibrotic effect of SOM230 was also apparent when treatment with SOM230 began 5 days after bleomycin instillation. This suggests that SOM230 may have a therapeutic effect on an ongoing fibrotic process. Furthermore, in in vitro studies (see online supplement), incubation with SOM230 inhibited the proliferation and collagen-1 mRNA expression of human lung fibroblasts.
Emerging data suggest that somatostatin may have interesting antifibrotic properties, both in vitro and in vivo, which we have confirmed in this study using an in vivo model of lung fibrosis.
Firstly, we demonstrated expression of somatostatin receptors in the lung. All sst receptor mRNA was detected in mice lung, although the sst5 receptor was expressed at a very low level. Interestingly, sst receptor mRNA expression appeared to be differentially modulated following bleomycin induced lung fibrosis: expression of sst2A and sst5 mRNA was strongly increased, expression of sst1 and sst4 mRNA was decreased whereas expression of sst3 mRNA remained unchanged. The increased expression of the sst2 receptor is of paramount importance as the sst2 receptor is thought to play a prominent role in the biological effects of somatostatin analogues.13 Increased mRNA expression was associated with an increase in sst2 positive cells on immunohistochemistry. The latter results are reminiscent of our previous report of increased uptake of radiolabelled octreotide in patients with IPF.3
Interestingly, both inflammatory cells and resident lung cells expressed the sst2 receptor in mice and human samples, as detected by immunohistochemistry. In the bleomycin mouse model, increased expression of sst2 was due, at least in part, to an influx of inflammatory cells, particularly monocytes and macrophages. Expression of sst2 has been shown to be upregulated in activated monocytes.14 Our results support data indicating that somatostatin analogues exert anti-inflammatory effects on lymphocytes, macrophages and neutrophils.15–20 Interestingly, SOM230 reduced alveolar inflammatory cell influx on day 3, demonstrating an anti-inflammatory action of the compound in this model.
Although somatostatin and its analogues have shown interesting antifibrotic properties in a number of animal models, they have never been tested previously in a model of pulmonary fibrosis. Octreotide is the most extensively studied molecule in this field. In rats, octreotide inhibits the development of liver fibrosis induced by extrahepatic biliary obstruction or carbon tetrachloride poisoning,21 22 inhibits oesophageal or digestive fibrosis induced by radiation6 23 and decreases the accumulation of connective tissue in a murine model of hepatic schistosomiasis.24 Somatostatin and its analogues develop their antifibrotic action through different mechanisms. Somatostatin has been shown to promote fibroblast apoptosis in vitro through its sst2 receptor25 whereas octreotide inhibits the proliferation of skin and orbital fibroblasts.26 27 In vitro, somatostatin modulates fibroblast proliferation through activation of the ERK and MAPkinase pathways.4 We show in this study that SOM230 inhibits human lung fibroblast proliferation in vitro. Some data suggest that somatostatin analogues may act through inhibition of TGFβ production5 23 28 although TGFβ inhibition was not apparent in all models studied.6 TGFβ is the prototypical profibrotic mediator in the lung. TGFβ has been shown to increase expression of the sst2 receptor in mice, through a Smad4 dependent pathway.29 Broad expression of sst receptors in the lung indicates that beside fibroblasts, SOM230 may have many cellular targets, both on resident cells and inflammatory cells, all possibly contributing to its protective action.
Interestingly, SOM230 seemed to downregulate macrophages and lymphocyte lung infiltration on day 3, but not neutrophils (table 1). This phenomenon could be due to a direct effect of SOM230 on leucocyte survival. Indeed, somatostatin inhibits proliferation and promotes apoptosis in lymphocytes30 and reduces macrophage viability,31 but such an effect has not been reported with neutrophils. SOM230 could also differentially modulate expression of chemoattractants and adhesion molecules. We did not specifically address this point. In one study, somatostatin did not modify neutrophil migration in vitro in response to different stimuli.32
The protective effect of SOM230 in our study is consistently associated with a decrease in TGFβ1 expression, observed both when SOM230 was given from day 0 to day 14, or when given later, from day 5 to day 14. Reduced expression of TGFβ1 may contribute to strong inhibition of lung CTGF expression that we observed on day 7 as CTGF expression is selectively induced by TGFβ1.33 In vitro, octreotide, a somatostatin analogue, inhibits expression of TGFβ and CTGF mRNA by hepatic stellate cells.34 Whether somatostatin analogues directly modulate CTGF expression is unknown but may be supported by our observation that SOM230 reduced lung CTGF mRNA in control mice treated with SOM230.
Interestingly, our results suggest that SOM230 could exert its antifibrotic properties through increased expression of antifibrotic mediators, such as HGF and KGF (see online supplement). HGF and KGF are produced mainly by fibroblasts.35 Our group has previously shown that human lung fibroblasts cultured from the IPF lung have a reduced capacity to secrete HGF and KGF36 37 and to activate proHGF.11 Reduced expression of TGFβ1 by SOM230 explains the increase in HGF and KGF expression as TGFβ1 is a known inhibitor of HGF and KGF expression.35
We observed that endothelial cells expressed the sst2 receptor in the lung, both in mice and in human IPF lung. This result is in agreement with previously reported evidence of the effect of SOM230 in reducing endothelial cell proliferation in vitro38 and its action as an antiangiogenic drug.39 This is particularly interesting as increased vascularity is a hallmark of IPF40 and reduction of aberrant angiogenesis reduces the development of bleomycin induced pulmonary fibrosis.41 These findings could further explain the antifibrotic effect of SOM230.
We have also shown that SOM230 inhibits collagen-1 mRNA expression by human lung fibroblasts stimulated in vitro with TGFβ1 (see online data supplement). Interestingly, maximal inhibition was obtained with low concentrations of SOM230 (10−10 M) that are likely to be obtained in vivo. Somatostatin has been shown previously to directly inhibit expression of collagen and α-smooth muscle actin in hepatic stellate cells.42 In that study, inhibition of collagen expression was also obtained only with low concentrations of somatostatin (10−9 M); the authors attributed this effect to desensitisation of somatostatin receptors with higher doses.42
In summary, the encouraging results of this study indicate that SOM230 exerts a protective antifibrotic action in vivo in the bleomycin model. SOM230 demonstrates its antifibrotic action, both through an anti-inflammatory effect and a direct effect on lung fibroblasts, as evidenced by our in vitro results obtained on human lung fibroblasts. The encouraging results obtained from this study provide a rationale for clinical trials based on somatostatin analogues in patients with pulmonary fibrosis.
Acknowledgments
SOM230 was a generous gift of Novartis Pharma (Basel, Switzerland). We thank Olivier Thibaudeau and the Plateau de Morphologie IFR 02 for his technical help in the lung histology study.
REFERENCES
Supplementary materials
web only appendices 63/3/258
Files in this Data Supplement:
Footnotes
-
↵ * R Borie and A Fabre contributed equally to this work.
-
Funding: AF was supported by a grant from the Collège des Professeurs de Pneumologie and the Association Française pour la Recherche Thérapeutique. SM-A was supported by a grant from the Fondation pour la Recherche Médicale (Prix Mariane Josso). Part of this work was supported by the Legs Poix (Chancellerie des universités de Paris). PS is the recipient of a Contrat d'Interface Inserm-Assistance Publique/Hôpitaux de Paris.
-
Competing interests: None.
-
Ethics approval: Yes.
Linked Articles
- Airwaves