


Abstract
Cyclosporin A (CyA) toxicity is a common occurrence in pediatric organ transplant patients. We hypothesized that reduced mdr1a expression in newborn and developing mice would affect CyA accumulation within organs and/or toxicity. For functional studies, CyA was administered (5 mg kg–1 i.p.) to 1-, 12-, and 19-day, and adult male and female mdr1a+/+ and mdr1a–/– mice. Peak blood CyA was lower in 1-, 12-, and 19-day-old (1000 ng ml–1) versus adult (1500 ng ml–1) mice but was similar in mdr1a+/+ and mdr1a–/– mice. Kidney mdr1a expression (measured by quantitative polymerase chain reaction) increased 2.5-fold in 19-day-old male and female mice and increased another 4-fold in adult females compared with adult males. Liver mdr1a expression increased 6-fold by day 12 compared with neonatal mice. Thereafter, maintenance of hepatic mdr1a expression in females and a reduction to neonatal levels in males was observed. Kidney/blood (8- to 9-fold) and liver/blood (12- to 15-fold) CyA levels were highest on days 12 and 19 and were not dependent on maturational changes in mdr1a mRNA levels. Adults had higher brain expression of mdr1a mRNA (3-fold), a corresponding 5-fold increase in immunodetectable P-glycoprotein, and 80% lower brain accumulation of CyA compared with 1-day-old mice. Conversely, in mdr1a-null mice, brain/blood CyA was similar in newborn and adult mice. A similar pattern was observed for the brain accumulation of the mdr1a substrate 3H-digoxin. We conclude that the risk for central nervous system drug toxicity could be higher in neonates or young children as a consequence of underdeveloped P-glycoprotein.
The genes ABCB1 (MDR1) in humans and abcb1a (mdr1a) and abcb1b (mdr1b) in mouse encode the drug efflux transporter protein P-glycoprotein (P-gp). P-gp is normally expressed in endothelial or epithelial cells of several organs including the brain, liver, intestine, and kidney (Gottesman and Pastan, 1993). Evidence supports a role for the P-gp transporter in the disposition or elimination of endogenous substances including corticosteroids, estrogens, and bilirubin (Karssen et al., 2001; Watchko et al., 2001; Kim and Benet, 2004). This transporter also plays a key role in the disposition and excretion of a wide array of clinically used drugs and provides protection against the accumulation of toxic xenobiotics within tissues (Schinkel et al., 1994; Mayer et al., 1996; Johnson et al., 2001). P-gp is a major component of the blood-brain drug permeability barrier, which prevents the accumulation of drugs [e.g., cyclosporin A (CyA), digoxin, dexamethasone, and antineoplastics] in the central nervous system (Cordon-Cardo et al., 1989; Schinkel et al., 1994, 1995; Beaulieu et al., 1997). One of the most striking examples of this barrier function is the 20- to 50-fold increase in CyA and digoxin accumulation that is observed in brains of adult mice completely deficient in the mdr1a gene compared with their wild-type littermates (Schinkel et al., 1995).
It is well known that maturational changes contribute to altered drug pharmacokinetics in pediatric versus adult populations (Strolin Benedetti and Baltes, 2003). In earlier studies, developmental changes in MDR1/mdr1 protein levels have been examined in mice, rats, or humans using the monoclonal antibody C219. The findings of reduced C219 immunoreactivity from those studies have suggested decreased P-gp in the brain, liver, and kidney in fetal or immature animals compared with adults (Schuetz et al., 1995; Matsuoka et al., 1999; Mahmood et al., 2001; Watchko et al., 2001). In support of the protein data, two recent studies have determined, by semiquantitative PCR, that mdr1a expression in the mouse brain and mdr1a and mdr1b expression in the rat brain, liver, and kidney increased with maturation (Tsai et al., 2002; Rosati et al., 2003). These findings suggested that underdeveloped MDR1 might reduce drug elimination or increase drug entry into the brain of neonates and young children, compared with adults. However, the earlier investigations did not determine whether reduced P-gp in neonates had functional consequences for the tissue disposition or elimination of P-gp substrates.
The major goal of this study was to perform in vivo functional studies in 1-day-old to adult mdr1a+/+ and mdr1a–/– mice to determine whether newborn and young mice had a greater propensity than adults to accumulate mdr1a substrates within blood/plasma and tissues. CyA is used extensively as an immunosuppressant agent in pediatric solid organ transplant recipients (Hoyer and Vester, 2004; Pape et al., 2004) and can produce serious toxic side effects including nephrotoxicity and neurotoxicity, which can hinder successful immunotherapy and adversely affect pediatric patient health (Jeruss et al., 1998; Pape et al., 2004; Taque et al., 2004). Based on this clinical/toxicological relevance in pediatric drug therapies, CyA was chosen as our primary substrate for the mdr1a functional studies. Digoxin is a well characterized substrate for mdr1a (Schinkel et al., 1995, 1997; Mayer et al., 1996; Goralski et al., 2003) and was used as an additional substrate to confirm maturational changes in function of this transporter. Also, we have used quantitative PCR (Q-PCR) to accurately determine the importance of gender as a variable that affects the developmental expression patterns of the mdr1a and mdr1b genes in mouse brain, liver, and kidney. Our major findings have revealed that maturational expression of the mdr1a gene was gender- and organ-dependent. Most striking was our novel demonstration that a reduction in mdr1a mRNA expression was associated with decreased P-gp in the brain of neonates and allowed for greater accumulation of CyA and digoxin within that organ. If P-gp follows a similar maturational expression profile in the human brain, there could be an increased risk for central nervous system (CNS) toxicity in neonates exposed to drugs known to be P-gp substrates.
Materials and Methods
The Dalhousie University Committee on Laboratory Animals approved all experimental procedures involving mice according to the guidelines of the Canadian Council on Animal Care. FVB wild-type and mdr1a–/– mice were kept on a 12-h day/night cycle, were housed in cages lined with pine bedding, and had free access to water and Purina mouse chow. The 1-day-old pups were kept with the mother during the experimental procedures.
DNA Isolation for Gender Determination of 1-Day-Old Pups and mdr1a Genotyping. Tail clips were digested with lysis buffer that contained 0.1 M Tris (pH 8.0), 5 mM EDTA, 0.2% SDS, 0.2 M NaCl, and 100 μg/ml proteinase K for 30 min at 50°C. DNA was precipitated with isopropanol, washed with 70% ethanol, and then resuspended in 50 to 100 μl of sterile distilled water. Male gender of 1-day-old pups was determined by amplification of a 185-base pair fragment of the Y-chromosome-specific zinc finger protein (zfy). The gender of adult control mice was independently verified by comparison of anogenital distance or the presence of mammary glands. The PCR mix (25 μl) contained 4 μl of resuspended tail DNA, (NH4)2SO4 buffer, 2 mM MgCl2, 0.25 mM deoxynucleotide-5′-triphosphates, 1 μM zfy primers (Table 1), and 1.25 units of Taq polymerase. The amplification protocol consisted of an initial denaturation at 95°C for 30 s followed by 30 cycles of denaturation at 95°C for 30 s, annealing at 50°C for 30 s, and elongation at 72°C for 1 min. PCR amplification of tail DNA with wild-type and mutant-specific primers (Table 1) were used to confirm the mdr1a+/+ and mdr1a–/– genotypes according to previously published methodology (Johnson et al., 2001). The primers for mitochondrial expoxide hydrolase (mEH) amplify a 367-base pair region from mouse genomic DNA and were included in all PCRs as a gender-independent positive control. Separation of PCR products on a 2.5% 0.5× TAE (Tris-acetate-EDTA) agarose gel was used to ensure the formation of a single product at the correct size.
PCR primers
RNA Isolation and Q-PCR Analyses. Total RNA was isolated from the brain, liver, and kidneys of 1-, 12-, and 19-day-old and 6-week-old adult male and female FVB wild-type mice using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Total RNA (5 μg) was reverse-transcribed using StrataScript Reverse Transcriptase (Stratagene, La Jolla, CA) with 12.5 ng/μl random hexamers pd(N)6 and 1.0 mM deoxynucleoside-5′-triphosphates according to the supplier's instructions. One microliter of the cDNA product was amplified by Q-PCR using a 125 nM concentration of gene-specific primers (Table 1) in a total volume of 20 μl with Brilliant SYBR Green QPCR Master Mix (Stratagene) using the MX3000p thermocycler (Stratagene). The amplification protocol consisted of a 10-min hot start at 94°C, followed by 35 cycles of denaturation at 94°C for 15 s, annealing at 60°C for 18 s, and elongation at 72°C for 30 s. Melting curves followed by separation of PCR products on a 2.5% 0.5× TAE agarose gel were used to ensure the formation of a single product at the appropriate size. The threshold cycle (CT) values were obtained with a threshold of 3 standard deviations above background. Relative gene expression normalized to ribosome polymerase II (rpII) expression was calculated using the –2ΔΔCT method (Livak and Schmittgen, 2001). Gene expression in 1-day-old male mice was assigned an arbitrary value of 1 and was used as the reference to which all other groups were compared.
Western Blots. Isolation of crude membrane fractions from brain, kidney, and liver and separation of proteins by SDS-polyacrylamide gel electrophoresis were performed according to previously published methods (Jette et al., 1996). The Lowry assay (Lowry et al., 1951) was used to determine total protein content, and the membrane fractions were stored at –80°C. Crude membranes were diluted in 2× Laemmli buffer and heated to 37°C for 5 min. Ninety micrograms (brain and kidney) or 30 μg (liver) of total protein was separated by electrophoresis on a 7.5% polyacrylamide gel and then transferred overnight to nitrocellulose membrane. The blots were blocked in 3% skim milk-Tris-buffered saline with Tween 20 (TBS-T, 0.1%) for 1 h at room temperature, and then incubated with the monoclonal antibody C219 (Idlabs, London, ON, Canada) (1:125) in 3% skim milk-TBS-T (0.1%) for 2 h at room temperature, followed by anti-mouse horseradish peroxidase-conjugated secondary antibody (1:25,000) in 3% skim milk-TBS-T (0.1%) for 1 h at room temperature. Immunoreactivity was detected by incubation with Fluorescent ECL-plus reagent (GE Healthcare, Little Chalfont, Buckinghamshire, UK) and visualized directly with a Storm 840 phospor imager (GE Healthcare).
CyA and 3H-Digoxin Tissue Distribution. Sandimune i.v. (cyclosporine, 50 mg/ml; Novartis, Basel, Switzerland) was diluted 1:100 in a sterile 5% glucose solution. 3H-Digoxin (1 μg ml–1) was made up in 5% sterile glucose solution and contained 1.75% ethanol (v/v) and 3.25% Cremofor EL (v/v). Mice were injected i.p. with a 5 mg kg–1 dose of the diluted CyA solution or a 10 μg kg–1 dose of the 3H-digoxin solution. The total volume of injection was proportional to weight (10 μl/g body weight). After 2 h, mice were anesthetized with enflurane and were killed by decapitation. Trunk blood, brain, liver, and kidney were collected for CyA and digoxin determination. Tissues were weighed, rinsed, and homogenized in phosphate-buffered saline (pH 7.4) with a Polytron homogenizer.
CyA Determination. The protocol for CyA determination was based on previously established methodology (Volosov et al., 2001). Blood samples (1 ml) were collected into microcentrifuge tubes containing 3.2 mg of EDTA and were stored at –20°C until preparation for liquid chromatography-mass spectrometry. CyA was extracted from 100 μl of whole blood or 100 μl of brain, liver, and kidney homogenate. Due to the small blood volume obtained from 1-day-old mice, the extractions were performed from 10 μl of whole blood diluted to 100 μl in phosphate-buffered saline. One hundred microliters of zinc sulfate (0.1 M) was added to each sample and vortexed for 5 s. Then, 300 μl of acetonitrile containing the internal standard ascomycin (30 ng/ml) was added to precipitate blood and tissue protein. Samples were vortexed for another 10 s and centrifuged for 10 min in a microcentrifuge (10,000g), and the supernatants were transferred to a clean vial. Thirty five microliters of each sample was then separated on a Zorbax SB-C18 analytical column (4.6 × 75 mm), 3.5-micron particle size, at 70°C isocratically at a flow rate of 1.6 ml/min. The mobile phase for separations contained 65% methanol and 35% ammonium acetate for 4 min. The ammonium adducts were then eluted into the tandem mass spectrometer with 100% methanol for 1 min 50 s (1.4 ml min) and quantified utilizing the heated nebulizer (APC1) source and the multiple reaction monitoring scan mode of the tandem mass spectrometer (AP1 3000) (Applied Biosystems/MDS Sciex, Concord, ON, Canada). The ammonium adducts, induced by ammonium acetate, of CyA and internal standard ascomycin are collected by the first quadrupole (Q1) and their characteristic products ions are collected by the third quadrupole (Q3). The peak height of the ion transitions was used to quantify the CyA levels by comparing to a calibration curve. Data analysis was performed by Analyst software version 1.3.1 [Applied Biosystems/MDS Sciex (Foster City, CA) 2003].
3H-Digoxin Determination. For 3H-digoxin measurement, 50 μl of plasma, 100 μl of liver and kidney homogenate, and 200 μl of brain homogenate (in duplicate) were added to 4 ml of Ready Safe scintillation cocktail (Beckman Coulter, Fullerton, CA) and counted for total dpm using a Beckman-Coulter LS5000TA scintillation counter.
Data Analysis. All groups represent the mean ± S.E.M. of at least three to five mice. A two-way analysis of variance (ANOVA) was used to measure the effect of the main variables (age and gender) and the interaction of those variables on the expression of mdr1a and mdr1b in the brain, liver, and kidney. A three-way ANOVA was used to measure the effect of age, gender, and mdr1a genotype and the interaction of those variables on the blood and tissue distribution of CyA and digoxin. An ANOVA with a p value ≤0.05 was considered significant. A Tukey's multiple comparison test was used for post hoc analysis of the significant ANOVA.
Drugs and Chemicals. Sandimune i.v. (Novartis) was obtained from the IWK Health Centre Pharmacy. 3H-Digoxin (250 μCi) was obtained from PerkinElmer Life and Analytical Sciences (Boston, MA). Oligonucleotide primers were synthesized by Sigma-Genosys Canada (Missisauga, ON, Canada). All other reagents were of the finest grade available and were purchased from commercial suppliers.
Results
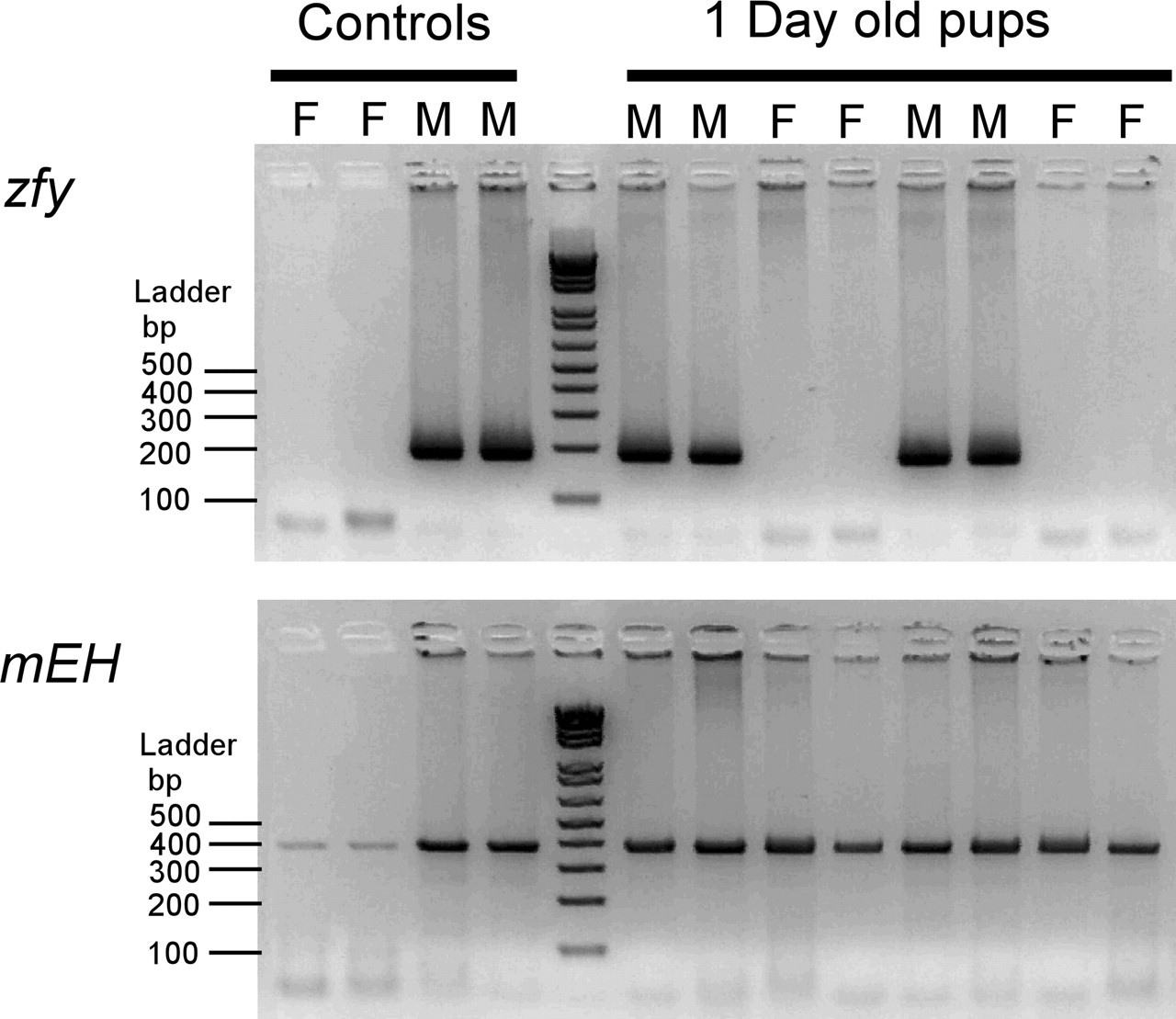
We have examined the effect of gender and maturation and the interaction of those variables on the levels of mdr1a and mdr1b genes in the mouse brain, liver, and kidney. Due to the inherent difficulties of gender determination by morphological examination of newborn pups, we have developed a PCR-based assay for unequivocal sex determination. This assay is based upon amplification of a 185-base pair PCR fragment from the Y-chromosome-specific gene (zinc finger protein, zfy). As shown in the representative gender-typing assay (Fig. 1), the zfy PCR product was detectable in adult male but not female mice. For the 1-day-old pups, the presence of the PCR product indicated male gender, whereas the absence of the PCR product indicated female gender. As a gender-independent control, a fragment of mEH DNA was amplified from male and female mice in a separate PCR.

Representative figure showing PCR gender typing of 1-day-old mice. A, the detection of the 185-base pair PCR product of the Y-chromosome-specific gene indicates male (M) gender. The absence of the zfy product indicates female (F) gender. B, as a gender-independent control, a fragment of mEH DNA was amplified from 1-day-old male and female mice in a separate PCR.
The expression of mdr1a mRNA normalized to the control gene rpII in the brain was dependent on age but not on gender (Fig. 2A). Mdr1a mRNA in the brain was at its lowest level in 1-day-old mice but increased with maturation to a level 2.5- to 3-fold higher in 19-day-old and 6-week-old mice. Conversely, brain mdr1b mRNA levels did not increase significantly with age and were also not affected by gender (Fig. 2B). The ratio of mdr1a/mdr1b expression increased from 70 in newborn to 130 in adult mice and indicated that the mdr1a gene was the primary expressed P-gp isoform in all age groups (Fig. 2C). The level of the control gene rpII was similar for both genders and in all age groups (Fig. 2D).
In the kidney, 1-day-old mice had the lowest relative expression of mdr1a mRNA (Fig. 3A). In 19-day-old male and female mice, mdr1a mRNA levels increased 2.5-fold compared with the 1-day-old mice. In adult female, but not male mice, mdr1a mRNA was increased another 4-fold and revealed gender-dependent expression in adult mice. A similar gender differential was observed for mdr1b mRNA levels in adult female versus male kidney (Fig. 3B). The mdr1a and mdr1b genes were expressed to a similar degree in the kidney (Fig. 3C). The level of the control gene rpII was similar in kidneys from males and females and in all age groups (Fig. 3D).
Similar to the condition in brain and kidney, 1-day-old male and female mice had the lowest levels of mdr1a mRNA in the liver (Fig. 4A). The mdr1a mRNA level then increased 6- to 8-fold in 12-day-old male and female mice compared with the newborn mice. Thereafter, mdr1a mRNA levels declined in male mice and remained elevated in female mice. This resulted in a 5-fold higher expression of mdr1a mRNA in the liver of adult female compared with adult male mice. Mdr1b seemed to be primarily a neonatal expressed gene in the liver since its expression was highest in 1-day-old mice (Fig. 4B). The amount of mdr1b mRNA declined by 75% in the liver of 12- and 19-day-old mice. In adult mice, the levels of hepatic mdr1b remained 50 to 60% lower than observed in newborn mice. Unlike mdr1a, the level of mdr1b mRNA in the liver was not dependent on gender. In all age groups, liver mdr1a levels were substantially higher than mdr1b levels (Fig. 4C). The difference (300- to 800-fold) was most predominant in the 12- to 19-day-old mice. The level of the control gene rpII was similar in liver from male and female mice and in all age groups (Fig. 4D).

The effect of maturation and gender on brain expression of mdr1a and mdr1b mRNA. The relative expression of mdr1a (A) and mdr1b (B) was determined by Q-PCR. The ratio of mdr1a to mdr1b threshold cycle CT values in the brain is shown in C. For each group, mdr1a and mdr1b expression was normalized to the control gene rpII (D). The 1-day-old male mice were chosen as the reference group (expression = 1.0) to which all other groups were compared. Each bar represents the mean ± S.E.M. of three to four mice. Data were compared by a two-way ANOVA followed by Tukey's HSD post hoc analysis of the significant ANOVA. ‡, p < 0.05, significantly higher gene expression (independent of gender) as compared with the 1-day-old mice.
Western blots on crude membranes obtained from brain, liver, and kidney of 1-day-old and adult mice confirmed that maturational and gender changes in mdr1a expression correlated with P-gp expression (Fig. 5). In the brain of 1-day-old mice, P-gp expression was about 20% of the levels observed in adult mice. Similar to mdr1a mRNA expression, no gender difference was observed for P-gp expression in the brain. P-gp was not detectable in the kidney membrane fractions prepared from 1-day-old mice but was detectable in the kidney membrane fractions prepared from adult mice. Consistent with mdr1a mRNA expression in that organ, P-gp expression was substantially higher in the adult females compared with males. In crude membrane fractions prepared from the liver, P-gp was detectable in 1-day-old male and female mice. The level of immunodetectable P-gp decreased in adult males and did not increase further in adult females. The gender difference in liver P-gp expression in adult mice was consistent with the gender difference in liver mdr1a mRNA expression.
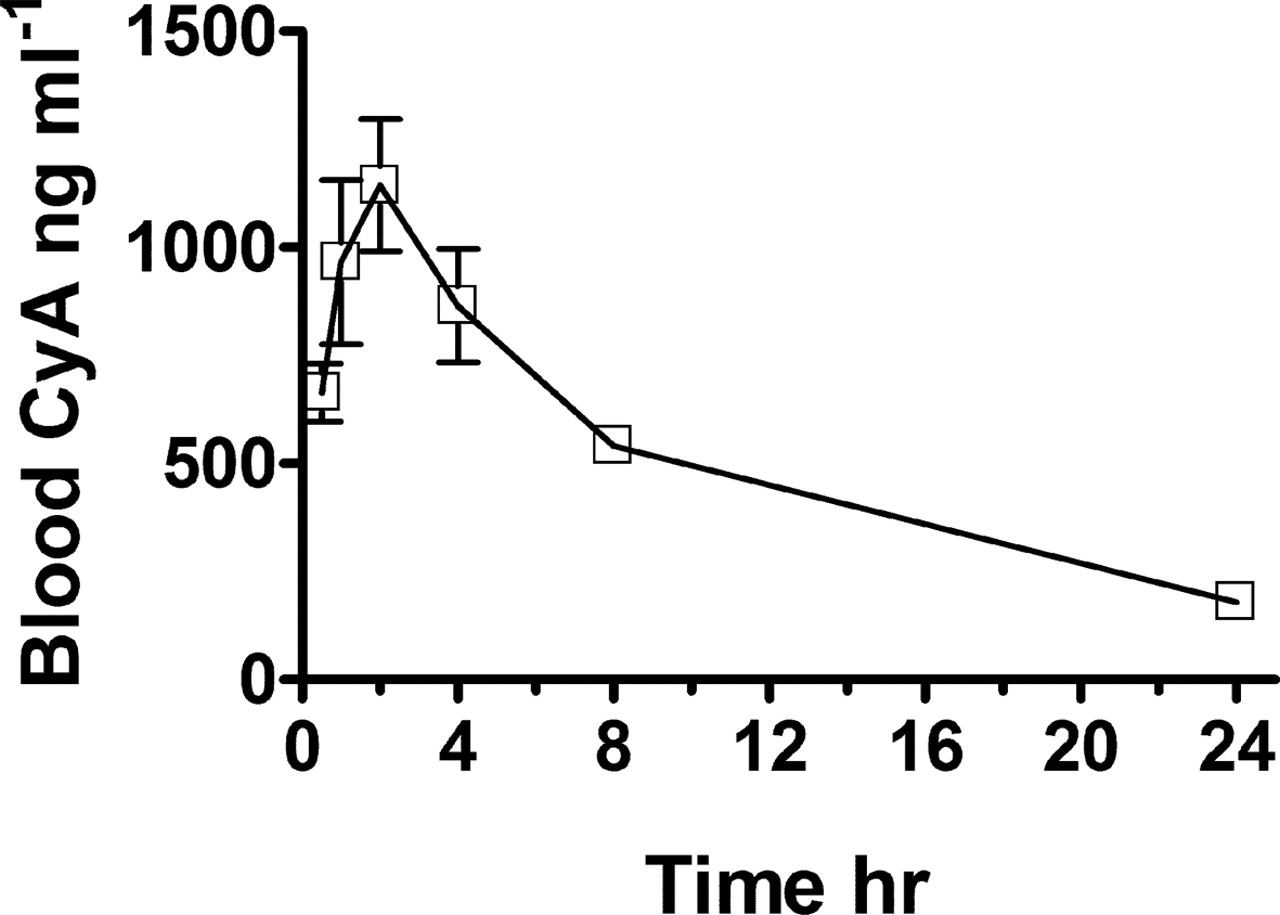
To establish conditions for the functional studies, we performed an initial experiment in which adult male wild-type mice were administered a therapeutically relevant dose (5 mg kg–1) of CyA by i.p. injection. Similar to the kinetics of CyA in humans (Jorga et al., 2004), peak drug levels (1000–1200 ng ml–1) in mouse blood were obtained 2 h after dosing, and 24-h trough levels were in the range of 100 to 200 ng ml–1 (Fig. 6). For subsequent experiments, blood and tissue cyclosporine concentrations were measured 2 h after dosing (C2). We have also validated the CyA analysis for detection of that drug in tissue samples because the assay procedures were initially developed for the detection of CyA in blood (Table 2). Thus, CyA was added to brain, liver, and kidney homogenates to yield low and high CyA concentrations that corresponded to the range of expected CyA levels in those tissues. The average measured CyA concentrations in brain, liver, and kidney were within 5% of the predicted values. The lower limit of detection was 500 pg (5 ng/ml) of CyA per 100 μl of homogenate. Thus, the mouse served as an appropriate model for examining CyA kinetics, and the described HPLC-tandem MS assay provides a highly sensitive method for determining blood and tissue CyA in this species.
Validation of CyA measurements in tissue homogenates
CyA was added to tissue homogenates to achieve a final concentration of 10, 100, or 1000 ng/ml. Samples were then extracted and CyA was measured by HPLC-MS/MS to validate measurement of that drug in the tissue homogenates.
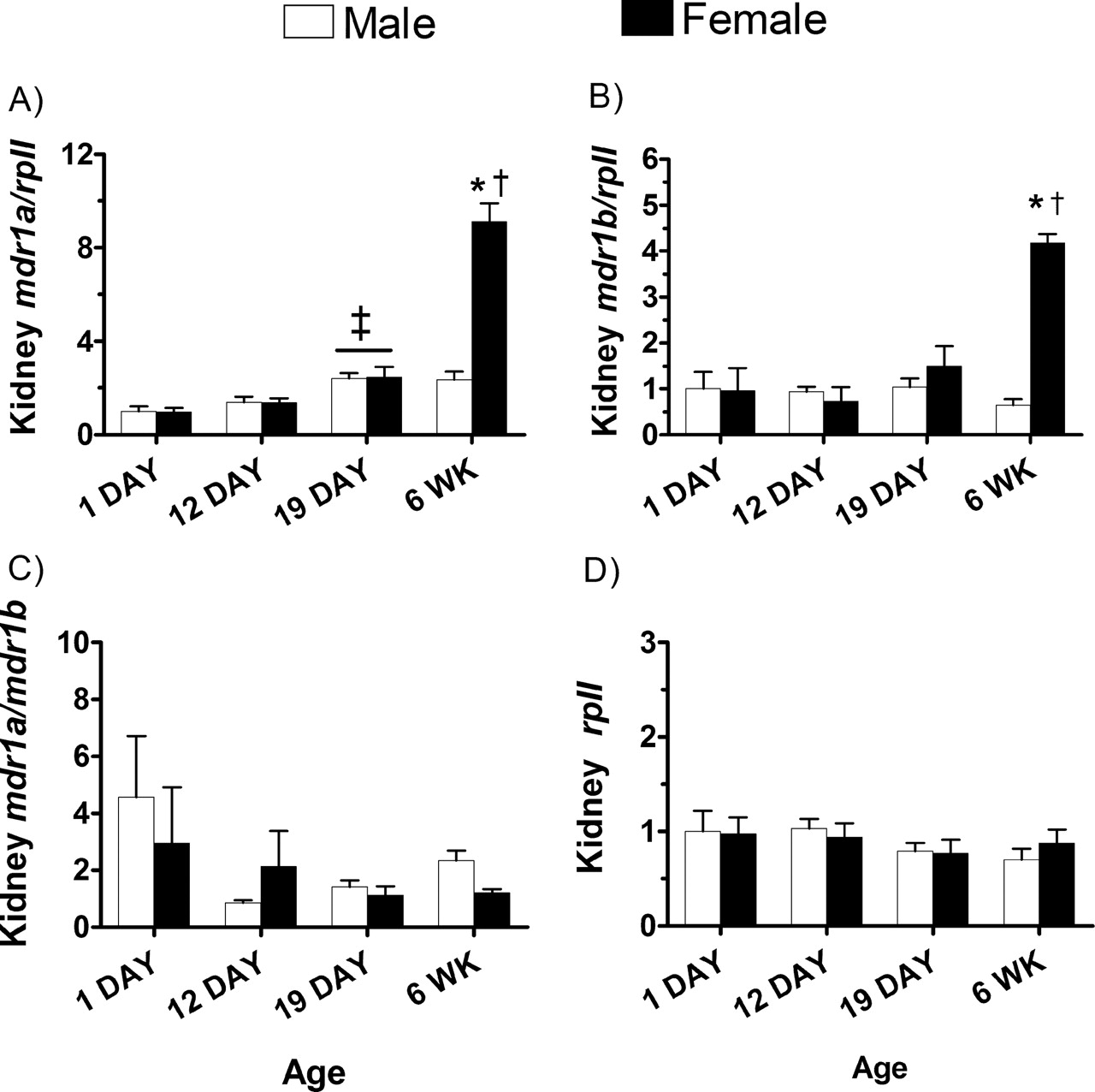
The effect of maturation and gender on kidney expression of mdr1a and mdr1b and mRNA. The relative expression of mdr1a (A) and mdr1b (B) was determined as described in the legend to Fig. 2. The ratio of mdr1a to mdr1b expression in the kidney and the expression of the housekeeping gene rpII are shown in panels C and D, respectively. Each bar represents the mean ± S.E.M. of three to four mice. Data were compared by a two-way ANOVA followed by Tukey's HSD post hoc analysis. ‡, p < 0.05, significantly higher gene expression (independent of gender) as compared with the 1-day-old mice. *, p < 0.05, significantly higher gene expression in adult females compared with all other age groups. †, p < 0.05, significantly higher gene expression in adult female compared with adult male mice.
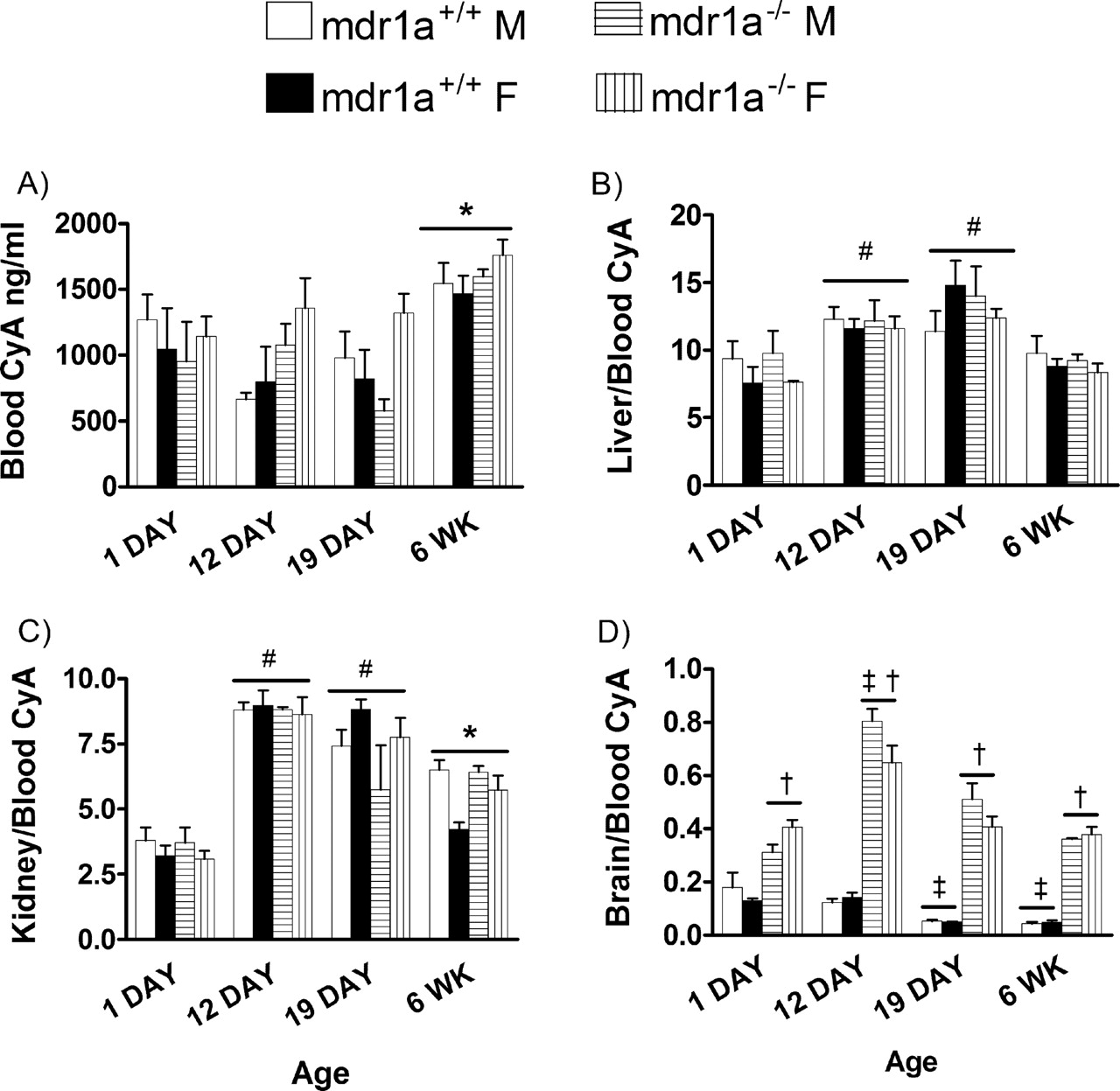
To confirm mdr1a-dependent effects and to reveal mdr1a-independent effects, the effects of ontogeny on CyA disposition were examined in mdr1a-expressing and mdr1a-null mice. Furthermore, within each age group, the mdr1a–/– mice served as a reference point for complete loss of mdr1a function. Blood CyA C2 values were on average one-third lower in 1-, 12-, and 19-day-old mice compared with adult mice but were not dependent on gender (Fig. 7A). C2 blood CyA was similar in mdr1a+/+ and mdr1a–/– mice at all ages and indicated that peak blood CyA after i.p. dose was not influenced by mdr1a mRNA levels at any age. CyA was highly concentrated in liver (8- to 12-fold) and kidney (4- to 8-fold) compared with the blood compartment (Fig. 7, B and C). The accumulation of CyA in the liver and kidney was elevated in the 12- and 19-day-old mice. However, this effect was not due to maturational changes in mdr1a mRNA levels since mdr1a+/+ and mdr1a–/– mice had similar accumulation of CyA in those organs in all age groups studied. Furthermore, the increased level of mdr1a in the liver and kidney of adult female mice was not associated with a decrease in CyA accumulation. In the brain (Fig. 7D), CyA accumulation was inversely related to age in wild-type mice. The 1-day-old mice had 3- to 4-fold higher brain/blood concentration ratios than adult wild-type mice, which was consistent with the pattern of mdr1a mRNA and P-gp expression during maturation. Conversely, brain/blood CyA was similar in 1-day, 19-day, and adult mdr1a–/– mice with a marked increase in brain/blood CyA in the 12-day-old mdr1a–/– mice.

The effect of maturation and gender on liver expression of mdr1a and mdr1b mRNA. The relative expression of mdr1a (A) and mdr1b (B) was determined as described in the legend to Fig. 2. The ratio of mdr1a to mdr1b expression in the liver and the expression of the housekeeping gene rpII are shown in C and D, respectively. Each bar represents the mean ± S.E.M. of three to four mice. Data were compared by a two-way ANOVA followed by Tukey's HSD post hoc analysis. ‡, p < 0.05, significantly lower gene expression compared with 1-day-old mice (independent of gender). *, p < 0.05, significantly higher gene expression as compared with the 1-day-old-male or female mice. †, p < 0.05, significantly higher gene expression in adult female compared with adult male mice.

Maturation and gender affect P-gp expression. Crude membrane fractions were prepared from brain, liver, and kidney of 1-day-old and adult male and female mice. P-gp expression in these tissues was determined by Western blot analysis using the antibody C219.
To provide further support for maturational changes in function of mdr1a, a tissue distribution study was performed under the same conditions, with the mdr1a substrate 3H-digoxin (Fig. 8). Similar to CyA, the plasma 3H-digoxin dpm measured 2 h after dosing was comparable in 1- to 19-day-old male and female mdr1a+/+ and mdr1a–/– mice (Fig. 8A). An effect of the mdr1a gene deletion became unmasked in adult mice, wherein the mdr1a–/– mice had 2-fold higher plasma digoxin compared with the wild-type male and female controls. The liver/plasma digoxin ratio peaked (6-fold) in 12-day-old mice, whereas the kidney/plasma ratio was higher in 1- and 12-day-old mice and decreased thereafter (Fig. 8, B and C). However, the maturational effect on digoxin accumulation in the liver and kidney in the 1- to 19-day-old mice was similar in male and female mice of both genotypes, indicating that the change was not gender- or mdr1a-dependent. Although not significant, the liver/plasma and kidney/plasma 3H-digoxin levels tended to be higher in the mdr1a–/– mice compared with the mdr1a+/+ mice. Comparable to the functional studies with CyA, 3H-digoxin accumulation in the brain of mdr1a+/+ mice was 5 times higher in 1-day-olds compared with adults and was consistent with the maturational increase in mdr1a mRNA and P-gp levels in that organ. In the mdr1a–/– mice, digoxin accumulation in the brain was similar in 1-day-old compared to adults and peaked in the 12-day-old mice. The -fold difference of CyA or 3H-digoxin accumulation in the brain of mdr1a–/– mice versus the age-matched mdr1a+/+ mice was lowest in 1-day-olds and increased with maturation, an indication that the mdr1a transporter is functionally active in newborn mice, albeit at a reduced level compared with adults (Table 3).
-Fold difference in brain/blood (CyA) and brain/plasma (digoxin) accumulation between mdr1a-/- and mdr1a+/+ mice of different ages
The -fold difference in CyA of mdr1a-/-/mdr1a+/+ = (brain/blood CyA mdr1a-/-/(brain/blood CyA mdr1a+/+). The -fold difference in digoxin dpm of mdr1a-/-/mdr1a+/+ = (brain/blood digoxin mdr1a-/-)/(brain/blood digoxin mdr1a+/+). Because there was no statistical difference between male and female mice, the data from these groups were combined.

Blood CyA levels versus time after dosing. Six-week-old male mice received a single dose of Sandimune (5 mg kg–1) by i.p. injection. Blood samples (10–15 μl) were taken from the end of the tail of each mouse at 0.5, 1.0, 2, 0, 4.0, 8.0, and 24 h after the CyA dose. Each symbol represents the mean ± S.E.M. of three mice. The standard errors are too small to be observed for the 8-h and 24-h time points.
Discussion
We present a novel characterization of gender- and age-dependent regulation of mdr1a and mdr1b mRNA levels and function of the P-gp transporter in the mouse brain, liver, and kidney. In mice and humans, P-gp expression is an early marker of blood-brain barrier development (Qin and Sato, 1995; Schumacher and Mollgard, 1997). In a previous report, antibody C219-detectable P-gp was 5-fold higher in the brains of adult FVB mice compared with newborn mice (Watchko et al., 2001; Tsai et al., 2002). We have confirmed a similar maturational increase in brain P-gp. This increase in brain P-gp was most likely attributable to brain mdr1a expression, which also increased postnatally and was 3-fold higher in adult compared with 1-day-old mice. Mdr1a is expressed (qualitatively) to a higher degree than mdr1b in the adult mouse brain (Croop et al., 1989; Schinkel et al., 1994). Similar to adult mice, our results have established that mdr1a is the predominant P-gp isoform expressed in newborn and maturing mice. The higher level of mdr1a compared with mdr1b mRNA in the adult mouse brain is consistent with functional data, whereby deletion of the mdr1b gene did not result in increased brain accumulation of the P-gp substrate digoxin (Schinkel et al., 1997).

The effect of maturation on the blood and tissue distribution of CyA. The 1-, 12-, and 19-day and 6-week male and female mdr1a+/+ and mdr1a–/– mice were administered 5 mg kg CyA by i.p. injection. Blood (A), liver/blood (B), kidney/blood (C), and brain/blood (D) CyA levels were measured 2 h later. Each bar represents the mean ± S.E.M of three to four mice. Data were compared by a three-way ANOVA followed by Tukey's HSD post hoc analysis of the significant ANOVA. *, p < 0.05, significantly different as compared with all other age groups (independent of gender or genotype). †, p < 0.05, significantly higher in mdr1a–/– mice compared with mdr1a+/+ mice of the same age (independent of gender). ‡, p < 0.05, significantly different compared with 1-day-old mice of the same genotype (independent of gender). #, p < 0.05, significantly different compared with 1-day-old mice (independent of gender or genotype).
The phenotype of increased brain accumulation and/or toxicity of P-gp substrates, including CyA, digoxin, dexamethasone, ivermectin, vincristine, and vinblastine, has been documented in adult mdr1a–/– mice (Schinkel et al., 1994, 1995; Johnson et al., 2001). However, the level of protection provided by mdr1a against drug accumulation in the brains of newborn mice has not been examined. Based on our observations of decreased mdr1a mRNA levels and P-gp, newborn mice should be less protected than adults. Consistent with this prediction, newborn wild-type mice achieved levels of CyA and 3H-digoxin accumulation in the brain that were about 50% of the levels achieved in newborn mdr1a–/– mice that lack expression of that transporter. In contrast, the relative accumulation of CyA and 3H-digoxin in the brains of adult mdr1a+/+ mice decreased to about 10% compared with the adult mdr1a–/– mice. These results indicated that the level of protection afforded by mdr1a against drug accumulation in the brain is lowest in newborn mice and is enhanced as the expression of that gene and its product, P-gp, increases during maturation. Our novel finding that newborn mdr1a+/+ but not mdr1a–/– mice had increased brain accumulation of CyA (3-fold) and 3H-digoxin (5-fold), compared with the respective adult mice, further supports the conclusion that the reduced brain expression of mdr1a in the neonate directly contributed to the greater accumulation of those mdr1a substrates in that organ. The 12-day-old mdr1a–/– mice had a peak in brain CyA and 3H-digoxin levels, suggesting a maturational change in the blood-brain barrier permeability to drugs that is unmasked only when mdr1a expression is completely absent. This change may reflect maturational expression patterns of other drug transporters (e.g., oatp2, mrp1, mrp2, and bcrp) that are present in the blood-brain barrier (Bauer et al., 2005).

The effect of maturation on the blood and tissue distribution of 3H-digoxin. The 1-, 12-, and 19-day and 6-week male and female mdr1a+/+ and mdr1a–/– mice were administered 10 μg kg–1 3H-digoxin by i.p. injection. Plasma (A), liver/plasma (B), kidney/plasma (C), and brain/plasma (D) dpm levels were measured 2 h later. Each bar represents the mean ± S.E.M of three to four mice. The groups were compared by a three-way ANOVA followed by Tukey's HSD post hoc analysis. *, p < 0.05, significantly different as compared with all other age groups (independent of gender or genotype). †, p < 0.05, significantly higher in mdr1a–/– mice compared with mdr1a+/+ mice of the same age group (independent of gender). ‡, p < 0.05, significantly different compared with 1-day-old mice of the same genotype (independent of gender). #, p < 0.05, significantly different compared with 1-day-old mice (independent of gender or genotype). **, p < 0.05, significantly different compared with adult male or female wild-type mice.
Use of calcineurin inhibitors (CyA and tacrolimus) for suppression of organ rejection in pediatric renal transplant patients can precipitate CNS toxicity, behavioral disturbances, and interruptions in immunosuppressive therapy (Awan et al., 1999; Parvex et al., 2001; Yamauchi et al., 2002; Taque et al., 2004). Approximately 10% to 15% of pediatric renal transplant patients will exhibit significant clinical CNS toxicity characterized by post-transplant seizures (Awan et al., 1999). Although the etiology of CNS toxicity is multifactorial, young age and drug inhibition of P-gp function in the brain have been implicated (Jeruss et al., 1998; Awan et al., 1999). Our data further suggest that the risk for adverse CNS toxicity may be higher in young patients as a consequence of not yet fully developed P-gp. Consistent with this hypothesis, a G2677T mutation in exon 21 of the human MDR1 transporter has been associated with a greater frequency of adverse CNS effects during immunotherapy with tacrolimus (Yamauchi et al., 2002).
Biological differences between males and females are well known to affect drug pharmacokinetics (Schwartz, 2003). We have identified for the first time that the developmental pattern of mdr1a mRNA expression in the liver and kidney is highly dependent on gender. In females, expression of the mdr1a gene increased in liver (5-fold) and kidney (6-fold) with age and was maintained in adulthood. In males, kidney expression increased 2.5-fold with maturation, whereas hepatic mdr1a expression was maximal in 12-day-old males and declined thereafter. Overall, females maintained a 4- to 5-fold higher mdr1a expression in the kidney and liver as compared with adult males. Similarly, adult females maintained higher levels of P-gp expression in the kidney and liver as compared with males. This pattern of expression is not restricted to the mdr1a transporter since similar gender-dependent expression of the oatp2 transporter mRNA or protein occurs in rat or mouse liver during maturation (Guo et al., 2002; Cheng et al., 2005). A previous study reported that mdr1a mRNA expression in rat liver and kidney increased minimally (2- to 3-fold) during maturation (Rosati et al., 2003). In comparison to our results, those findings could reflect a species difference in developmental expression of the mdr1a transporter. Furthermore, the gender of the rats was not identified or included as a variable in the earlier study by Rosati et al. (2003) and could also contribute to the differences in findings. In line with the gender-related expression difference of mdr1a mRNA and P-gp in the kidney, adult male mice tended to have higher plasma and kidney levels of 3H-digoxin as compared with the adult wild-type female mice. Consistent with the findings of an earlier study (Schinkel et al., 1994), we also demonstrated a gender-dependent (female > male) expression of the mdr1b isoform in the kidney but not the liver. Higher expression levels of liver and kidney drug transporters such as P-gp may be required for disposition and elimination of female sex steroids. This idea is supported by a recent study which showed that estradiol, estrone, and estriol were transported substrates and inducers of MDR1 expressed in MDCK cells (Kim and Benet, 2004). Although not the focus of this study, female sex hormones could contribute to the increased hepatic and kidney mdr1a expression in female mice. Unlike the kidney and liver results, our findings show that the brain levels of mdr1a or mdr1b genes and P-gp or accumulation of CyA and digoxin in that organ were not gender-dependent. Those results are supported by earlier findings that showed gender independence of verapamil, morphine, and quinidine uptake into the brain of adult mice (Dagenais et al., 2001).
In our functional studies, we observed that 12- and 19-day-old mice had higher liver/blood and kidney/blood CyA levels than did newborn and adult mice. The increase in organ CyA accumulation could be especially important in the kidney, because one of the main side effects of therapy with that drug in children is nephrotoxicity (Pape et al., 2004). A recent study has demonstrated that MDR1 3435TT genotype of the donor kidney was a major risk factor for CyA nephrotoxicity and was not explained by CyA dose or blood levels of the drug (Hauser et al., 2005). These data indicate that increased kidney levels of CyA might be associated with certain MDR1 polymorphisms. In comparison with the toxicological data from that human study, our functional studies in mice indicated that maturational or gender changes in mdr1a were not a limiting factor for determining peak CyA levels in the blood, or accumulation of that drug in the liver and kidney. However, it remains to be determined whether a lower level of mdr1a expression decreases elimination of CyA from the kidney or liver over time, resulting in higher tissue trough levels.
Within each of the younger age groups (1-, 12-, or 19-day-old), CyA or 3H-digoxin accumulation in the liver and kidney was similar in wild-type versus mdr1a-null mice. This similarity indicates that changes in drug accumulation, in those organs, in young mice could not be explained by maturational changes in mdr1a mRNA levels. However, a phenotype of increased plasma, liver, and kidney 3H-digoxin but not CyA began to appear in adult mdr1a–/– mice as compared with the wild-type mice. Schinkel et al. (1995) demonstrated similar results for CyA and digoxin in adult mdr1a+/+ versus mdr1a–/– mice 4 or 8 h after dosing with those drugs. Thus, the effect of mdr1a expression on blood and tissue disposition of P-gp substrates was age-, organ-, and drug-dependent.
In summary, we have demonstrated that maturation of mdr1a expression in the mouse liver and kidney is dependent on gender. Importantly, our functional studies have demonstrated that CyA and 3H-digoxin accumulation in the brain was highly dependent upon mdr1a expression: in contrast, accumulation of these drugs in the liver and kidney was not affected significantly by the reduced mdr1a expression levels in these organs of newborn mice. Considering these findings, it will be important to address whether the risk for CNS toxicity due to increased drug levels in brain is higher in young patients as a consequence of decreased MDR1 expression and function.
Acknowledgments
Technical assistance was provided by Gordon Nash.
Footnotes
-
This work was supported by operating grants from the Canadian Institutes of Health Research. K.B.G. was supported by postdoctoral fellowships from The IWK Health Centre, The Canadian Institutes of Health Research, and the Reynolds Foundation.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.007427.
-
ABBREVIATIONS: P-gp, P-glycoprotein; CyA, cyclosporin A; PCR, polymerase chain reaction; Q-PCR, quantitative PCR; CNS, central nervous system; zfy, Y-chromosome-specific zinc finger protein; mEH, mitochondrial expoxide hydrolase; CT, threshold cycle; rpII, ribosome polymerase II; TBS-T, Tris-buffered saline with Tween 20; ANOVA, analysis of variance; C2, cyclosporine concentration measured 2 h after dosing; HSD, honestly significantly different.
- Received September 16, 2005.
- Accepted November 9, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}