


Abstract
Human tumor cells lines with acquired resistance to cisplatin (DDP) and carboplatin (CBDCA) are often cross-resistant to copper and vice versa, and some DDP-resistant cells overexpress the copper export pump ATP7B. We sought to demonstrate that ATP7B directly mediates resistance to DDP and CBDCA by stably transfecting human carcinoma cells with a vector designed to express ATP7B. Increased expression of ATP7B rendered all three cell lines tested more resistant to a 1-h exposure to DDP (1.6–2.6-fold), CBDCA (1.5–1.6-fold), and copper (1.2–1.4-fold). The effect of ATP7B on the cellular pharmacology of 64Cu and [14C]CBDCA was investigated in more detail using one cell pair (2008 cells transfected with an empty vector or an ATP7B-expressing vector). In the 2008/ATP7B subline, steady-state copper levels were decreased under both basal and copper-supplemented conditions, as was steady-state CBDCA content upon exposure to 50 μM [14C]CBDCA. Over the first 5 min, the average rate of accumulation of copper and CBCDA in the 2008/ATP7B cells was reduced by 37 and 61%, respectively. Efflux was more rapid from 2008/ATP7B cells for both copper and CBDCA. Two-compartment modeling indicated that the second phase of efflux was increased by a factor of 3.9-fold for CBCDA and to an even greater extent for copper. We conclude that expression of ATP7B regulates sensitivity to CBDCA as well as to DDP and copper and that a transporter that normally mediates copper homeostasis modulates the cellular pharmacology of CBDCA.
Acquired resistance to DDP and CBDCA is currently believed to be multifactorial in origin; however, impaired accumulation of drug is the single most commonly observed alteration when cells selected for resistance are compared with the sensitive cells from which they were derived (Andrews and Howell, 1990). The mechanisms by which DDP and CBDCA enter and exit from cells are not well characterized. Cellular accumulation of these drugs is relatively slow compared with many other classes of chemotherapeutic agents. Given their degree of hydrophilicity and polarity, transporters are likely to be required for both influx and efflux. Indeed, uptake is influenced by factors that suggest the involvement of both active and passive transporter-mediated processes (Gately and Howell, 1993).
One curious feature of cells with acquired DDP resistance is that they exhibit cross-resistance to a wide variety of metalloids, including arsenite (Naredi et al., 1995), antimony (Naredi et al., 1995; Chen et al., 1998), and cadmium (Lee et al., 1995). Recently, we reported that these cells are also cross-resistant to copper (Katano et al., 2002a), and that cells selected for acquired resistance to copper are cross-resistant to DDP (Safaei and Howell, 2001). Copper homeostasis is maintained by a complex system of transporters and chaperones that serve to both protect Cu(I) against oxidation and to prevent the production of toxic reactive oxygen species as the copper enters and is distributed throughout the cell (O'Halloran and Culotta, 2000). The central feature of this system is a group of proteins with unique metal binding sequences that chelate copper into protective pockets and exchange it through intimate protein-protein interactions such that copper is virtually never free in the cell (Pufahl et al., 1997; Rae et al., 1999). The primary uptake transporter for copper is hCTR1 (Zhou and Gitschier, 1997; Moller et al., 2000), and its function in mammals is essential for embryonic survival (Kuo et al., 2001; Lee et al., 2001). hCTR1 transfers copper to one of three known copper chaperones: Cox17, hCCS1, and HAH1. Cox17 delivers copper to cytochrome c oxidase, hCCS1 transfers copper to SOD1, and HAH1 hands it to one or another of the two P-type ATPases, ATP7A and ATP7B, that are reported to sequester copper into the trans-Golgi network where it is loaded onto copper-dependent enzymes in the secretory pathway and subsequently exported from the cell (Klomp et al., 1997). Mutations that disable ATP7A cause Menkes disease, and mutations that disable ATP7B cause Wilson's disease (Bull et al., 1993; Tanzi et al., 1993; Yamaguchi et al., 1993). ATP7A and ATP7B both have structural features common to other heavy metal P-type ATPases, including six metal binding sequence motifs in the N-terminal cytoplasmic domain (Solioz and Vulpe, 1996). Both molecules undergo copper-induced redistribution from the trans-Golgi network; ATP7A traffics to the plasma membrane (Petris et al., 1996; Cobbold et al., 2002), whereas ATP7B becomes relocalized to vesicular structures (Hung et al., 1997; Schaefer et al., 1999; Roelofsen et al., 2000; Vanderwerf et al., 2001).
A direct link between copper transport and DDP resistance has been identified by Komatsu et al. (2000) who found that prostate cancer cells selected for resistance to DDP expressed increased levels of ATP7B and that cells transfected with ATP7B were resistant to both copper and DDP. Increased levels of ATP7B mRNA or protein expression have been noted in several major human malignancies, including ovarian cancer (Nakayama et al., 2002), gastric carcinoma (Ohbu et al., 2003), breast cancer (Kanzaki et al., 2002), and various malignant cell lines (Nakayama et al., 2001). We have noted that cells selected for DDP resistance frequently overexpress either ATP7A or ATP7B (Katano et al., 2002b). To further investigate the role of ATP7B in platinum drug resistance, we molecularly engineered human carcinoma cell lines to overexpress ATP7B and examined them for changes in their sensitivity to CBDCA. We report here data demonstrating that ATP7B renders cells resistant to CBCDA as well as copper and DDP and that this is associated with parallel reductions in the cellular accumulation of both copper and CBDCA.
Materials and Methods
Drugs and Reagents. Cupric sulfate, CBDCA, and other chemicals were obtained from Sigma-Aldrich (St. Louis, MO) and Fisher Scientific Co. (Tustin, CA). DDP (PLATINOL-AQ) was a gift from Bristol Laboratories (Princeton, NJ). The rabbit polyclonal antiserum for ATP7B was generously provided by Dr. Jonathan D. Gitlin (Washington University, St. Louis, MO). Polyclonal antibody against actin was from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). Secondary antibodies were purchased from Amersham Biosciences (Piscataway, NJ). Protein concentration was measured using the Bio-Rad protein assay kit from Bio-Rad (Hercules, CA). Western blotting reagents were purchased from Bio-Rad. 64Cu was purchased from the Mallinkrodt Institute (St. Louis, MO), and [14C]CBDCA was purchased from Amersham Biosciences.
Cells and Vectors. The human ovarian carcinoma 2008 and IGROV-1 cell lines, and the human head and neck squamous carcinoma cell line UMSCC10b were grown in drug-free RPMI 1640 medium plus 10% fetal calf serum and maintained in humidified air containing 5% CO2 at 37°C. A pRC/CMV vector containing the full-length ATP7B cDNA and expressing a G-418 resistance marker, constructed as described previously (Terada et al., 1998; Harada et al., 2000), was generously provided by Dr. Toshihiro Sugiyama. Cells were transfected with pRc/CMV-ATP7B or empty vector with LipofectAMINE (Invitrogen, Carlsbad, CA) according to the manufacturer's directions. Transfected cells were selected in the presence of 500 μg/ml G-418, and all surviving clones were combined for form a multiclonal population. The 2008/ATP7B cells were further engineered to express GFP by infecting them with the amphotropic pMSCV-EGFP retrovirus generously provided by Dr. Martin Hass (Fink et al., 1998; Norris et al., 1998). Seven days after infection, the 5% of the population with the brightest green fluorescence was isolated by flow activated cell sorting and grown to mass culture in medium containing 500 μg/ml G-418 and 1 μg/ml puromycin.
Assays of Drug Sensitivity. Colony assays were performed using triplicate cultures of 200 cells/35-mm plate grown in 5 ml of medium containing different concentrations of DDP, CBDCA, or CuSO4 until visible colonies had formed (10–14 days). The dishes were rinsed twice with PBS, fixed with 100% methanol, and stained with a 0.1% crystal violet solution. A ChemiImager 400 instrument (Alpha Innotech, San Leandro, CA) was used for counting colonies of >50 cells. Enrichment assays were performed by preparing a population containing 10% 2008/ATP7B-GFP cells and 90% 2008/EV cells, seeding 105 cells into 100-mm culture dishes, incubating them together for 24 h, and then exposing them to graded concentrations of DDP, CBDCA, or copper for 1 h. Fresh non–drug-containing medium was then added and the cells grown for 5 days before being harvested by trypsinization and analyzed for the percentage of 2008/ATP7B-GFP cells in the surviving population by flow cytometry. Each assay was performed with triplicate cultures.
Cellular Pharmacokinetic Assays. Uptake and efflux measurements were made using 35-mm dishes seeded with 106 cells each and incubated in medium until they were 75 to 80% confluent. For cellular accumulation experiments, the medium was replaced by 1 ml of fresh medium containing 2 μM 64CuSO4 or 50 μM[14C]CBDCA and the cells were incubated at 37°C. Copper and CBDCA concentrations were selected based on prior reports indicating that alterations in the cellular pharmacokinetic parameters could be detected at these concentrations (Shen et al., 2000; Lee et al., 2002b). Efflux rates were measured by exposing the cells to 2 μM 64CuSO4 or 50 μM [14C]CBDCA for 1 h, rinsing them with fresh medium, and incubating them in drug-free medium at 37°C. At the requisite time points in both types of experiments, the medium was poured off and the dishes were quickly rinsed three times with ice-cold PBS after which the cells were harvested using a plastic scraper and transferred to scintillation vials containing 3 ml of scintillation solution (National Diagnostics, Atlanta, GA). 64Cu and [14C]CBDCA were quantified by scintillation counting. Six separate dishes were used for each time point in each experiment. Cells harvested from a separate group of six dishes were used to measure protein content by the Bradford assay.
Measurement of Basal Copper Content. Cultures were quickly rinsed three times with ice-cold PBS, and cells were harvested into 15 ml of ice-cold PBS using a rubber policeman. After centrifugation at 3000 rpm for 10 min, the cells were resuspended in PBS, an aliquot was used for protein assay, and the remainder was digested in 68% nitric acid. Cell lysates were heated for 2 h at 65°C diluted to 5% nitric acid and assayed for platinum and copper content. An inductively coupled plasma optical emission spectroscopy apparatus (model 3000DV; PerkinElmer Life Sciences) at the Analytical Facility at the Scripps Institute of Oceanography was used for copper and platinum assays.
Pharmacokinetic Analysis. Mean data were fitted using a two-compartment pharmacokinetic model assuming a first order disposition process using WinNonlin Professional 3.1 (Pharsight, Mountain View, CA).
Western Blotting. Cells were rinsed twice with PBS, scraped from the dish in PBS, and centrifuged for 10 min at 2500 rpm. Cells were lysed in 0.25% Nonidet P-40 in 100 mM Tris HCl, pH 8, supplemented with 1 mM p-amidinophenylmethylsulfonyl fluoride hydrochloride and 1 mM γ-amino-n-caproic acid (Sigma-Aldrich) at 4°C and for 30 min. Postnuclear fractions were obtained by centrifugation of cell lysates for 10 min at 600g. Samples containing 50 to 100 μg of protein were electrophoresed on 4 to 10% SDS polyacrylamide gels and then blotted onto nitrocellulose filters using a Bio-Rad Mini Transblot apparatus (Bio-Rad). Blots were incubated for 1 h with 5% fat-free dry milk in TBS at room temperature and then overnight in primary antibody against ATP7B from Dr. Jonathan D. Gitlin mixed with 5% milk in TBS at 4°C at a dilution of 1:3000. Blots were washed three times for 15 min each at room temperature with 0.025% Tween 20 in TBS. The secondary antibody was diluted 1:1000 in 5% fat-free dry milk in TBS and added to blots for 1 h at room temperature. Blots were washed again at room temperature three times for 15 min each in 0.025% Tween 20 in TBS. The extent of specific staining was quantified by chemiluminescence using the ECL kit from Amersham Biosciences Inc. The membrane was immediately exposed to Fuji medical X-ray film (Super RX; Fujifilm, Kanagawa, Japan). A ChemiImager 400 instrument (Alpha Innotech) was used for determining the relative density of protein bands.
Statistics. Tests of significance used Student's t test; p values of <0.05 were considered significant.
Results
ATP7B Confers Resistance to Copper, DDP, and CBDCA. Three different cell lines sensitive to both DDP and CBDCA were used in these experiments, including the human ovarian carcinoma cell lines 2008 and IGROV-1 and the human head and neck squamous cell carcinoma UMSCC10b. Each parental line was transfected with either a vector that constitutively expresses ATP7B from a CMV promoter or with a control vector containing no insert. Both vectors contained a marker for resistance to G-418. All clones that grew in the presence of G-418 were pooled to form a population that was then tested by Western blot analysis for its relative level of ATP7B using a polyclonal antibody directed at the N-terminal domain of ATP7B. Figure 1 shows that the 170-kDa ATP7B protein was present in all three ATP7B-transfected populations but was not detected in any of the parental cell populations.

Western blot analysis of ATP7B expression in empty vector and ATP7B-expressing vector transfected cells. The same blots were probed with antibody to actin to document equal lane loading.
The sensitivity of the empty vector- and ATP7B vector-transfected populations to DDP, CBDCA, and CuSO4 was tested in clonogenic assays using a 1-h drug exposure. The concentration-survival curves are shown in Fig. 2, and the IC50 values determined from these curves are summarized in Table 1. The expression of ATP7B conferred moderate but significant degrees of resistance to all three compounds in all three ATP7B-expressing cell populations. Of note is the fact that ATP7B expression conferred a greater degree of resistance to DDP than to copper in each of the three pairs of cell lines tested.

Clonogenic survival of cells control and ATP7B-expressing cells as a function of DDP, CBDCA, or CuSO4 concentration. ▪, cells transfected with empty vector; □, cells transfected with ATP7B-expressing vector. A to C, IGROV-1/EV compared with IGROV-1/ATP7B; D to F, 2008/EV compared with 2008/ATP7B; G to I, UMSCC10b/EV compared with UMSCC10b/ATP7B. Each data point represents the mean of three independent experiments each performed with triplicate cultures; vertical bars are ± S.E.M.
Sensitivity of human cancer cell lines to a 1-h exposure to DDP, CBDCA, and CuSO4 IC50 values are presented as mean ± S.D.
The effect of low level ATP7B expression on sensitivity to DDP, CBDCA, and copper was further examined in the 2008/EV and 2008/ATP7B cell pair using an assay capable of detecting enrichment for ATP7B-expressing cells in the population that survives drug exposure. The 2008/ATP7B cells were engineered to express GFP by infection with a viral vector. The 2008/EV and 2008/ATP7B-GFP cells were then mixed to form a population containing 10% 2008/ATP7B-GFP cells as determined before drug exposure by flow cytometric analysis. The mixed population was then exposed to increasing concentration of DDP, CBDCA, or copper for 1 h and then cultured for 5 days before the fraction of 2008/ATP7B-GFP cells was again determined by flow cytometry. This assay has the advantage that both the control and ATP7B-expressing cells were exposed to identical conditions in the same culture throughout the experiment (Torrance et al., 2001). The results of these experiments, presented in Fig. 3, demonstrated that in the absence of drug exposure, the fraction of 2008/ATP7B cells did not change significantly during the 5 days of culture. However, all three drugs produced very substantial degrees of enrichment for ATP7B-expressing cells, even at the lowest concentration of drug tested. These results confirm that the level ATP7B expression present in the population of 2008/ATP7B cells was sufficient to confer a biologically important degree of resistance.
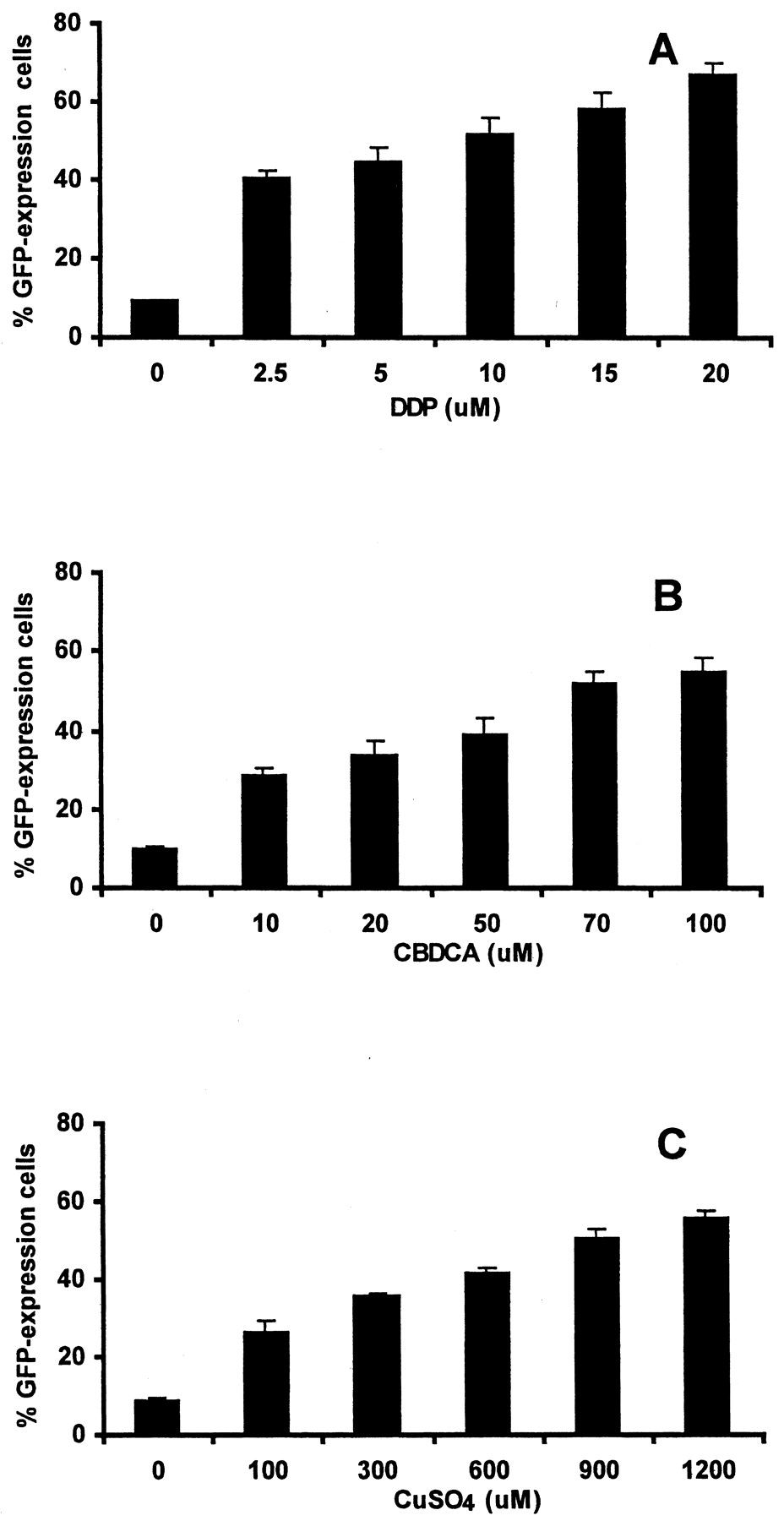
Enrichment for ATP7B-expressing cells as a function of DDP, CBDCA, and copper concentration. Each column indicates the mean percentage of 2008/ATP7B cells in the surviving population 5 days after exposure of a starting population containing 10% 2008/ATP7B and 90% 2008/EV cells to the indicated concentration of drug for 1 h. The 2008/ATP7B cells were engineered to express GFP. Each column represents the mean of three independent experiments; vertical bars, ± S.E.M.
ATP7B Alters Copper and CBDCA Steady-State Accumulation. Studies of the cellular pharmacology of DDP at physiologically relevant concentrations are problematic because of the lack of a readily available radiolabeled form of the drug and the limited sensitivity of alternative analytic approaches. Thus, we focused on the effect of ATP7B expression on the cellular accumulation of copper, which could be measured by using 64Cu, and CBDCA, for which 14C-labeled drug was available. Studies of the accumulation of copper and CBDCA were carried out in the 2008/EV and 2008/ATP7B pair of cell lines. The steady-state level of copper in the cell is quite sensitive to the activity of ATP7B (La Fontaine et al., 1998). Figure 4A shows the levels of copper in the 2008/EV and 2008/ATP7B cells when grown in the tissue culture medium to which no copper had been added, and Fig. 4B shows the levels when grown in medium containing 2 μM copper for 24 h. Steady-state accumulation was reached for both copper by 24 h (data not shown). The basal content of copper in the 2008/EV cells was 1.3-fold higher than that in the 2008/ATP7B cells (p < 0.05). The copper content of the 2008/EV cells was 1.9-fold higher than that of the 2008/ATP7B cells when grown in the copper-supplemented medium (p = 0.00002). Figure 4C shows the level of CBDCA in 2008/EV and 2008/ATP7B cells exposed to 50 μM CBDCA for 24 h, at which time steady state had been achieved. The CBCDA content was 2.8-fold higher in the 2008/EV than in the 2008/ATP7B cells (p = 0.00005). Thus, the increased expression of ATP7B produced a reduction in the accumulation of both copper and CBDCA under steady-state conditions.

Basal and steady-state cellular content of copper and platinum. A, basal content of copper in cells growing in medium without any added copper as measured by inductively coupled plasma optical emission spectroscopy apparatus. B, Copper content of cells grown in medium containing 2 μM 64Cu for 24 h. C, CBCDA content of cells grown in medium containing 50 μM [14C]CBDCA for 24 h. Each histogram represents the mean of three independent experiments each performed with three (CBDCA) or six (copper) separate cultures. Vertical bars, ± S.E.M.
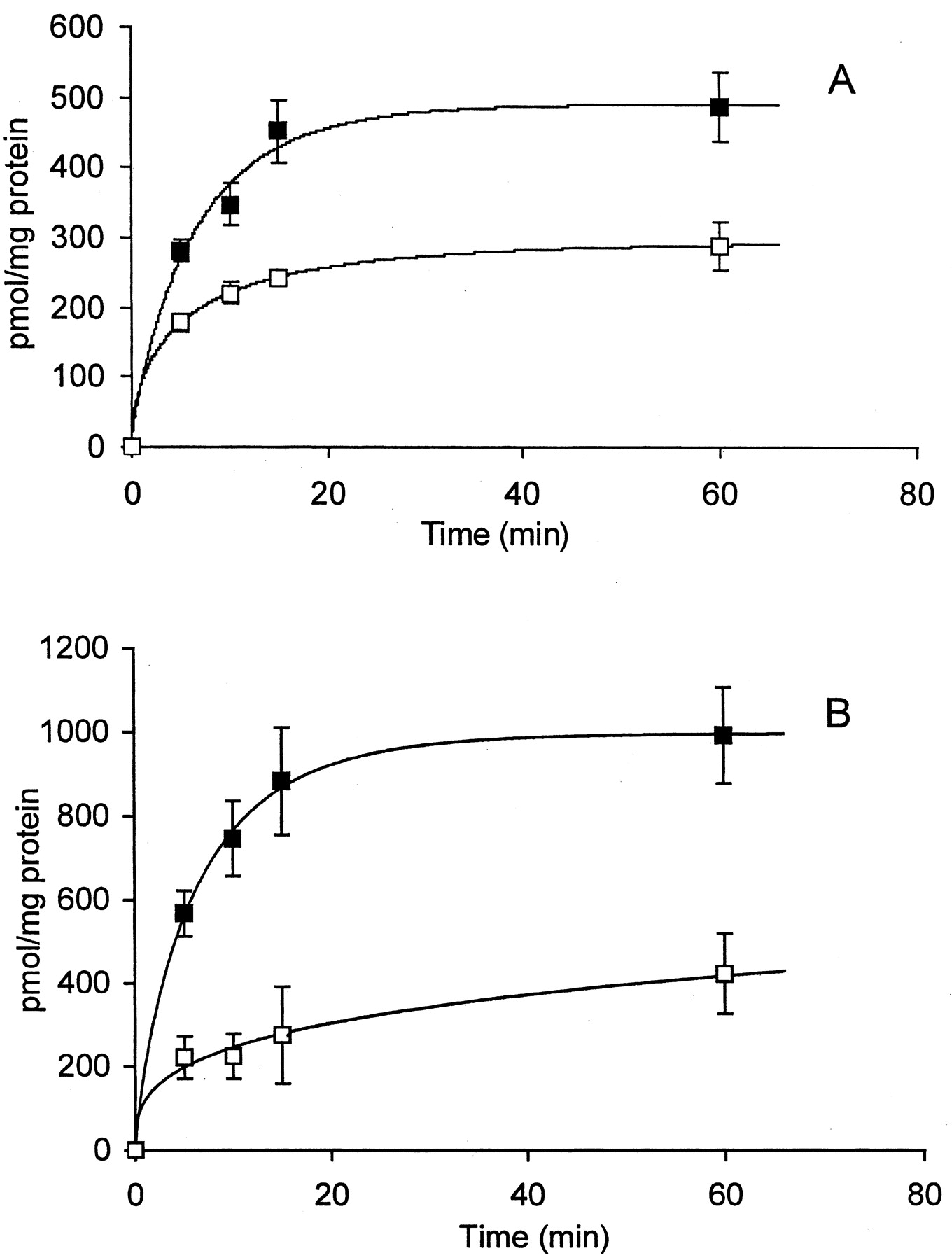
ATP7B Alters Copper and CBDCA Uptake and Efflux. The decreased steady-state accumulation of copper and DDP in the 2008/ATP7B cells must reflect either reduced influx, decreased intracellular binding, increased efflux, or some combination of all three effects. Figure 5 compares the time course of copper and CBDCA accumulation in the 2008/EV and 2008/ATP7B cells over the first hour of exposure. For both copper and CBDCA the extent of accumulation over the first hour was substantially less in the 2008/ATP7B cells than in the 2008/EV cells. Thus, expression of ATP7B markedly reduced the rate of accumulation of copper and CBDCA over this time in a parallel manner.

Time course of accumulation of copper (A) and CBDCA (B) in 2008/EV (▪) and 2008/ATP7B cells (□). Each data point represents the mean of measurements made on three (CBDCA) or six (copper) separate cultures per time point; vertical bars, ± S.E.M.
Accumulation at 1 h can still reflect differences in intracellular binding and efflux as well as initial influx rate. To determine the relative contribution of changes in influx, ideally one would like to measure the initial influx rate over a very short period. However, copper and CBDCA both enter cells relatively slowly, and the shortest period over which the influx of copper and CBDCA could be accurately measured was 5 min. Over this period, the average rate of accumulation of copper in the 2008/EV cells was 56 ± 7 (S.E.M.) pmol/mg of protein/min, whereas in the 2008/ATP7B cells the rate was reduced by 37% to 36 ± 6 (S.E.M.) (p = 0.0007). The rate of accumulation of CBDCA was 113 ± 11 (S.E.M.) pmol/mg of protein/min in the 2008/EV cells, whereas it was reduced by 61% to 44 ± 10 (S.E.M.) pmol/mg of protein/min in the 2008/ATP7B cells (p = 0.0009). Thus, when measured over as short an interval as possible, increased expression of ATP7B resulted in a clear decrease in cellular accumulation for both copper and CBDCA.
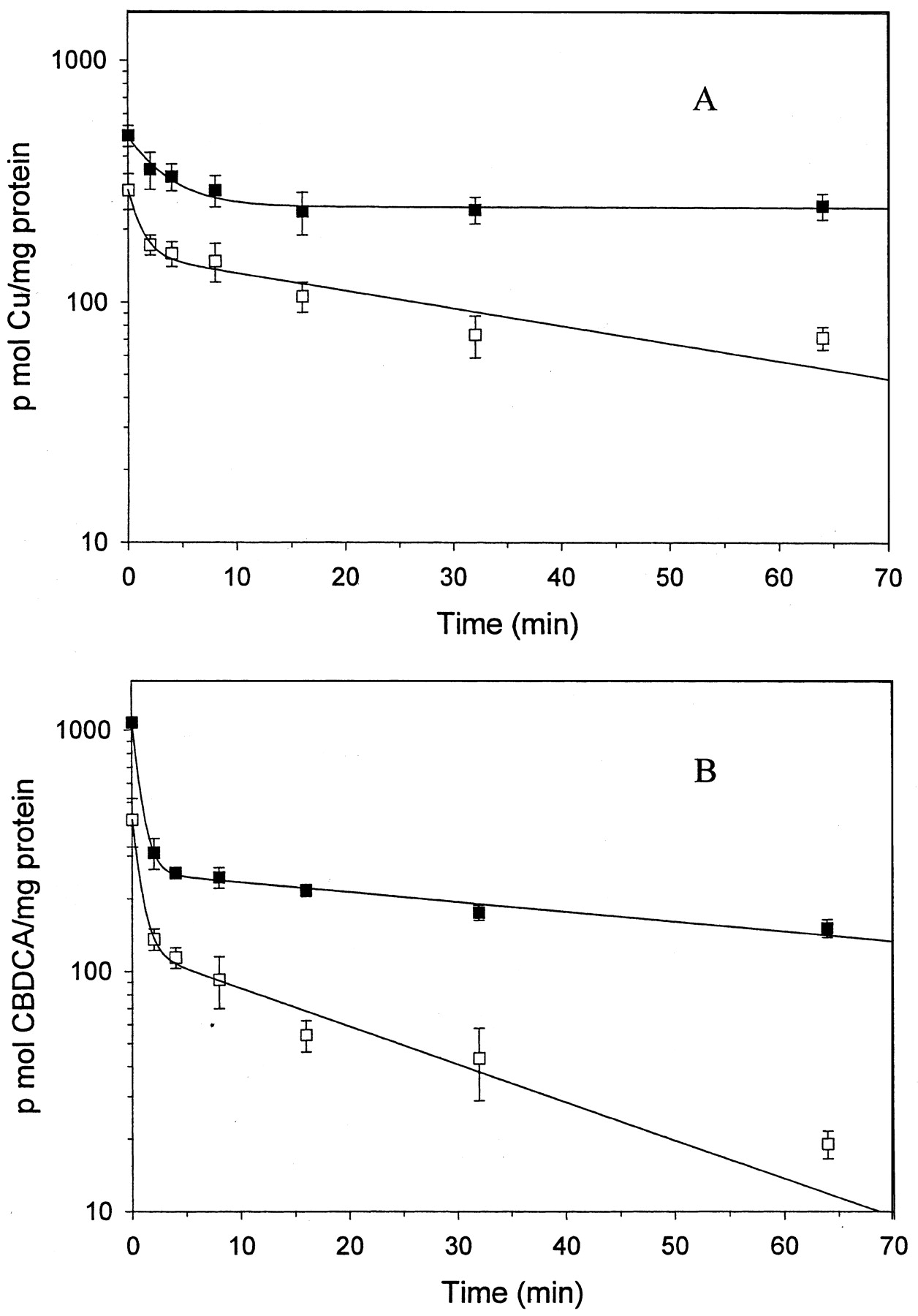
To examine the contribution of changes in efflux to the reduced accumulation the 2008/EV and 2008/ATP7B cells were loaded by exposure to either 2 μM copper or 50 μM CBDCA for 1 h. At each of multiple time points after removal of the drugs, six cultures were harvested using a rapid sampling technique. Figure 6 shows the efflux curves over the first hour after the end of drug exposure. For both copper and CBDCA, the efflux curves were characterized by an initial rapid phase, extending over approximately the first 5 min, followed by a much slower phase. Inspection of the curves indicates that increasing the level of ATP7B expression increased the efflux for both copper and CBDCA. The data were fitted to a two-compartment model, and the estimated half-lives for the initial and terminal phases of efflux are presented in Table 2. For copper, the initial phase of efflux was approximately 3-fold more rapid from the 2008/ATP7B than from the 2008/EV cells. Because the second phase of copper efflux from the 2008/EV cells was so slow, a half-life for this phase could not be accurately estimated but it was clearly shorter in the 2008/ATP7B cells. For CBDCA two-compartment modeling yielded similar half-lives for the initial phase of efflux from the 2008/EV and 2008/ATP7B cells, but efflux during the second phase was 3.9-fold faster from the 2008/ATP7B cells. Thus, increased expression of ATP7B increased at least the second phase of efflux for both copper and CBDCA in a parallel manner and to a substantial degree. It is important to note that the absolute efflux rates cannot be directly compared between copper and CBDCA because the extent of intracellular binding of these two metalloids is not known.

Copper (A) and CBDCA (B) efflux from cells preloaded with drug for 60 min. ▪, 2008/EV cell line; □, 2008/ATP7B cell line. Each data point represents the mean of measurements made on three (CBDCA) or six (copper) separate cultures per time point; vertical bars, ± S.E.M.
Efflux half-lives of copper and CBDCA estimated from two-compartment analysis
Discussion
The results reported here provide evidence that the expression of ATP7B in human carcinoma cells modulates sensitivity to both of the major clinically used platinum-containing chemotherapeutic agents DDP and CBDCA, as well as to copper. In three different human cell lines, expression of ATP7B from a mammalian promoter resulted in a modest increase in ATP7B protein levels in the population of transfected cells, and this was accompanied by a statistically significant degree of resistance to each of the three compounds when assessed using colony formation assays. When measured with this type of assay, these changes constitute significant levels of resistance to DDP and CBDCA, certainly high enough that one would expect differences in in vivo responsiveness (Andrews et al., 1990). For the 2008/EV and 2008/ATP7B cell pair, the results of the colony formation assays were confirmed by enrichment assays that demonstrated a large increase in the ATP7B-expressing cells in the population that survived drug exposure. It is noteworthy that the magnitude of the change in IC50 value was greater for DDP than for copper in each of the three cell lines. The reason for this is not known, but it is consistent with the results of our prior studies of cross-resistance between DDP and copper in cells selected for resistance to either DDP (Katano et al., 2002a) or copper (Safaei and Howell, 2001), and the results reported by Komatsu et al. (2000) in epidermoid cells transfected with ATP7B. In all of these systems, relatively modest degrees of resistance to copper were associated with significantly higher degrees of resistance to DDP.
The cellular pharmacology studies in the 2008/EV and 2008/ATP7B pair of cell lines provide evidence that the mechanism by which ATP7B expression renders cells resistant to copper and CBDCA is through reduction in cellular drug accumulation. The ATP7B-expressing cells exhibited reduced steady-state levels of copper when grown in regular tissue culture medium, and when grown in medium supplemented with 2 μM CuSO4. The same effect was observed when the cells were exposed to CBDCA for 24 h, and the magnitude of the effect on CBDCA steady-state levels was even greater than for copper. The diminished steady-state levels must reflect either decreased influx, reduced cellular binding, or enhanced efflux. Compared with many other types of compounds, the influx of copper and CBDCA is relatively slow, making it difficult to obtain accurate measurements of initial influx rate even with the use of radioactive forms of the compounds. However, the time course of uptake over the first hour of copper or CBDCA exposure suggests that measurements of accumulation at 5 min provide a useful estimate of the initial influx rate, and such measurements have been used by others for this purpose (Lee et al., 2002a; Puig et al., 2002). Given the evidence that ATP7B functions primarily in copper export, it is of particular interest that increased expression of ATP7B reduced the average initial accumulation rate by 37% for copper and by fully 61% for CBDCA. This may be explained in part by the fact that the initial efflux rate for both copper and CBDCA is quite rapid. Two-compartment modeling yielded an estimate for the initial efflux half-lives from the ATP7B-expressing cells that were short enough that concurrent drug export may have operated to reduce the apparent initial cellular accumulation rate. Alternatively, high levels of ATP7B may sequester copper chaperones needed for optimal inwardly directed transfer of copper. CTR1 is thought to hand copper to ATOX1 for further transfer to ATP7B (Hamza et al., 2001). ATOX1 binds to ATP7B (Walker et al., 2002) and high levels of ATP7B may limit the availability of ATOX1 for partnering with CTR1. We (Lin et al., 2002) have confirmed the observation made by Ishida et al. (2002) that loss of CTR1 function in Saccharomyces cerevisiae markedly reduces the uptake of DDP and further demonstrated that CTR1 modulates the uptake of CBDCA, oxaliplatin, and other DDP analogs as well as copper. Thus, if overexpression of ATP7B indirectly impairs CTR1 transport capacity, it would be reasonable to expect it to reduce DDP and CBDCA influx as well.
Increased expression of ATP7B was associated with more rapid efflux of both copper and CBDCA from the cell. For both compounds the efflux curve was characterized by two phases; the first phase was very much more rapid than the second. For both copper and CBDCA, efflux during the second phase was substantially faster from the 2008/ATP7B cells than from the 2008/EV cells. An accurate estimate of the magnitude of this change could not be developed for copper because the efflux half-life for the 2008/EV cells was so long. However, in the case of CBDCA this phase of efflux was 3.9-fold faster from the 2008/ATP7B cells. Interestingly, two-compartment modeling suggested that although increased expression of ATP7B produced a 2.9-fold increase in initial efflux of copper, it had little impact on the initial efflux of CBDCA. This may be related to differences in the intracellular binding of these two compounds and/or their positioning in cellular subcompartments relative to that of ATP7B. Although it is appropriate to compare the 2008/EV and 2008/ATP7B cells with respect to efflux of copper and CBDCA, the inability to assess free intracellular levels limits the validity of comparing the absolute rate of efflux of copper to that of CBDCA.
A number of questions about the transport of DDP and CBDCA by ATP7B remain to be addressed. Given the ability of ATP7B to render cells resistant to DDP as well as CBDCA, one might expect ATP7B to alter the initial cellular accumulation and enhance the initial efflux of DDP and oxaliplatin as well as that of CBDCA. Komatsu et al. (2000) noted reduced accumulation and retention of DDP in epidermoid cells transfected with ATP7B, but detailed cellular pharmacokinetic studies of DDP have yet to be presented. The export of copper is thought to proceed first through sequestration into the trans-Golgi and subsequently via vesicle migration to the cell surface (Walker et al., 2002). It remains to be determined whether the platinum-containing drugs are similarly sequestered into vesicles of the secretory pathway. ATP7B is found predominantly in the trans-Golgi network or a closely related endosomal compartment. However, it has been reported that copper induces redistribution of ATP7B to other areas of the cell (Hung et al., 1997; Schaefer et al., 1999; Roelofsen et al., 2000; Vanderwerf et al., 2001). Whether the platinum-containing drugs cause a similar redistribution is the focus on ongoing studies. How large a change in ATP7B levels is required to produce clinically significant degrees of resistance to DDP and CBDCA is a remaining question of substantial clinical significance.
The demonstration that increased expression of ATP7B produces DDP and CBDCA resistance in several different cell systems and that it does so by modulating the cellular pharmacology of these drugs provides a solid foundation for investigation of the expression of this transporter in human tumors known to be intrinsically sensitive or resistant to the platinum-containing drugs. Together with evidence that the major copper influx transporter CTR1 mediates the uptake of DDP (Ishida et al., 2002; Lin et al., 2002), this suggests the unifying concept that DDP and CBDCA enter the cell, are distributed to subcellular locations, and are exported from the cell in part using the transporters and chaperones that evolved primarily to mediate copper homeostasis.
Acknowledgments
We thank Drs. Toshihiro Sugiyama, Michael Petris, Jonathan D. Gitlin, and Martin Haas for providing essential vectors and reagents, W. Naerdemann for excellent technical assistance, and C. Zacharia for project management assistance.
Footnotes
-
This study was supported in part by National Institutes of Health grant CA78648. This work was conducted in part by the Clayton Foundation for Research-California Division. R.S. and S.B.H. are Clayton Foundation investigators.
-
ABBREVIATIONS: DDP, cisplatin; CBDCA, carboplatin; GFP, green fluorescent protein; PBS, phosphate-buffered saline; TBS, Tris-buffered saline; CMV, cytomegalovirus.
- Received January 21, 2003.
- Accepted April 30, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}