


Abstract
In an attempt to provide a better understanding of the scope and limitations of animal models used in some drug development programs and to further our understanding of potential metabolic bioactivation reactions, we have undertaken studies to profile the monoamine oxidase A and B (MAO-A and -B, respectively) activities in liver and brain mitochondrial preparations obtained from a variety of species using a series of 1-methyl-4-aryl-1,2,3,6-tetrahydropyridinyl substrates. Mitochondrial preparations were incubated with substrates at 37°C in the presence or absence of clorgyline, (R)-deprenyl, or a mixture of these two propargylamines to inhibit MAO-A, MAO-B, or both enzymes. The rates of formation of the corresponding dihydropyridinium metabolites were estimated spectrophotometrically. MAO-B was found to be the principal enzyme present in all tissues. Human liver displayed more MAO-A activity than the liver of any other species studied; subhuman primates displayed little or no detectable MAO-A activity. The properties of the preparations from rat liver were most similar to those from human liver with respect to the MAO-A/MAO-B ratios and the kinetic parameters of the four substrates used to profile enzymatic activity. The kinetic properties of mitochondrial preparations from bovine liver, a commonly used source of purified MAO-B preparations, were consistently different from all of the other species studied. The mitochondrial preparations from rabbit brain and liver also were unusual in that they displayed relatively low MAO activities. Additionally, these enzyme activities were considerably less susceptible to inhibition by clorgyline and (R)-deprenyl. Finally, an exceptionally low MAO-B liver/brainVmax/Km ratio was observed with the mitochondria obtained from the C57BL/6 mouse, an effect that may contribute to the susceptibility of this strain to the toxic effects of the parkinsonian-inducing neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine.
The flavoproteins monoamine oxidase A and B (MAO-A and -B) are the principal enzymes responsible for the metabolism of the neurotransmitters dopamine, norepinephrine, epinephrine, and serotonin (Waldmeier, 1987; Kopin, 1994). These enzymes are found throughout the body and are localized in the outer mitochondrial membrane. For reasons that remain unclear, often only one form of the enzyme is present in a specific organ and/or within a specific cell type (Trendelenburg et al., 1987; Yu et al., 1992). Extensive studies have been performed over the years to characterize the properties of MAO-A and -B (Kalgutkar and Castagnoli, 1995; Silverman, 1995; Castagnoli et al., 1997; Wouters, 1998).
In addition to their roles in the regulation of neurotransmitter levels, these enzymes also catalyze the oxidation of xenobiotic amines (Strolin Benedetti and Tipton, 1998) including dietary tyramine (Hauptmann et al., 1996). Furthermore, it is well known that the parkinsonian inducing neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine [MPTP (1)] is an excellent MAO-B substrate (Trevor et al., 1987). The neurotoxicity of MPTP is dependent on its MAO-B-catalyzed oxidation to the dihydropyridinium intermediate 1-methyl-4-phenyl-2,3-dihydropyridinium species (MPDP+; 2), which undergoes further oxidation to the 1-methyl-4-phenylpyridinium species (MPP+; 3), the ultimate toxin.
In view of the diverse nature of MAO substrates and inhibitors, a better appreciation of the factors that may influence the interactions of MAO-A and MAO-B with xenobiotics, as has been detailed for the cytochrome P-450 family of oxidases (Guengerich, 1997), could prove useful in the design and development of new therapeutic agents. As an initial effort toward this goal, we have undertaken a series of systematic studies aimed at characterizing species- and organ-dependent differences in MAO activity. Some evidence for such differences has been reported. Garrick and Murphy (1980) have observed species differences in the inhibition curves of clorgyline, (R)-deprenyl, and pargyline, all potent mechanism-based inactivators of MAO-A and/or MAO-B, on the deamination of dopamine, serotonin, tyramine, and 2-phenylethylamine. Dramatic species differences in the inhibition of MAO-B by various oxadiazolones and oxadiazolethiones also have been described (Krueger et al., 1995). Furthermore, the substrate and inhibition properties of a series of β-aminoamides were shown to be tissue dependent (O’Brien et al., 1995).
In the present study, the rates of α-carbon oxidation of a panel of tetrahydropyridinyl substrates have been compared. These substrates were selected in part because of our interests in MAO-catalyzed metabolic bioactivation processes (Castagnoli et al., 1997). All of the corresponding dihydropyridinium metabolites absorb light at wavelengths considerably longer than those of the substrate molecules, making it possible to estimate the rates of oxidation with the spectrophotometric assay described below.
Materials and Methods
Chemicals.
The following tetrahydropyridinyl derivatives (see Fig. 1 for structures), used as substrates in these studies, were synthesized in our laboratory: 1,1-methyl-4-(3-methoxyphenyl)-1,2,3,6-tetrahydropyridine (4; Youngster et al., 1987), 1-methyl-4-(1-methyl-2-pyrrolyl)-1,2,3,6-tetrahydropyridine (5; Nimkar et al., 1996), 1-methyl-4-(3-ethyl-2-furanyl)-1,2,3,6-tetrahydropyridine (6; Yu and Castagnoli, 1999), and, as described below, 1-methyl-4-(2-isopropylphenyl)-1,2,3,6-tetrahydropyridine (7). Spectroscopic data and elemental analyses for the oxalate salt of 4 has not been reported and is as follows: 4 · C2H2O4 is a white solid (MeOH/Et2O): m.p.: 147°C; 1H NMR [dimethyl sulfoxide (DMSO)-d6] δ 7.28 (1 H, t,J = 8.0 Hz), 7.08 (1 H, d, J = 7.7 Hz), 6.99 (1 H, t, J = 2.2 Hz), 6.88 (1 H, dd,J = 2.4 Hz, J = 8.0 Hz), 6.04 (1 H, M), 3.79 (2 H, M), 3.77 (3 H, s), 3.34 (2 H, M), 2.81 (3 H, s), 2.73 (2 H, bs); 13C NMR (DMSO-d6) δ 165.1, 159.5, 140.0, 133.6, 129.6, 117.2, 117.1, 113.4, 110.5, 55.1, 51.2, 49.5, 41.6, 23.8; gas chromatography (GC)-EIMS, m/z (rel intensities) 203 (100%), 202 (63), 159 (20), 145 (24), 115 (20), 96 (59), 82 (15), 51 (11); UV (nm, MeOH) 213, 246, 288. Analysis calculated (C15H19NO5): C, 61.42; H, 6.53; N, 4.78. Found: C, 61.28; H, 6.49; N, 4.73. Compound 7 was prepared by the condensation of the lithium salt derived from 2-isopropylbromobenzene with 1-methyl-4-piperidone followed by acid catalyzed dehydration of the resulting carbinol. The product was purified as its oxalate salt: 7 · C2H2O4(MeOH/Et2O): m.p.: 204–205°C; 1H NMR (DMSO-d6) δ 7.30 (1 H, dt,J = 7.8 Hz, J = 1.5 Hz), 7.24 (1 H, dd, J = 7.8 Hz, J = 1.5 Hz), 7.13 (1 H, dt, J = 7.6 Hz,J = 1.5 Hz), 7.02 (1 H, ddd, J= 7.6 Hz, J = 1.5 Hz, J = 0.4 Hz), 5.49 (1 H, M), 3.70 (2 H, M), 3.29 (2 H, t, J= 6.0 Hz), 3.09 (1 H, sept, J = 6.9 Hz), 2.80 (3 H, s), 2.53 (2 H, M), 1.14 (6 H, d, J = 6.9 Hz);13C NMR (DMSO-d6) δ 163.8, 145.5, 136.3, 127.6, 127.3, 125.1, 118.7, 51.1, 49.6, 41.8, 28.6, 28.2, 24.0; GC-EIMS, m/z (rel intensities) 215 (95), 214 (85), 200 (86), 172 (45), 157 (100), 142 (52), 128 (48), 115 (42), 96 (53); UV (nm, MeOH) 212, 242, 265. Analysis calculated (C17H23NO4): C, 66.86; H, 7.59; N, 4.59. Found: C, 67.02; H, 7.42; N, 4.43.
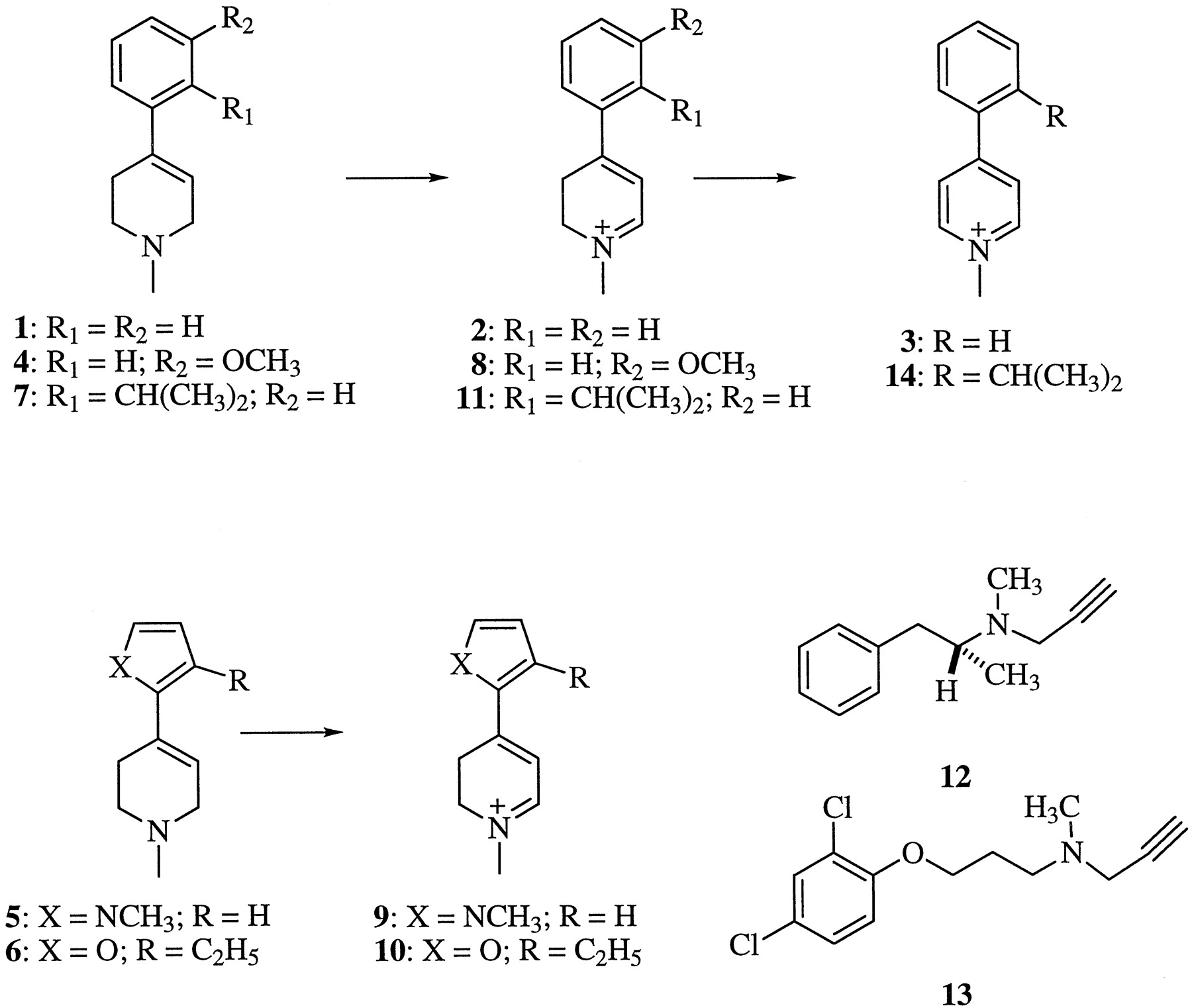
Chemical structures of study compounds.
The structures (and reported syntheses) of the corresponding dihydropyridinium metabolites are 2 (Chiba et al., 1985; for MPTP) 8 (for 4), 9 (Nimkar et al., 1996; for 5), 10 (for 6), and 11 (for 7). Clorgyline and (R)-deprenyl were obtained from Research Biochemicals Inc. (Natick, MA). Deferoxamine mesylate was obtained from Sigma Chemical Co. (St. Louis, MO). All other chemicals were reagent, bioanalytical, or HPLC grade (Fisher Scientific Products, Fair Lawn, NJ).
Preparation of Mitochondria and Semipurified MAO-A and -B.
Protocols for obtaining tissues were approved by the appropriate committee at the institution of origin. In addition, these were reviewed and approved by the Virginia Tech Animal Care Committee, as were the protocols for all animal procedures performed at Virginia Tech. Receipt of the human tissue was approved by the Virginia Tech Committee on Human Experimentation. Experiments were carried out with mitochondria prepared from brains and livers of male C57BL/6 and ICR mice, Sprague-Dawley rats, New Zealand White rabbits, Beagle dogs, cynomolgus monkeys, and baboons (Papio papio ursinus). For human and bovine preparations, only liver samples were available. The preparation of the mitochondrial fractions followed literature procedures (Salach and Weyler, 1987) except that minor volume changes were made with the brain preparations. Tissue homogenates from individual animals were prepared as follows (dog,n = 2; baboon, n = 3; monkey,n = 2). Pooled tissue homogenates were prepared using the following numbers of animals per pooled preparation (number of pooled preparations): rabbit, n = 2 (2); rat,n = 2 to 10 (3); ICR mice, n = 5 (2); and C57BL/6 mice, n = 3 to 12 (3). Individual human liver homogenates were prepared from tissues obtained from five donors. Data derived from individual animals or humans are indicated in the tables. These preparations were stored in aliquots of 100 to 200 μl at −70°C. The aliquoted samples were thawed as needed and mixed with glycerol-containing buffer [50 mM sodium phosphate buffer, pH 7.4, containing 50% (w/v) glycerol] for the initial protein concentration. All protein concentrations were determined according to the Coomassie Brilliant Blue dye binding method of Bradford (1976) with BSA used as standard. For the assays, these were further diluted using 50 mM sodium phosphate buffer, pH 7.4, to a concentration appropriate for the experiment. Solutions of all inhibitors and substrates also were prepared in the same phosphate buffer. Each assay was run in duplicate, and the results differed by less than 5%.
Semipurified human placental MAO-A and bovine liver MAO-B were obtained according to the method of Salach (1987) with minor modifications as previously described (Kalgutkar et al., 1994). Final enzyme preparations (6–8 μM) were stored at −15°C.
Optimization of Preincubation Conditions to Inhibit MAO-A or -B Activity by Selective Inhibitors.
These studies used the 1-methyl-2-pyrrolyl analog 5 because it is a good substrate for both forms of the enzyme. The stability of the dihydropyridinium metabolite 9, its relatively large molar extinction coefficient (ε = 25,000 M−1), and its maximal absorbance at 420 nm, which is far removed from mitochondrial background, made 5 an excellent substrate for these experiments (Flaherty et al., 1996).
The optimal preincubation time to inhibit the MAO-A activity in the mitochondrial preparations was determined as follows. Mixtures (125 μl) consisting of previously diluted rat liver mitochondria (0.6 mg protein/ml) and 3.75 × 10−8 M (R)-deprenyl were incubated in phosphate buffer at 37°C for 15 min to inhibit MAO-B activity in the preparation. At this time, 125 μl of a stock solution of clorgyline (6 × 10−8 M) was added, the incubations were continued at 37°C, and 250 μl of the methylpyrrolyl analog 5 (2 mM) was added at varying times (0–25 min). After an additional 15 min, the reactions were quenched by the addition of 20 μl of 70% aqueous HClO4, the denatured protein was removed by centrifugation at 16,000g for 5 min, and the absorbance of the dihydropyridinium metabolite was measured on a Beckman DU7400 diode array spectrophotometer. To estimate the time course for the inhibition of MAO-B by (R)-deprenyl, the corresponding experiment was performed in which rat liver mitochondria were treated first with clorgyline (3 × 10−8M) followed by the addition of (R)-deprenyl (3 × 10−7 M) and then substrate (2 mM). Control values (100% activity) were obtained by measuring the extent of oxidation of 5 by mitochondria that had been treated only with the first inhibitor.
The optimal concentrations of the selective inhibitors were determined as follows. Mixtures (262.5 μl) consisting of diluted rat liver mitochondria (0.29 mg protein/ml) and varying concentrations (3 × 10−4 to 3 × 10−10M) of clorgyline or (R)-deprenyl were incubated at 37°C for 15 min. At this time, 237.5 μl of the methylpyrrolyl analog 5 (2 mM) was added, and the absorbance of the dihydropyridinium metabolite formed was measured after 15 min. Control samples (100% activity) were made by incubating the mitochondrial preparation with no inhibitor.
Studies on 2-Isopropylphenyl Analog 7.
Compound 7 was examined as a potential MAO-A-selective substrate. Evidence of the instability of the resulting dihydropyridinium metabolite 11, however, led us to attempt to identify the corresponding pyridinium oxidation product 14. A 1-h postincubation mixture of 7 (100 μM) and MAO-A (0.13 μM) in glycerol-containing buffer was treated with NaBD4 in 500 μl of methanol. The mixture was left at room temperature for an additional hour and then extracted with 1.5 ml of ethyl acetate. The organic layer was dried over anhydrous sodium sulfate, filtered, and evaporated under a gentle stream of nitrogen. The residue was reconstituted with 100 μl of methanol, and 1 μl was analyzed by GC (model HP 5890)–mass spectrometry (model HP 5970; GC-MS) using an HP-1 capillary column (12.5 M × 0.2 mm × 0.33 μM). The injection port temperature was 225°C. The GC oven temperature was 60°C for 1 min and then was increased at 25°C/min to 275°C.
Determination of MAO-A/MAO-B (12) in Mitochondrial Preparations.
These studies were performed using the 1-methyl-2-pyrrolyl analog 5 as substrate (see above) at 2 mM, a concentration that is well above the Kmvalue for this compound in all the species examined (see Table 3). The activities of total MAO, MAO-A, or MAO-B were measured after 15-min preincubations of mitochondria with no inhibitor or with 3 × 10−7 M (R)-deprenyl (12) or 3 × 10−8 M clorgyline (13), respectively. The activity remaining when both inhibitors were present was defined as the residual activity. When significant, the residual activity was subtracted from the estimated MAO-A and -B activities. The percentages of MAO-A, MAO-B, and residual activities were calculated with the sum of these activities as 100%. In related experiments, mitochondrial preparations were preincubated with the iron chelator deferoxamine (2 × 10−4 M, final concentration) in the presence or absence of both inhibitors to examine the influence of adventitious iron on the oxidation of the tetrahydropyridinyl substrates (Di Monte et al., 1995).
Vmax/Km values for the substrates 1, 4, 5, and 6 examined in the study
Determination of Mitochondrial MAO Activity of Selected Tetrahydropyridinyl Substrates.
Mitochondrial preparations (500 μl final volume containing 0.15 mg protein/ml for liver samples and 0.3 mg protein/ml for brain samples) were incubated with various concentrations of the tetrahydropyridinyl substrates. In some instances, higher protein concentrations were used to give rates of oxidation that were sufficiently fast to measure with confidence. The incubations were run for 15 to 60 min, depending on the rate of product formation, with gentle agitation in a 37°C water bath. When assaying for ratios of MAO-A/MAO-B activity with the mixed substrates, the 1-methyl-2-pyrrolyl (5) and the 3-ethyl-2-furanyl (6) analogs, the mitochondrial preparations were preincubated in 237.5 μl of the buffer containing either no inhibitor, 3 × 10−8 M clorgyline, 3 × 10−7 M (R)-deprenyl, or both inhibitors for 15 min, followed by the addition of 237.5 μl of the substrate solution. In all experiments, the reactions were quenched by the addition of 20 μl of 70% perchloric acid, and the denatured protein was removed by centrifugation at 16,000g for 5 min. The incubation times for each substrate were 45 min for the methoxyphenyl analog 4 and MPTP, 5 to 15 min for the pyrrolyl analog, and 6 to 60 min for the furanyl analog. The λmax values and the molar extinction coefficients (ε) of the dihydropyridinium metabolites generated in these MAO-catalyzed reactions were 343 nm and 16,000 M−1 for 2 (Kalgutkar et al., 1994) and 8, 420 nm and 25,000 M−1 for 9 (Bai, 1991), and 400 nm and 23,000 M−1 for 10 (Yu and Castagnoli, 1998). A synthetic standard of 8 was not available, so the ε value of 2 was used. Values for Vmax andKm were obtained from Lineweaver-Burk double reciprocal plots of velocities versus substrate concentrations. Duplicate analyses gaveVmax/Km values that differed by 10% or less.
Results
Stability of MAO Activity in Mitochondria.
MAO-B activities in dog brain and liver were assayed at 0, 1, 2, 4, 6, 9, and 12 months in triplicate. Under the conditions of the present study, MAO-B activities in dog brain and liver were stable over a period of 12 months when stored at −70°C. The ranges forVmax/Km(mean ± S.E.) given as nmol MPDP+ formed/min/mg protein of the activities observed in these studies were 21.0 to 25.7 (23.6 ± 0.69) for brain and 62.2 to 83.0 (69.0 ± 2.77) for liver. Our general observations suggest that the other mitochondrial preparations also were stable under these storage conditions.
Time- and Concentration-Dependent Inhibition of MAO by Clorgyline and (R)-Deprenyl.
Estimations of the MAO-A/MAO-B activity ratios required that kinetic measurements be made when one form of the enzyme was selectively and quantitatively inhibited in the presence of the other form. These studies used the pyrrolyl analog 5 (2 mM), a mixed MAO-A/MAO-B substrate.
MAO-A activity in rat liver mitochondria that had been preincubated with (R)-deprenyl was fully inhibited by clorgyline by 15 min (Fig. 2A). Similarly, MAO-B activity was completely inhibited by (R)-deprenyl by 10 min (Fig.2B). Therefore, a 15-min preincubation period was selected for both inhibitors. Note that the inactivation reactions were too rapid to capture a true 0-min time point, and thus initial plotted activity does not equal 100% in this or the following experiment.

Time course for the inhibition of the MAO-A-catalyzed oxidation of the pyrrolyl substrate 5 (2 mM) by 3 × 10−8 M clorgyline in rat liver mitochondrial preparations (0.15 mg protein/ml) preincubated for 15 min with 3 × 10−7 M (R)-deprenyl (A) and that of the corresponding MAO-B-catalyzed reaction by 3 × 10−7 M (R)-deprenyl in mitochondria preincubated with 3 × 10−8 M clorgyline (B). In both plots, the 5% residual activity observed when both inhibitors were present was subtracted.
Figure 3A shows the influence of various concentrations of the selective MAO-A inhibitor clorgyline on the rate of oxidation of the pyrrolyl substrate 5 by rat liver mitochondrial preparations. The corresponding plot for the inhibition of MAO-B activity is shown in Fig. 3B. The selective inhibition of MAO-A in these preparations was indicated by the leveling of the curve in the concentration range of 10−7 to 10−8 M inhibitor. As reported elsewhere (Murphy et al., 1979), MAO-B activity is inhibited by clorgyline only at much higher concentrations (>10−6 M). The biphasic curve shown in Fig. 3B indicates that (R)-deprenyl selectively inhibits MAO-B within the concentration range of 10−6 to 10−7 M. MAO-A is inhibited only at concentrations of >10−6 M. Consequently, 3 × 10−8 M clorgyline and 3 × 10−7 M (R)-deprenyl were used to inhibit MAO-A and MAO-B activities in mitochondrial preparations, respectively. The higher concentration of (R)-deprenyl relative to clorgyline was dictated by the higher concentrations of MAO-B found in all mitochondrial preparations.

Concentration-dependent inhibition by clorgyline (A) and by (R)-deprenyl (B) of the MAO-catalyzed oxidation of the pyrrolyl substrate 5 (1 mM) in rat liver mitochondrial preparations (0.15 mg protein/ml). Mitochondria were preincubated with the inhibitors for 15 min at 37°C before assay for remaining activity.
Linearity of Production of Metabolites with Time.
The linearity of the rates of dihydropyridinium metabolite formation in these mitochondrial preparations was examined to determine whether the kinetic measurements were being made under steady-state conditions. Figure 4 illustrates the time-dependent increase in the concentration of MPDP+ (2) from MPTP (1) by human liver mitochondria. The increase in absorption at λmax 343 nm with time was reasonably linear (r2 = 0.995) for at least 50 min. Because no changes in the shape of absorption curves were observed during this period of time, we concluded that MPDP+ is stable under these conditions. Similar results were observed with the other substrates (4–6) used in this study. This was not the case, however, with the 2-isopropylphenyl analog 7, a reported MAO-A-selective substrate (Singer and Ramsay, 1991), as discussed below.

Time-dependent increase in the dihydropyridinium metabolite MPDP+ (2) formed from 200 μM MPTP (1) by human liver mitochondria (0.15 mg protein/ml). After subtraction of the background absorption of the mixture, the spectrum were recorded every 5 min for 50 min at 37°C.
Attempts to Develop an MAO-A-Selective Substrate.
MPTP and its 3-methoxy analog 4 are both highly selective MAO-B substrates. In the C57BL/6 mouse brain preparation, which contains about 15% MAO-A activity by our estimates, theVmax/Km values for MPTP in the presence of 3.3 × 10−8 M clorgyline (38.0 nmol product formed/min-mg protein) and in the absence of clorgyline (37.8 nmol product formed/min-mg protein) were essentially the same. The corresponding values for the 3-methoxy analog 4 using human liver mitochondria (18% mean MAO-A activity) were 99.7 nmol/min-mg protein in the presence of 3.3 × 10−7 M clorgyline and 97.3 nmol/min-mg protein in the absence of clorgyline. Consequently, it was possible to assay MAO-B activity without inhibiting MAO-A.
According to a literature report (Singer and Ramsay, 1991), the 4-(2-isopropylphenyl) analog 7, a selective MAO-A substrate, should be a useful reagent to measure MAO-A activity in the presence of catalytically active MAO-B. Figure 5shows the results of studies on the metabolism of 7 using the semipurified placental MAO-A preparation. The time-dependent increase in absorption at 320 nm was consistent with formation of the dihydropyridinium metabolite 11. As required for an MAO-A-mediated reaction, preincubation of the enzyme with clorgyline blocked the production of 11. Also consistent with the earlier report (Singer and Ramsay, 1991), no significant formation of 11 was observed when 7 was incubated with solubilized MAO-B. As shown in Fig. 5, however, the 320-nm absorbing chromophore formed initially in the MAO-A-catalyzed oxidation of 7 underwent a hypsochromic shift with time, consistent with its further oxidation to the pyridinium species 14. Evidence to support this suggestion was obtained by treating the mixture, after a 1-h incubation period, with NaBD4. GC-EIMS analysis (Nimkar et al., 1996) of the resulting reaction mixture showed strong (relative to the base peak) ion intensities at m/z217 (50%), which corresponds to 7-2,6-d2, the expected NaBD4 reduction product of 14 (Fig.6), as well as at m/z 216 (45%) and m/z 215 (40%), the expected fragments resulting from the loss of a hydrogen atom and a deuterium atom, respectively, from the parent ion (Mabic and Castagnoli, 1998). The corresponding mass spectral characteristics of standard 7 were similar to those of 7-d2. This instability might be due to the twist between the heterocyclic ring and the isopropylphenyl ring due to the steric interactions of the isopropyl group with the protons at C3 and C5. These steric interactions would decrease the overlap of the phenyl π-electrons with the dihydropyridinium moiety leading to a decrease in the pKa of 11 and an increase in the equilibrium concentration of the corresponding dihydropyridine free base 15. Compound 15 can undergo autoxidation, a process that is well characterized for related systems (Peterson et al., 1985). Alternatively, the free base may be a substrate for MAO-A. This instability of 11 obviated the possibility of using compound 7 to estimate MAO-A activity.

Time-dependent oxidation of 1-methyl-4-(2-isopropylphenyl)-1,2,3,6-tetrahydropyridine (7) by MAO-A. Semipurified MAO-A (20 nM) was mixed with the substrate (1 mM) in a quartz cell. A UV-VIS spectrum of the mixture was immediately recorded as blank. Then, after subtraction of the blank, the production of the metabolite was measured every 3 min for 30 min at 37°C. Note the shift in λmax with time corresponding to the conversion of the initially formed dihydropyridinium metabolite 11 to the corresponding pyridinium compound 14.
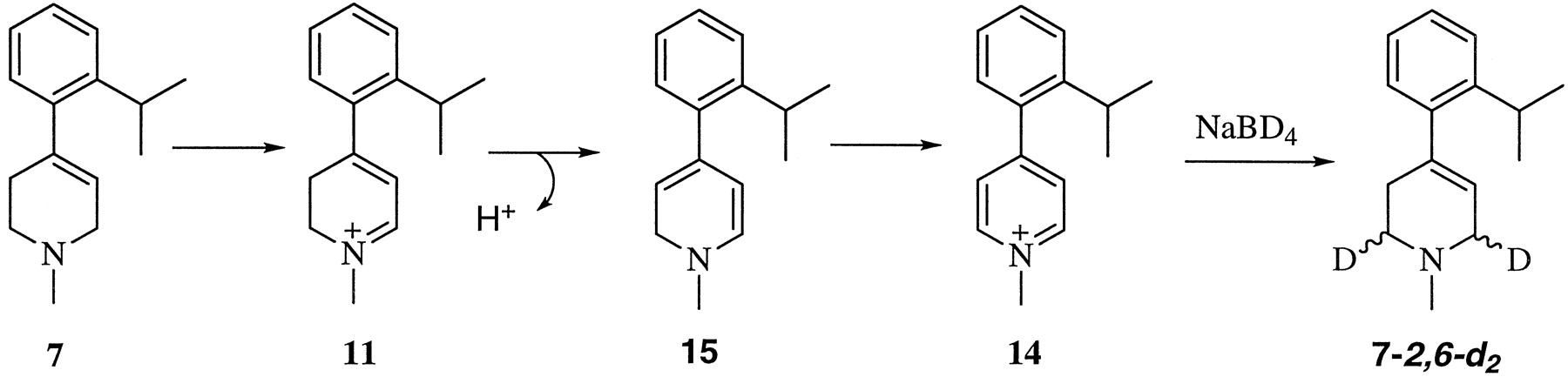
The metabolic fate of 1-methyl-4-(2-isopropylphenyl)-1,2,3,6-tetrahydropyridine (7) and subsequent oxidation of the resulting dihydropyridinium metabolite 11 to form the pyridinium species 14.
Species Differences in MAO-A and -B Catalytic Activity.
Studies to estimate the MAO-A and -B activities in the tissues of interest used the pyrrolyl substrate 5, a mixed A/B substrate, and the selective MAO-B inhibitor (R)-deprenyl (12) to estimate MAO-A activity and the selective MAO-A inhibitor clorgyline (13) to estimate MAO-B activity. Figure 7summarizes the results obtained with dog brain mitochondria and are typical of the quality of the data generated in these studies. The results obtained with all of the tissues examined are summarized in Table 1 numerically and in Fig.8 graphically. These results were obtained with a saturating concentration of the substrate. Estimates ofVmax and Km also were obtained for 5 and the second mixed substrate, the furanyl analog 6 (Table 2). In general, all of the catalytic activity could be accounted for in terms of the A and B forms of the enzyme. The brain and liver mitochondrial preparations isolated from the rabbit were exceptional in that a high percentage of “residual” oxidase activity was present when both MAO-A and -B were inhibited as illustrated in Fig. 9 for the liver preparation.

UV-VIS scans of incubation mixtures containing the 1-methyl-2-pyrrolyl analog 5 (2 mM) in the presence of mitochondrial preparations from dog brain with and without inhibitors. Mitochondrial preparation (0.3 mg protein/ml at final concentration) were preincubated with either no inhibitor, 3 × 10−8 M clorgyline (Clo), 3 × 10−7(R)-deprenyl (Dep), or clorgyline plus (R)-deprenyl for 15 min at 37°C, following which 2 mM concentration of the 1-methyl-2-pyrrolyl analog in phosphate buffer was added. The incubation was continued for an additional 15 min, and the reaction was quenched by the addition of perchloric acid.
MAO-A and MAO-B activities observed with various mitochondrial preparations as estimated by the rates of oxidation of the MAO-A/MAO-B mixed substrate 5 after selective inhibition of each form of the enzyme

Percent brain and liver MAO-A, MAO-B, and residual activity observed in the mitochondrial preparations of various species. Technical details were as described in Table 1.
Kinetic parameters for the human liver mitochondrial MAO-A and MAO-B catalyzed oxidation of 1-methyl-4-(1-methyl-2-pyrrolyl)-1,2,3,6-tetrahydropyridine (5) and 1-methyl-4-(3-ethyl-2-furanyl)-1,2,3,6-tetrahydropyridine (6)

UV-VIS scans of incubation mixtures containing the 1-methyl-2-pyrrolyl analog 5 (2 mM at final concentration) in the presence of mitochondrial preparations from rabbit liver (0.15 mg protein/ml at final concentration) with and without inhibitors. Technical details were as described in Fig. 7.
Species-Dependent MAO Kinetic Profiles Observed with Various Tetrahydropyridinyl Substrates.
A second major thrust of these studies has been to focus on the characterization of the species-dependent differences in liver and brain mitochondrial MAO activity as defined by the kinetic behavior of a panel of tetrahydropyridinyl substrates. In these experiments, we measured total enzyme activity for all four substrates. The results reported in Table3 were obtained by estimates of reaction rates using substrate concentrations that bracketed their apparentKm values. TheVmax and Kmvalues (both of which are listed) were determined from Lineweaver-Burke plots; included is the range of the ratios ofVmax/Km.
Discussion
Species Differences in MAO-A and -B Catalytic Activity.
One of the principal goals of this study was to characterize the MAO-A and -B activities present in the mitochondrial preparations obtained from the species under study. Several measurements of the levels of MAO-A and -B activity using radiographic-based enzyme activities (Saura et al., 1992) have appeared in the literature. Results from a specific inhibitor binding assay indicate that up to 80% of the total MAO activity in human liver is due to MAO-B (Saura et al., 1996). Histochemical studies have shown that MAO activity in the marmoset brain is essentially all due to MAO-B (Willoughby et al., 1988). Similar results have been reported for other subhuman primates byRiachi and Harik (1992), who used a [3H]pargyline binding assay to measure MAO activity. In contrast to the above results, Murphy et al. (1979), who estimated activity by the extent to which clorgyline and (R)-deprenyl inhibited tyramine deamination, reported up to 30% MAO-A activity in various regions of the vervet monkey brain.
In the absence of an MAO-A-selective substrate, we elected to pursue this goal using the 1-methyl-2-pyrrolyl analog 5, a mixed MAO-A/MAO-B substrate, and clorgyline (to inhibit MAO-A and estimate MAO-B activity) or (R)-deprenyl (to inhibit MAO-B and estimate MAO-A activity). The substrate concentration was 2 mM, well above theKm values for all of the enzyme preparations (see Table 3 and discussion below). Consequently, all of these estimates were obtained under Vmaxconditions. Figure 7, which illustrates the type of data generated in these studies, shows the ultraviolet-visible (UV-VIS) scans of incubation mixtures of 5 and dog brain mitochondrial preparations (0.3 mg protein/ml) in the presence and absence of the inhibitors. The spectrum obtained from the sample preincubated with clorgyline indicates the MAO-B activity in the mitochondria and that with (R)-deprenyl indicates the MAO-A activity. When the sample was preincubated with both inhibitors, the remaining activity was less than 3% of the total. From these data, we have estimated dog brain MAO-B activity to be 94%, MAO-A activity to be about 3%, and residual activity to be less than 3% (Table 1).
The results obtained with all of the mitochondrial preparations are summarized in Table 1 and Fig. 8. MAO-B was the principal enzyme found in all liver and brain preparations. Of particular interest, when considering models for human studies, was the observation that the human liver preparation displayed the highest level of MAO-A activity (18%) of all species studied. As observed earlier by May et al. (1991), significant (13%) MAO-A activity also was present in the rat liver. In all of the other species studied, the levels of MAO-A activity were 1% or less. Consequently, the subhuman primate, such as the cynomolgus monkey, may be a less reliable model than the rat when attempting to define a role for peripheral MAO-A in drug development studies. Brain levels of MAO-A in the subhuman primates also were very low. All rodent brains, however, displayed higher MAO-A activity, with the rat being the highest at 25%. Unfortunately, we do not have human brain MAO-A and -B values for comparison at this time. According toKalaria et al. (1988), different regions of human brain contain up to 20% MAO-A.
In general, residual activities were either very low (less than a few percentage points of the total activity) or undetectable. The rabbit, however, was an exception. The MAO-A activity in rabbit liver preparations was below levels of detection, but about 27% of the total activity remained after preincubation with both clorgyline and (R)-deprenyl (Fig. 9). Brain preparations displayed about 7% MAO-A activity, but again, under conditions that inhibited over 95% of all activity in other species, residual brain activity (18%) in the rabbit remained high. This behavior has been confirmed with both sets of pooled rabbit brain and liver mitochondrial preparations. The same levels of residual activities were observed even when the protein concentrations were halved, suggesting that the residual activity is not due to the presence of exceptionally high concentrations of MAO-B. The factors responsible for this behavior have not been identified. Because these residual activities were not affected by deferoxamine, it is unlikely that adventitious iron was responsible as has been reported for the oxidation of MPTP in the presence of glial cells (Di Monte et al., 1995). The possibility that semicarbazide-sensitive amine oxidase contributed to this residual activity, as reported by Gómez et al. (1988) to account for some of the amine oxidase activity observed in rat liver, can be ruled out because the semicarbazide-sensitive amine oxidase catalytic pathway proceeds via an azomethine intermediate that cannot be formed from these tertiary amines (Holt et al., 1998). In ongoing studies, researchers are attempting to provide a more detailed characterization of rabbit liver and brain MAO activity.
The values listed in Table 1 reflect maximum velocities and do not provide an estimate of catalytic efficiency that is better expressed byVmax/Km. A more complete picture of the total (MAO-A plus MAO-B) enzyme activity profiles for the four substrates selected in this study is provided below. We have estimated theVmax/Km values for the human liver mitochondrial enzyme-catalyzed oxidation of the MAO-A and MAO-B mixed substrates (analogs 5 and 6) for both forms of the enzyme (Table 2). The results show that even thoughVmax for MAO-B is almost four times higher than it is for MAO-A for the pyrrolyl compound 5, the lowerKm value for MAO-A suggests that the overall efficiencies at low substrate concentrations may be about the same for the two enzymes. The furanyl substrate 6 is a much poorer MAO-A substrate in terms of Vmax, suggesting that the B form of the enzyme would be the principal in vivo catalyst for this substrate.
Species-Dependent MAO Kinetic Profiles Observed with Various Tetrahydropyridinyl Substrates.
An inspection of Table 3 leads to several observations. Liver activity was higher than brain activity in all animals examined except for the C57BL/6 mouse, the only animal with a liver/brain ratio ofVmax/Km of less than 1 (range, 0.3–0.7). The particularly low value ofVmax/Km for the oxidation of MPTP by liver mitochondria is principally due to the highKm value (520 μM) for this substrate. It may be reasonable to speculate that the increased susceptibility of the C57BL/6 mouse to the neurotoxic effects of MPTP compared with other rodents may be due in part to the limited systemic detoxification of this compound by liver MAO-B.
A second observation of interest was the very low MAO activity (Vmax/Km = 6–33 nmol/min-mg protein/mM) observed for the rabbit brain mitochondrial preparations. These low values reflect particularly lowVmax values, which, in the case of MPTP, was only 0.2 nmol/min-mg protein. The corresponding values for the other rodent brain activities examined ranged from about 1 to 2.4, whereas the values for the other species ranged from 1.3 (dog) to about 2.0 (subhuman primates). The rabbit liver activity also was low for MPTP compared with the other species with the single exception of the C57BL/6 mouse. A comparison of ratios of theVmax/Km values showed that the rabbit (0.2) and C57BL/6 (0.1) liver enzymes are particularly poor catalysts for MPTP relative to the human enzyme. The human-to-animal ratios for the other species vary from 0.7 (ICR mouse) to 1.4 (baboon). The exceptionally low values of this ratio for the rabbit and C57BL/6 mouse should be kept in mind when interpreting systemic versus central metabolic dispositions of MAO substrates.
A final unexpected outcome of these studies was the exceptionally highVmax values observed for bovine liver mitochondria. For example, the Vmax value with MPTP as substrate was 12.9 nmol/min-mg protein versus a range of 1.2 (rabbit) to 6.3 (baboon) nmol/min-mg protein for the other species studied. Because the Km value also was relatively high (190.6 μM), theVmax/Km value (68 nmol/min-mg protein/mM) was similar to the values observed with other species. Many of the mechanistic studies reported in the literature for MAO-B have relied on the bovine liver preparation reported by Salach and Weyler (1987). A better understanding of this exceptional behavior could prove of value when attempting to evaluate to what extent the mechanistic insights gained with the bovine liver enzyme may reflect the catalytic pathway of MAO-B present in other species.
Summary.
Perhaps the most significant finding of these comparative studies is the greater similarities in the MAO activity profiles between humans and rodents (particularly the rat) than that between humans and subhuman primates. The livers obtained from baboons and monkeys were essentially devoid of MAO-A activity, whereas the human and rat preparations were relatively rich in MAO-A activity, although, as with all tissues examined, MAO-B also was the dominant enzyme present in these species. A comparison of the substrate profiles among the various species documented many similarities. The ratios ofVmax/Km were higher in the liver (on a per-mg protein basis) than in the brain in all species except the C57BL/6 strain of mouse. This difference was not due to lower Km values for the liver enzymes. The factors contributing to these tissue-dependent differences in activity remain to be identified. Similar studies with other MAO-containing tissues (gut, heart, lung, platelet) would help to identify potentially important tissue- and species-dependent differences in the interactions of xenobiotics with these flavoenzymes. The relatively weak MAO-B activity observed in the liver mitochondria isolated from the C57BL/6 mouse, which is due primarily to the very high Km value for the liver enzyme, opens the possibility of limited systemic detoxification of MAO-B substrates in this strain of mouse. Other significant differences in MAO activity identified by these studies include the unusually low MAO-B activity displayed by brain and liver mitochondrial preparations isolated from the New Zealand White rabbit. These low activities were coupled with a resistance to inactivation by (R)-deprenyl. At the other end of the spectrum was the exceptionally robust activity of the bovine liver preparations, which reflects primarily the highVmax values for the substrates examined. Overall, the results of this effort should form the basis for future studies that will help to further identify the tissue- and species-dependent interactions of xenobiotics with MAO-A and -B.
Acknowledgments
We thank Dr. Douw G. Van der Nest, Antoinette Fick, and Cor J. J. Bester at the Experimental Animal Center and our other colleagues at Potchefstroom University for Christian Higher Education, South Africa, for assistance with baboon mitochondria preparations; Dr. David Moore, David Gemmell, and the staff of the Laboratory Animal Resources, Virginia Tech for their assistance in obtaining all rodent tissues; and Dr. Jim Bowen, Dr. Spencer Johnston, and Ann Clapsaddle at the Virginia–Maryland Regional College of Veterinary Medicine for generously providing the Beagle dog tissues. We also thank Lisa Hazelwood for her technical assistance with mitochondrial preparations and enzyme assays.
Footnotes
-
Send reprint requests to: Dr. Neal Castagnoli, Jr., Department of Chemistry, Virginia Tech, Blacksburg, VA 24061-0212. E-mail: ncastagnoli{at}chemserver.chem.vt.edu
-
↵1 This work was supported by Pharmacia & Upjohn, The Nitto Foundation, Japan, and the Harvey W. Peters Research Center for the Study of Parkinson’s Disease and Other Disorders of the Central Nervous System.
- Abbreviations:
- MAO
- monoamine oxidase
- MPDP+
- 1-methyl-4-phenyl-2,3-dihydropyridinium species
- MPP+
- 1-methyl-4-phenylpyridinium species
- MPTP
- 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
- GC
- gas chromatography
- MS
- mass spectrometry
- DMSO
- dimethyl sulfoxide
- UV-VIS
- ultraviolet-visible
- Received May 4, 1999.
- Accepted July 15, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}