Article Text

Abstract
BACKGROUND/AIMS Hepatocellular carcinoma (HCC) is a common malignant tumour worldwide, and its differential diagnosis from benign lesions of the liver is often difficult yet of great clinical importance. In the present study, we analysed whether glypican-3 is useful in differentiating between benign and malignant liver diseases and whether it influences the growth behaviour of HCC.
METHODS Northern blot analysis and in situ hybridisation.
RESULTS Northern blot analysis indicated that expression of glypican-3 mRNA was either low or absent in normal liver, in focal nodular hyperplasia (FNH), and in liver cirrhosis. In contrast, expression of glypican-3 mRNA was markedly increased in 20 of 30 and moderately increased in five of 30 HCC samples. The average increase in glypican-3 mRNA expression in HCC was significant compared with expression in normal liver (21.7-fold increase, p<0.01). In comparison with FNH or liver cirrhosis, glypican-3 mRNA expression in HCC was increased 7.2- (p<0.05) and 10.8-fold (p<0.01), respectively. In addition, pushing HCCs exhibited significantly higher glypican-3 mRNA expression than invading tumours (p<0.05). In situ hybridisation analysis demonstrated weak expression of glypican-3 mRNA in normal hepatocytes and bile ductular cells, and weak to occasionally moderate signals in hepatocytes forming nodules of liver cirrhosis and in regenerated hepatic nodules of FNH. In contrast, glypican-3 in situ hybridisation signals were intense in hepatic cancer cells with even higher levels in pushing HCCs than in invading HCCs.
CONCLUSIONS These findings suggest that glypican-3, in many cases, has the potential to differentiate between benign and malignant liver diseases.
- focal nodular hyperplasia
- liver cirrhosis
- hepatocellular cancer
- glypican-3
Abbreviations used in this paper
- HCC
- hepatocellular carcinoma
- FNH
- focal nodular hyperplasia
- HSPGs
- heparansulphate proteoglycans
- HGF
- hepatocyte growth factor
- EGF
- epidermal growth factor
- HB-EGF
- heparin binding EGF-like growth factor
- SDS
- sodium dodecyl sulphate
Statistics from Altmetric.com
Hepatocellular carcinoma (HCC) is one of the most severe sequelae of chronic liver disease. In spite of recent therapeutic advances, this malignancy continues to be a significant cause of cancer related morbidity and mortality in Asian and Western countries.1 Surgical resection remains the treatment of choice for these tumours. Unfortunately, only 10–20% of primary liver tumours are found to be resectable at the time of diagnosis.2 Advances in radiological imaging over the past two decades have focused attention on early detection of hepatic nodular lesions. Various pathological entities with a nodular appearance are predominantly composed of hepatocytes or tumour cells of hepatocytic origin, including benign and malignant neoplasms as well as tumour-like lesions such as focal nodular hyperplasia (FNH). The differential diagnosis of these nodules is often difficult, especially in view of the limited material which is obtained by needle biopsy.3 FNH and liver cirrhosis are the most common benign liver diseases, which may present with a nodular appearance. Differentiating between focal nodules in FNH or regenerative nodules in cirrhotic livers and malignant nodules is of great clinical importance4 because of the resulting therapeutic consequences. However, differentiation between benign and malignant nodules, especially of the nodules of liver cirrhosis, is often difficult and even the sensitive and specific magnetic resonance imaging has its limitations in these cases.
Glypican-3 belongs to the glypican family of GPI anchored heparansulphate proteoglycans (HSPGs) which plays an important role in cellular growth, cell differentiation, and cell migration. To date, six distinct members of the human glypican family have been identified, and their importance in various diseases has recently been recognised.5 Mutations in the glypican-3 gene are responsible for the Simpson-Golabi-Behmel syndrome in humans which is characterised by pre- and postnatal overgrowth and by a number of other abnormalities.6 ,7 In addition, glypicans seem to influence tumour pathogenesis in various human malignancies.8-10 For example, in mesotheliomas, induction of glypican-3 expression inhibits cell growth, and glypican-3 mRNA levels are significantly reduced in human malignant mesothelioma tumours and cell lines.8 ,10 In addition, in MCF-7 breast cancer cells, enhanced glypican-3 expression leads to apoptosis.8 Glypican-3 expression is lost in some ovarian cancer cell lines, and restoration of glypican-3 expression inhibits the growth of these cells, suggesting that glypican-3 may function as a tumour suppressor in ovarian cancer.9 Recent studies have shown that a variety of growth factors and their receptors, such as hepatocyte growth factor (HGF), c-met, epidermal growth factor (EGF), and heparin binding EGF-like growth factor (HB-EGF) are upregulated in HCC.11-13 Inasmuch as HGF and HB-EGF require membrane bound HSPGs as a co-receptor for their signalling, the presence of glypican may regulate the growth behaviour of HCC. Furthermore, there is also increasing evidence that HSPGs, such as syndecan-1, -2, -3, or -4, may play an important role in the normal liver, in chronic liver disease, and in the carcinogenesis of primary liver tumours.14-16 Furthermore, preliminary results indicate that glypican-3 is differentially expressed in HCC compared with the normal liver and that it may serve as a tumour marker in this malignancy.17
In the present study, we evaluated expression of glypican-3 mRNA in two morphological subtypes of HCC—pushing and invading tumours, as described previously18 —compared with normal liver tissues as well as with the common benign liver diseases FNH and liver cirrhosis. We now report that glypican-3 is highly expressed in many hepatic cancers but not in benign hepatic disorders, and that there are differences in its expression between pushing and invading tumours. These data suggest that glypican-3 may be useful in differentiating between HCC and benign hepatic nodules and that its presence may influence the growth characteristics of HCCs.
Materials and methods
HUMAN LIVER SAMPLES
Normal human liver tissue samples (10 female and five male donors; median age 53 years; range 28–56) were obtained from previously healthy organ donors (n=9) and from patients undergoing hemihepatectomy due to colorectal metastasis (n=6). In the case of liver resection, tissue specimens were taken at the farthest distance from the metastasis. Histological analysis of the liver specimens revealed normal liver tissue in all cases. FNH tissues were obtained from three male and four female patients (median age 58 years; range 39–68) who underwent liver resection due to a growing nodular hepatic tumour. Liver cirrhosis tissue samples were obtained from 28 patients (20 males and eight female; median age 63 years; range 32–72), 21 of whom underwent liver transplantation (eight hepatitis B, 10 hepatitis C, and three alcoholic) and seven of whom underwent surgery and hepatic exploration/resection due to unclear hepatic nodules which were identified as liver cirrhosis by histopathological examination. HCCs were obtained from 30 patients (20 males and 10 female; median age 64 years; range 33–78) undergoing liver resection. According to the TNM (tumour-node-metastasis) classification and histopathological grading of the UICC (Union Internationale Contre le Cancer),19there were no stage I tumours, two stage II tumours, 14 stage III tumours, and 14 stage IV tumours. Tumour grading showed four well differentiated tumours (grade 1), 14 moderately differentiated tumours (grade 2), 11 poorly differentiated tumours (grade 3), and one undifferentiated tumour (grade 4). Nineteen tumours were pushing carcinomas and 11 were invading tumours (table 1), which were classified as previously described.18 Freshly removed tissue samples were fixed in 5% formaldehyde solution for 12–24 hours and paraffin embedded for histological analysis and in situ hybridisation. In addition, tissue samples were frozen in liquid nitrogen immediately on surgical removal and maintained at −80°C until use for RNA extraction. All studies were approved by the ethics committee of the University of Bern.
Glypican-3 mRNA expression in patients with hepatocellular carcinoma
NORTHERN BLOT ANALYSIS
Total RNA was extracted using the guanidinium isothiocyanate method20 ,21 followed by electrophoresis under denaturing conditions in a 1.2% agarose/1.8 M formaldehyde gel.20 ,21 RNA was electrotransferred onto nylon membranes (Gene Screen, Du Pont, Boston, Massachusetts, USA) and cross linked by UV irradiation. The filters were then prehybridised, hybridised, and washed under high stringency conditions. Prehybridisation was performed overnight at 65°C in 50% formamide, 0.5% sodium dodecyl sulphate (SDS), 5× SSC, 5× Denhardt's solution (1×Denhardt's=0.02% ficoll, 0.02% polyvinylpyrrolidone, and 0.02% bovine serum albumin), 250 μg/ml salmon sperm DNA, and 50 mM sodium phosphate (pH 6.5). The blots were then hybridised for 18 hours at 65°C in the presence of 1×106 cpm/ml of the 32P labelled antisense glypican-3 cRNA probe, washed twice at 65°C in 1× SSC and 0.5% SDS, and washed twice at 65°C in 0.1× SSC and 0.5% SDS.
All blots were rehybridised with a human 32P labelled 7S cDNA probe to assess equivalent RNA loading and transfer.20 ,21 The blots were prehybridised overnight at 42°C in a buffer containing 50% formamide, 1% SDS, 0.75 M NaCl, 5mM EDTA, 5× Denhardt's solution, 100 μg/ml salmon sperm DNA, 10% dextran sulphate, and 50 mM sodium phosphate (pH 7.4). Following hybridisation with 1×105 cpm/ml of the 32P labelled 7S cDNA probe, membranes were rinsed twice at 50°C in 2× SSC and washed three times at 55°C in 0.2× SSC and 2% SDS.
All blots were exposed at −80°C to Fujix ray film with Kodak intensifying screens for 1–10 days. The intensity of the radiographic bands was quantified by a computerised video system and the Image-pro-plus 3.0 software (Media Cybernetics, Silver Spring, Maryland, USA). The ratios of the optical densities of the RNA levels (glypican-3/7S) were calculated for each sample.
IN SITU HYBRIDISATION
In situ hybridisation was performed as previously reported.20 ,21 Briefly, 4 μm formaldehyde fixed, paraffin embedded tissue sections were post-fixed with 4% paraformaldehyde in phosphate buffered saline for five minutes, and incubated with 0.2 M HCl for 20 minutes. The samples were prehybridised at 50°C for at least one hour in 50% formamide (v/v), 4× SSC, 2× Denhardt's solution, and 250 μg RNA/ml. Hybridisation was performed overnight at 50°C in 50% (v/v) formamide, 4× SSC, 2× Denhardt's solution, 500 μg RNA/ml, and 10% dextran sulphate (w/v). The final concentration of the digoxigenin labelled glypican-3 probes (antisense and sense) was approximately 0.5 ng/μl. After hybridisation, sections were washed and treated with RNase (Roche Diagnostics, Rotkreuz, Switzerland). The samples were then incubated with an antidigoxigenin antibody conjugated with alkaline phosphatase (dilution 1:500; Roche Diagnostics). For colour reaction, 5-bromo-4-chloro-3-indolyl phosphate and nitroblue tetrazolium (Sigma, Buchs, Switzerland) were used. For control experiments, the slides were incubated with RNase or with the corresponding sense probes. Pretreatment of the slides with RNase abolished the hybridisation signal produced by the antisense probe. Furthermore, incubation with the sense probe failed to produce any specific signals.
PREPARATION OF cRNA AND cDNA PROBES
A 239 bp fragment (base pairs 927–1165, Genbank accession number: HSV50410) of the human glypican-3 cDNA was subcloned into the pBSIISK+ vector carrying promoters for the DNA dependent T3 and T7 RNA polymerases.21 The glypican-1, glypican-4, and glypican-6 cDNA probes were generated as described previously.21 A 190 bp fragment of mouse 7S that cross hybridises with human 7S was used to verify equivalent RNA loading in northern blot analysis, as described previously.20 ,21
For northern blot analysis, the glypican-3 antisense cRNA probe was radiolabelled with 32P CTP (Du Pont, International, Regensdorf, Switzerland) using an in vitro transcription system (Promega Biotechnology, Madison, Wisconsin, USA), and the 7S cDNA, and the other glypican probes were labelled with 32P dCTP (Du Pont) using a random primer labelling system (Roche Diagnostics), as described previously.21
For in situ hybridisation, the glypican-3 cRNA probe was labelled with digoxigenin. After linearisation, the glypican-3 cDNA was transcribed using the Ribomax System (Promega Biotechnology). The transcription resulted in digoxigenin labelled sense and antisense riboprobes specific for glypican-3 mRNA.20 ,21
STATISTICAL ANALYSIS
Results are expressed as median (range) or mean (SEM). For statistical analysis the Mann-Whitney U test, Student'st test, and the Spearman correlation test were used. Significance was defined as p<0.05.
Results
NORTHERN BLOT ANALYSIS
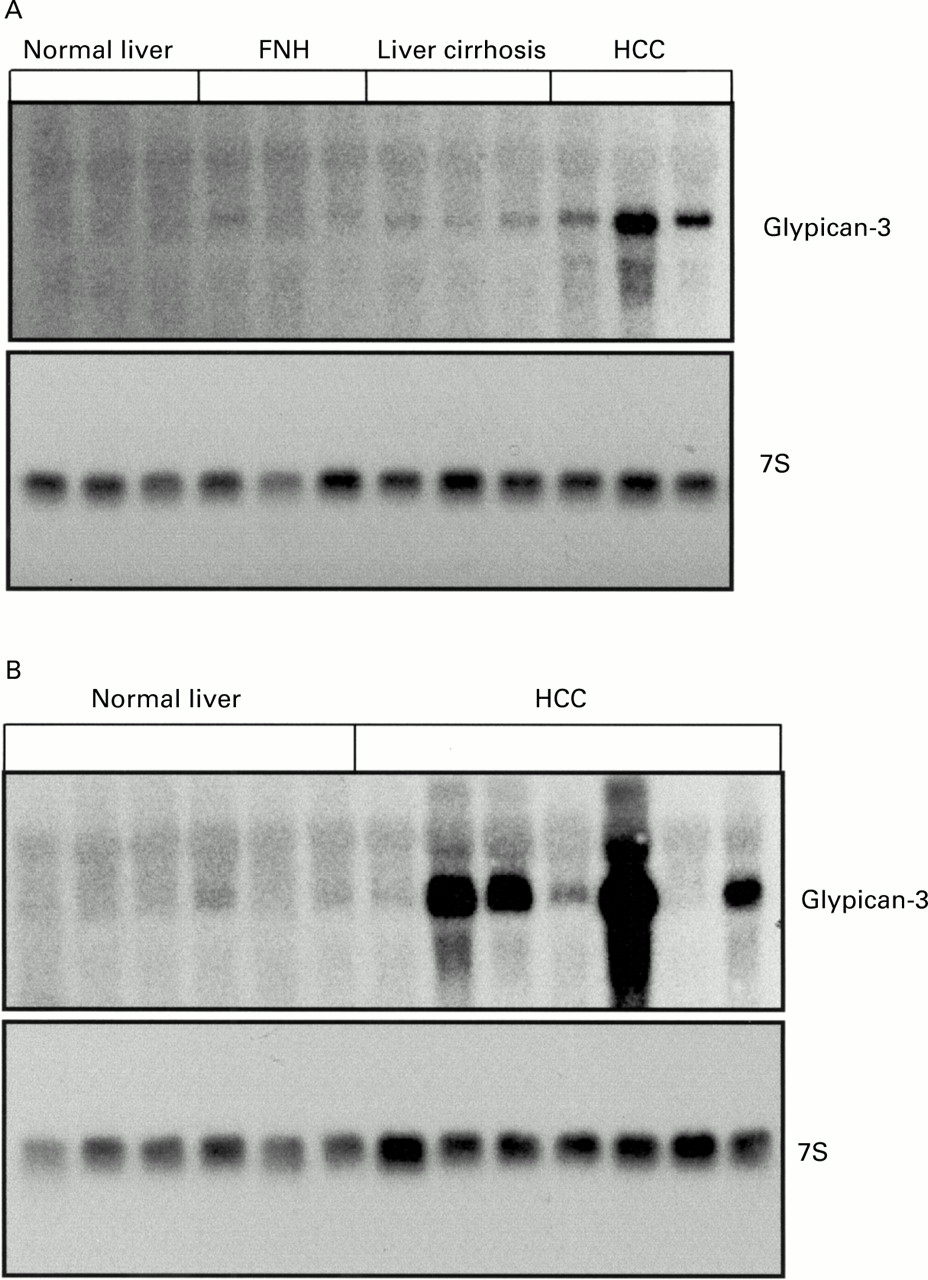
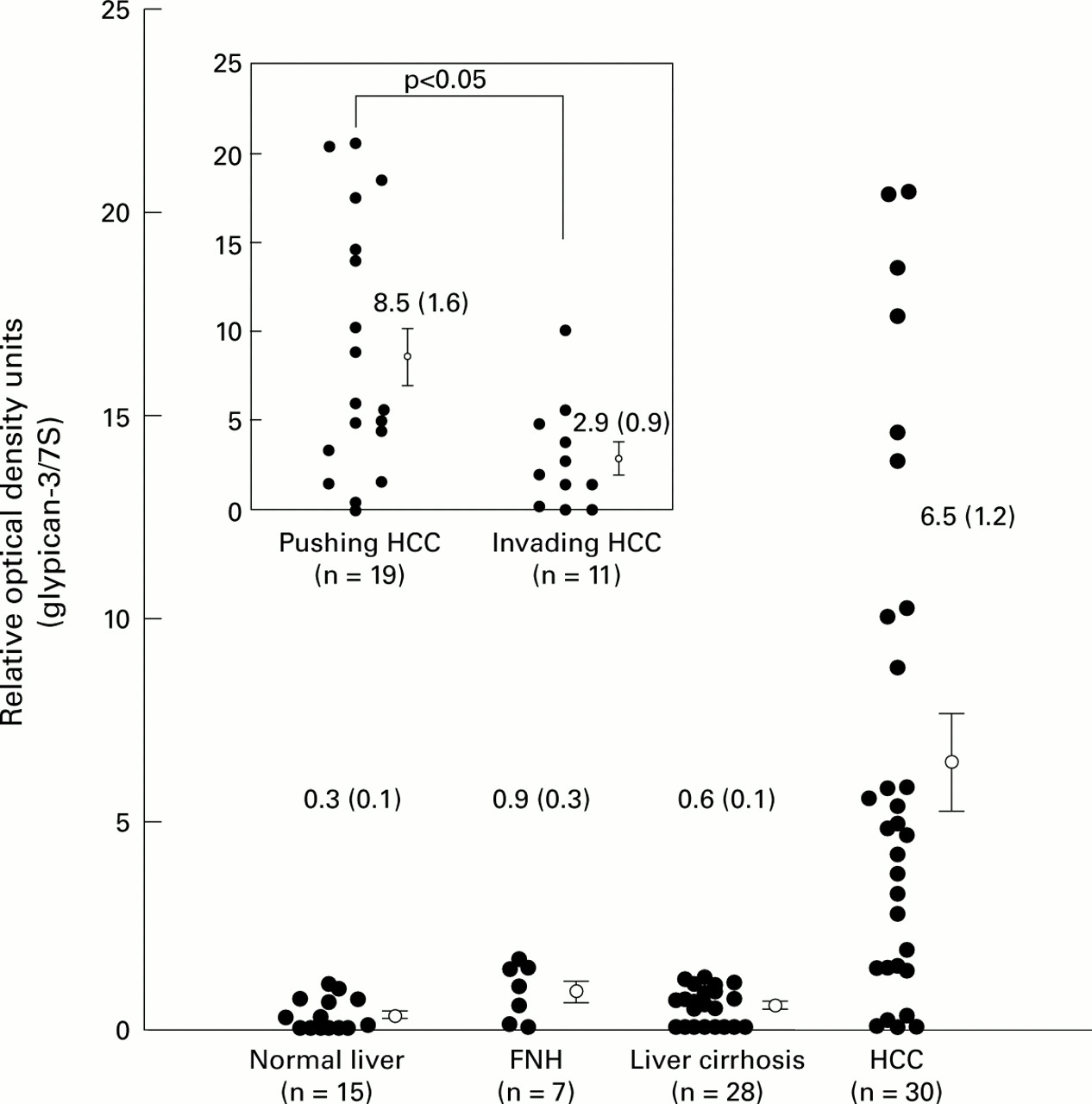
We first screened three normal liver samples, three cirrhotic liver specimens, and three HCC samples for expression of various glypicans. It has been shown previously that expression of glypican-2 and glypican-5 is restricted to the adult and fetal brain.21 Therefore, we examined expression of the other four known members of the glypican family in the above mentioned tissues. This analysis revealed that glypican-3, but not the other members of the glypican family, was highly expressed in HCC but not in liver cirrhosis or in the normal liver (fig 1). Therefore, we examined expression of glypican-3 more thoroughly in various liver tissues. Northern blot analysis was performed on total RNA isolated from 15 normal liver tissues, seven FNH tissue samples, 28 liver cirrhosis samples, and 30 HCC samples. The 2.3 kb glypican-3 mRNA transcript was below the level of detection in six of 15 (40%) normal liver tissue samples and was weakly present in nine of 15 (60%) normal liver tissue samples. Glypican-3 mRNA was moderately present in three and weakly present in four FNH tissue samples and weakly in 21 of 28 (75%) liver cirrhosis tissue samples. In the remaining seven liver cirrhosis samples, glypican-3 mRNA levels were below the level of detection (fig2A). In contrast, in HCC tissues, glypican-3 mRNA was expressed at different levels. Thus glypican-3 mRNA was present at high levels in 20 of 30 (67%), at moderate levels in five of 30 (17%), at low levels in three of 30, and was below the level of detection in two of 30 HCC samples (fig 2A, 2B, fig 3). Densitometric analysis of all northern blots indicated that compared with normal liver tissues, FNH, liver cirrhosis, and HCC exhibited threefold (NS), twofold (NS), and 21.7-fold increases (p<0.01) in glypican-3 mRNA expression levels. When FNH and liver cirrhosis samples were compared with HCC samples, HCC samples had 7.2-fold (p<0.05) and 10.8-fold (p<0.01) higher glypican-3 mRNA expression values than FNH and liver cirrhosis samples, respectively (fig 3). Glypican-3 mRNA values were above the mean value of the non-malignant groups in 25 of 30 (75%) HCC samples.

Northern blot analysis of glypican-1, -3, -4, and -6 in normal liver, liver cirrhosis, hepatocellular carcinoma (HCC), and focal nodular hyperplasia (FNH). Total RNA (20 μg) was size fractionated, blotted, and hybridised with the indicated32P labelled probes.

Northern blot analysis. (A) Glypican-3 mRNA in normal liver, focal nodular hyperplasia (FNH), liver cirrhosis, and hepatocellular carcinoma (HCC). (B) Northern blot analysis of glypican-3 mRNA in normal liver and HCC. Total RNA (20 μg) was size fractionated, blotted, and hybridised with 1×106 cpm/ml32P CTP labelled glypican-3 cRNA probe and 1×105 cpm/ml 32P dCTP 7S cDNA probe.

Densitometric analysis. The ratio of the optical density of glypican-3 mRNA to the corresponding 7S signals was calculated and expressed as mean (SEM). FNH, focal nodular hyperplasia; HCC, hepatocellular carcinoma. p<0.01 (HCC v normal); p<0.05 (HCC v FNH); p<0.01 (HCC v liver cirrhosis). Insert: comparison of expression of glypican-3 mRNA between pushing and invading tumours.
LOCALISATION OF GLYPICAN-3 mRNA EXPRESSION BY IN SITU HYBRIDISATION
In situ hybridisation was performed in normal liver, FNH, liver cirrhosis, and HCC tissues to determine the exact cellular site of glypican-3 mRNA synthesis. Although glypican-3 mRNA was not detected in normal liver samples by northern blot analysis, it was weakly present in the cytoplasm of all bile ductal cells and all hepatocytes (fig 4A,4B). In FNH (fig 4C, 4D) and liver cirrhosis (fig 4E, 4F) tissue sections, glypican-3 mRNA signals were also weakly to occasionally moderately present in the cytoplasm of hepatocytes forming regenerated hepatic nodes. Furthermore, bile ductal cells in liver cirrhosis (fig4E, 4F) and in bile ductal cells adjacent to the FNH (fig 4D, insert) exhibited weak to moderate glypican-3 mRNA staining. In contrast, in HCC, moderate to intense glypican-3 mRNA signals were present in the cytoplasm of most cancer cells (fig 4G, 4H). HCC cells of pushing tumours exhibited moderate to intense glypican-3 in situ hybridisation signals (fig 4H, insert) whereas cancer cells of invading tumours exhibited lower glypican-3 expression levels (fig 4G, upper left insert). However, in most cases the glypican-3 mRNA signals were still more intense in comparison with those observed in benign liver diseases. In situ hybridisation of consecutive tissue sections with the glypican-3 sense probe did not produce any specific signal (fig 4G, insert).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
In situ hybridisation of glypican-3 in normal liver tissue (A, B), focal nodular hyperplasia (FNH) (C, D), liver cirrhosis (E, F), and hepatocellular carcinoma (HCC) (G, H). In normal liver tissues, weak glypican-3 mRNA signals were present in bile ductal cells and in hepatocytes (magnification: A ×50, B ×100). In FNH (magnification: C ×50, D ×100; D insert, bile duct adjacent to FNH tissue ×100) and liver cirrhosis (magnification: E ×50, F ×100), glypican-3 mRNA signals were weakly to occasionally moderately present in hepatocytes and weakly to moderately present in bile ductal cells in liver cirrhosis and in bile ductal cells adjacent to FNH tissue. In HCC, glypican-3 mRNA was more intensely present in tumours with a pushing growth behaviour (magnification: H ×50, H insert ×200) than in those with an invading growth behaviour (magnification: G ×50, G upper left insert ×200). In situ hybridisation in HCC with the glypican-3 sense probe showed no signal (magnification: G insert ×100).
RELATIONSHIP OF GLYPICAN-3 mRNA EXPRESSION TO HISTOPATHOLOGICAL AND CLINICAL DATA
Due to the variation in glypican-3 mRNA expression in HCC samples, we analysed whether histopathological and clinical parameters showed a relationship with glypican-3 mRNA expression. According to the densitometric data, there were no differences in expression between stage III and stage IV tumours (p>0.05) and between well/moderately differentiated tumours (grades 1 and 2) and poor/undifferentiated tumours (grades 3 and 4) (p>0.05) (table 1). Comparison of HCC with different growth characteristics revealed that pushing tumours exhibited 2.9-fold higher glypican-3 mRNA levels than invading tumours (p<0.05) (fig 2 insert). Furthermore, there was no significant difference between glypican-3 expression and age or sex of the patient (table 1).
Discussion
It has been shown previously that glypican-3 is expressed at high levels in HCC and that it may serve as a tumour marker, especially for small tumours.17 However, the above mentioned study did not address the potential importance of other nodular lesions in the liver, in the context of high glypican-3 expression observed in HCC, nor did it investigate glypican-3 mRNA expression with respect to histological subtype of HCC. In this study, glypican-3 mRNA expression was examined in the normal liver, FNH, liver cirrhosis, and HCC. Glypican-3 mRNA was weakly expressed in some normal liver samples, in all FNH samples, and in some liver cirrhosis samples by northern blot analysis. In contrast, glypican-3 mRNA levels were increased in 75% of HCC samples compared with the mean value of the non-malignant groups. These findings indicate that glypican-3, but not other glypicans, may be a potential marker for HCC, and suggest that glypican-3 may be useful in the differential diagnosis of benign and malignant nodular hepatic lesions.
Interestingly, in HCCs, glypican-3 mRNA levels were significantly higher in pushing than ininvading tumours. According to a previous study, the pushing tumour type grows expansively and compresses the surrounding normal tissue, creating a rim of compressed atrophic liver parenchyma that forms a pseudocapsule between the malignant and surrounding benign tissue. Most pushing tumours are resectable despite their size. In contrast, the invading tumour type grows without anatomical clear margins, and there is no pseudocapsule separating the tumour from the surrounding normal liver which limits curative resectability in many cancers.18 Therefore, classification of a pushing or invading growth pattern in HCC reflects the local growth behaviour of this malignancy—that is, pushing tumours have a less aggressive growth behaviour than invading tumours. It is suggested that while glypican-3 may serve as a tumour marker for HCC, this seems especially true for the expansively growing pushing tumour type.
Statistical analysis of the densitometric northern blot data did not show any relationship between glypican-3 mRNA levels and tumour stage or tumour grading. However, most of our patients belonged to tumour stages III and IV (93.3%), and therefore more samples, including earlier tumour stages, are required to determine if the alteration of glypican-3 expression is related to tumour stage. However, the two stage II disease samples exhibited enhanced glypican-3 mRNA expression, indicating that in early tumour stages high levels of glypican-3 are also present. This is consistent with our observation that glypican-3 expression seems to be related to the histological subtype of HCC rather than tumour stage.
In the present study, glypican-3 mRNA expression was slightly higher in FNH and liver cirrhosis compared with normal controls, although no significant differences were found. In situ hybridisation revealed the presence of glypican-3 mRNA in the hyperplastic hepatocytes in FNH and liver cirrhosis. Furthermore, moderate glypican-3 mRNA signals were also present in bile ductal cells in FNH and liver cirrhosis tissues. In these two benign chronic liver disorders, the hyperplastic nodules, which are the main components of this disease,22 ,23 are composed of normal hepatocytes but not bile ductular cells. Therefore, only weak glypican-3 mRNA signals were detected in both disorders by northern blot analysis.
The exact biological functions of glypican-3 are still not well understood. However, there is increasing evidence that glypican-3 is mainly involved in growth inhibitory functions, and that loss of glypican-3 expression may contribute to uncontrolled cell growth. Alteration of glypican-3 expression due to gene mutations leads, for example, to an overgrowth syndrome characterised by multiple embryonic abnormalities.6 ,7 Additional studies in mesotheliomas, breast cancers, and ovarian cancers indicate that glypican-3 has growth inhibitory functions, influences apoptosis, and may function as a tumour suppressor gene in these disorders.8-10 In our study, HCCs expressed high levels of glypican-3 mRNA, suggesting that glypican-3 may act as a tumour promoter in this malignancy. In contrast, the pushing tumours that display a less aggressive local growth behaviour exhibited increased glypican-3 levels in comparison with invasive tumours that grow aggressively into the adjacent normal liver tissue. These observations suggest that while glypican-3 may promote local cancer cell growth, it may also inhibit tissue invasion and metastasis. Alternatively, enhanced glypican-3 levels observed in HCCs might just be an epiphenomenon resulting from induction of a yet unknown factor or through other mechanisms. Irrespective of this, further studies are required to answer these questions.
To date, there are no commercial antibodies available for glypican-3. Therefore, it is not known if glypican-3 protein levels parallel changes in glypican-3 mRNA levels in liver diseases, and if glypican-3 protein is detectable and can be measured in the serum of patients with HCC. Further studies may clarify these issues. Our study provides a potentially promising tool to better differentiate HCC from benign liver disorders.
In conclusion, glypican-3 mRNA is markedly overexpressed in HCC compared with normal liver, focal nodular hyperplasia, or liver cirrhosis. In addition, in HCC, pushing tumours express significantly more glypican-3 mRNA than the invasive subtype. These findings suggest that glypican-3 may play a role in the pathobiology of HCC by influencing local growth behaviour, and that it may serve as a new marker for differentiation of benign and malignant liver disease.
Abbreviations used in this paper
- HCC
- hepatocellular carcinoma
- FNH
- focal nodular hyperplasia
- HSPGs
- heparansulphate proteoglycans
- HGF
- hepatocyte growth factor
- EGF
- epidermal growth factor
- HB-EGF
- heparin binding EGF-like growth factor
- SDS
- sodium dodecyl sulphate