


Abstract
In this study, we investigated whether luteolin monoglucuronide was converted to free aglycone during inflammation using human neutrophils stimulated with ionomycin/cytochalasin B and rats treated with lipopolysaccharide (LPS). β-Glucuronidase activity was assayed using 4-methylumbelliferyl-glucuronide and methanol extracts of rat plasma containing luteolin monoglucuronide. The released 4-methylumbelliferone, a fluorescent molecule, was quantified by fluorometry. Deglucuronidation of luteolin monoglucuronide was examined by high-performance liquid chromatography (HPLC) analysis. HPLC analyses showed that the supernatants obtained from neutrophils stimulated with ionomycin/cytochalasin B hydrolyzed luteolin monoglucuronide to free luteolin. β-Glucuronidase activity in human serum from patients on hemodialysis increased significantly compared with that from healthy volunteers. The β-glucuronidase activity in rat plasma increased after i.v. injection of LPS. The ratio of luteolin to luteolin monoglucuronide in plasma of LPS-treated rats also increased. These results suggest that during inflammation β-glucuronidase is released from stimulated neutrophils or certain injured cells and then deglucuronidation of flavonoids occurs.
There have been many studies reporting that flavonoids absorbed in vivo are metabolized to glucuronide or sulfate conjugates, which are polar and water-soluble. These metabolites circulate in the blood and are excreted into bile and urine (Scalbert and Williamson, 2000). The glucuronidation process is very important for detoxification of xenobiotics. It has been unclear whether the flavonoid glucuronides can function in vivo like flavonoid aglycones. The glucuronides have less antioxidant activity than aglycones (Manach et al., 1995). Although aglycones are thought to enter cells by passive diffusion, it is not clear whether glucuronides can enter cells. Deglucuronidation of flavonoids has been reported to occur in the human large intestine (Heneghan, 1988). The conjugates of flavonoids excreted into bile can be hydrolyzed to aglycones by intestinal bacteria, which have β-glucuronidase and arylsulfatase. However, β-glucuronidase, which is a lysosomal enzyme, is also present in mammalian cells and is known to be released from neutrophils and eosinophils during inflammation (Marshall et al., 1988).
Luteolin has been shown to have antioxidant (Shimoi et al., 1994), antitumorigenic (Yasukawa et al., 1989), and anti-inflammatory/antiallergic (Yamamoto et al., 1998) activities and to be an inhibitor of protein kinase C (Ferriola et al., 1989) and lipoxygenase (Yamamoto et al., 1998). Our previous studies have demonstrated that luteolin has a radioprotective effect and a protective effect on doxorubicin-induced cardiotoxicity in mice (Shimoi et al., 1994; Sadzuka et al., 1997). HPLC1 and liquid chromatography/mass spectrometry analyses of rat plasma after administration of luteolin showed that the main metabolite was luteolin monoglucuronide and that free luteolin was also present. Luteolin monoglucuronide and free luteolin were also detected in human serum after ingestion of luteolin (Shimoi et al., 1998).
In the present study, we used luteolin as a model compound and investigated whether luteolin monoglucuronide is converted to free luteolin during inflammation in in vitro reaction with human neutrophils stimulated with ionomycin/cytochalasin B and in LPS-treated rats.
Materials and Methods
Chemicals.
Luteolin (Fig. 1) was isolated from perilla seed, purified by HPLC, and confirmed by mass spectrometry and NMR spectral data at the Oryza Oil and Fat Chemical Co., Ltd. (Ichinomiya, Japan). All other chemicals were of reagent grade.

Chemical structures of luteolin and 4-methylumbelliferone.
Neutrophils.
Human neutrophils were isolated from venous blood of two healthy volunteers, who gave informed consent before the experiment, by using standard dextran sedimentation and gradient separation on Lymphoprep (NYCOMED, Oslo, Norway). Contaminating erythrocytes were lysed by 0.6 M KCl treatment. Neutrophils were >95% pure, and viability exceeded 98% as assessed by trypan blue dye exclusion. Purified neutrophils (5.4 or 7.6 × 106 cells) were resuspended in saline G (200 μl). They were stimulated with 5 μg/ml cytochalasin B (Sigma, St. Louis, MO) for 5 min at 37°C and further treated with 1 μM ionomycin (Sigma) for 20 min at 37°C. After centrifugation for 10 min at 4°C at 750g, the supernatant was obtained.
Treatment of Rats with LPS.
Male SD rats (7–9 weeks old; SLC Inc., Hamamatsu, Japan) weighing 180 to 200 g were housed in an air-conditioned room with free access to CE-2 commercial food pellets (Clea, Tokyo, Japan). The diet was changed to a synthetic basic diet, which consisted of 38% corn starch, 25% casein, 10% α-starch, 8% cellulose powder, 6% minerals, 5% sugar, 2% vitamins, and 6% linoleic acid (Oriental Yeast Co., Tokyo, Japan) 1 week before the experiment. Three rats were assigned to each experimental group. Animals were maintained and handled according to the Guidelines for the Regulation of Animal Experimentation Committee of the University of Shizuoka. The rats fasted overnight, received an i.v. injection of LPS (Escherichia coli 0111:B4; Difco, Detroit, MI) at a dose of 750 μg/kg, and were anesthetized with ethyl ether 4.5 h after injection. The rat blood was withdrawn from the abdominal aorta into EDTA-treated tubes, and the plasma was prepared by centrifugation for 10 min at 900g for β-glucuronidase assay. Luteolin (50 μmol/kg) was administered to rats by gastric intubation 3.5 h after LPS injection, and the plasma was prepared 0.5, 1, and 4.5 h after oral administration of luteolin for HPLC analysis.
Sample Preparation of Human Serum.
Venous blood of patients on hemodialysis and healthy volunteers who gave informed consent was collected in vacuum containers early in the morning at the Social Insurance of Chukyo Hospital (Nagoya, Japan). The serum was prepared by centrifugation for 10 min at 900g.
β-Glucuronidase Assay.
β-Glucuronidase activity in rat plasma and human serum was assayed using the FluorAce β-glucuronidase reporter assay kit (Bio-Rad, Hercules, CA). Briefly, 50 μl of assay buffer and 10 μl of sample were added to each sample well of a 96-well microplate. The microtiter plate was incubated at 37°C for 30 min. β-Glucuronidase hydrolyzes 4-methylumbelliferyl-glucuronide resulting in release of the fluorescent molecule 4-methylumbelliferone (Fig. 1). The fluorescence of this molecule was measured on a microplate fluorometer using an excitation wavelength of 360 nm and emission wavelength of 460 nm.
To know whether β-glucuronidase released from the stimulated human neutrophils is able to hydrolyze flavonoid glucuronides to free aglycone, supernatants, which were obtained from the culture medium after neutrophils from two human healthy volunteers had been stimulated with or without ionomycin/cytochalasin B, were reacted with methanol extracts of plasma of rats orally administered luteolin at 37°C for 2 h. Methanol extracts of plasma were obtained by the method described previously (Shimoiet al., 1998). In brief, rat blood was withdrawn from the abdominal aorta into heparinized tubes after administration of luteolin (50 μmol/kg in propyleneglycol) by gastric intubation. The plasma (0.5 ml) was acidified with the same volume of 0.01 M oxalic acid. This solution was applied to a Sep-Pak C18 cartridge. After washing the cartridge with 0.01 M oxalic acid, methanol/water/0.5 M oxalic acid (25:73:2, v/v/v), and distilled water, the methanol eluate was obtained. The eluate was evaporated to dryness. The reaction mixture of the supernatant and methanol extract dissolved in a small amount of ethanol was acidified with the same volume of 0.01 M oxalic acid. This solution was applied to a Sep-Pak C18 cartridge, and then the methanol eluate, obtained by the same method as mentioned above, was evaporated to dryness. The residue dissolved in methanol was subjected to HPLC. The ratio of luteolin to luteolin monoglucuronide was determined by the peak areas on HPLC chromatograms.
HPLC Analysis.
HPLC analysis was carried out by the method described previously (Shimoi et al., 1998). Briefly, the samples were analyzed chromatographically by a JASCO HPLC system (JASCO, Inc., Tokyo, Japan) using a Capcell Pak C18-UG120 column (150 × 4.6 mm; Shiseido, Tokyo, Japan) and UV detection (349 nm). Elution was performed using methanol with 0.03% trifluoroacetic acid (solvent A) and water with 0.01% trifluoroacetic acid (solvent B) in gradient conditions at the flow rate 0.7 ml/min.
Statistical Analysis.
Statistical analysis was performed using Student's t test. Any difference between the two groups with a value of P< 0.05 was considered significant.
Results and Discussion
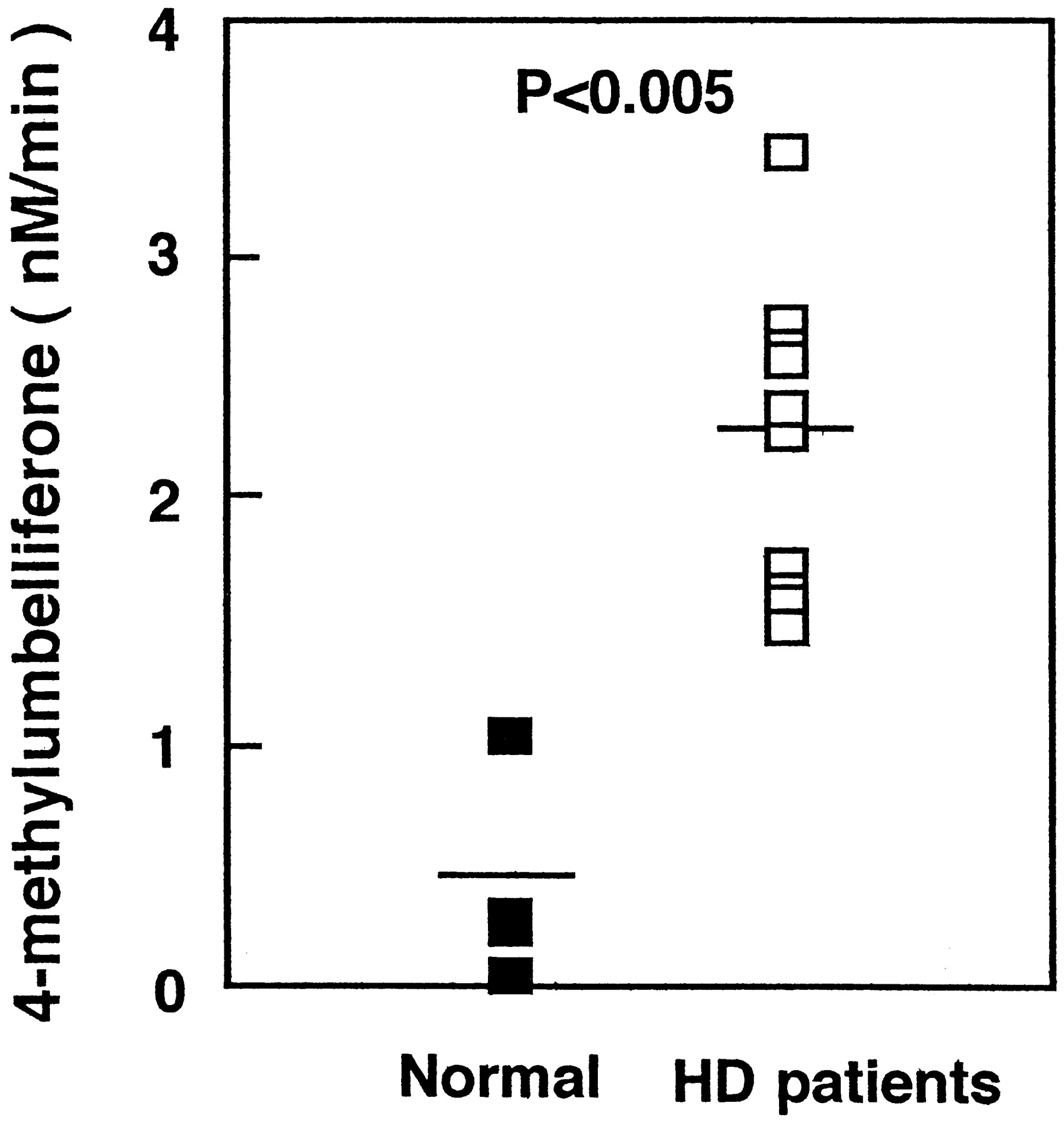
β-Glucuronidase, which hydrolyzes glucuronides to liberate free aglycones, is located in the lysosomes and in endoplasmic reticulum (Wakabayashi, 1970; Paigen, 1989). This enzyme plays an important role in the hydrolysis of estrogen glucuronides (Zhu et al., 1996) and oligosaccharides (Buddecke and Hoefele, 1966). Moderate oxidative stress was found to rapidly induce partial lysosomal rupture, which was followed by apoptosis and further loss of intact lysosomes. The release of hydrolytic enzymes from the lysosomal compartment to the cytosol is an important initiating event in the apoptotic process (Brunk Ulf, 1999). β-Glucuronidase activity in both the liver and kidney increases with age perhaps because increasing fragility of lysosomal membrane allows these enzyme to leak out (Cristofalo and Kabakijiam, 1975). In inflammation, β-glucuronidase is known to be released from granulocytes including neutrophils (Marshall et al., 1988). It has been reported that serum levels of proinflammatory cytokines such as interleukin-1 and C-reactive protein, circulating markers of inflammation, were increased in patients on hemodialysis (Docci et al., 1990; Pereira, 1995). Therefore, we measured β-glucuronidase activity in serum from patients on hemodialysis and healthy volunteers. As shown in Fig. 2, β-glucuronidase activity in serum from patients on hemodialysis increased significantly to 2.26 ± 0.66 nM/min (mean ± S.D.) compared with that from healthy volunteers, 0.42 ± 0.44 nM/min (mean ± S.D.) (P < 0.005).
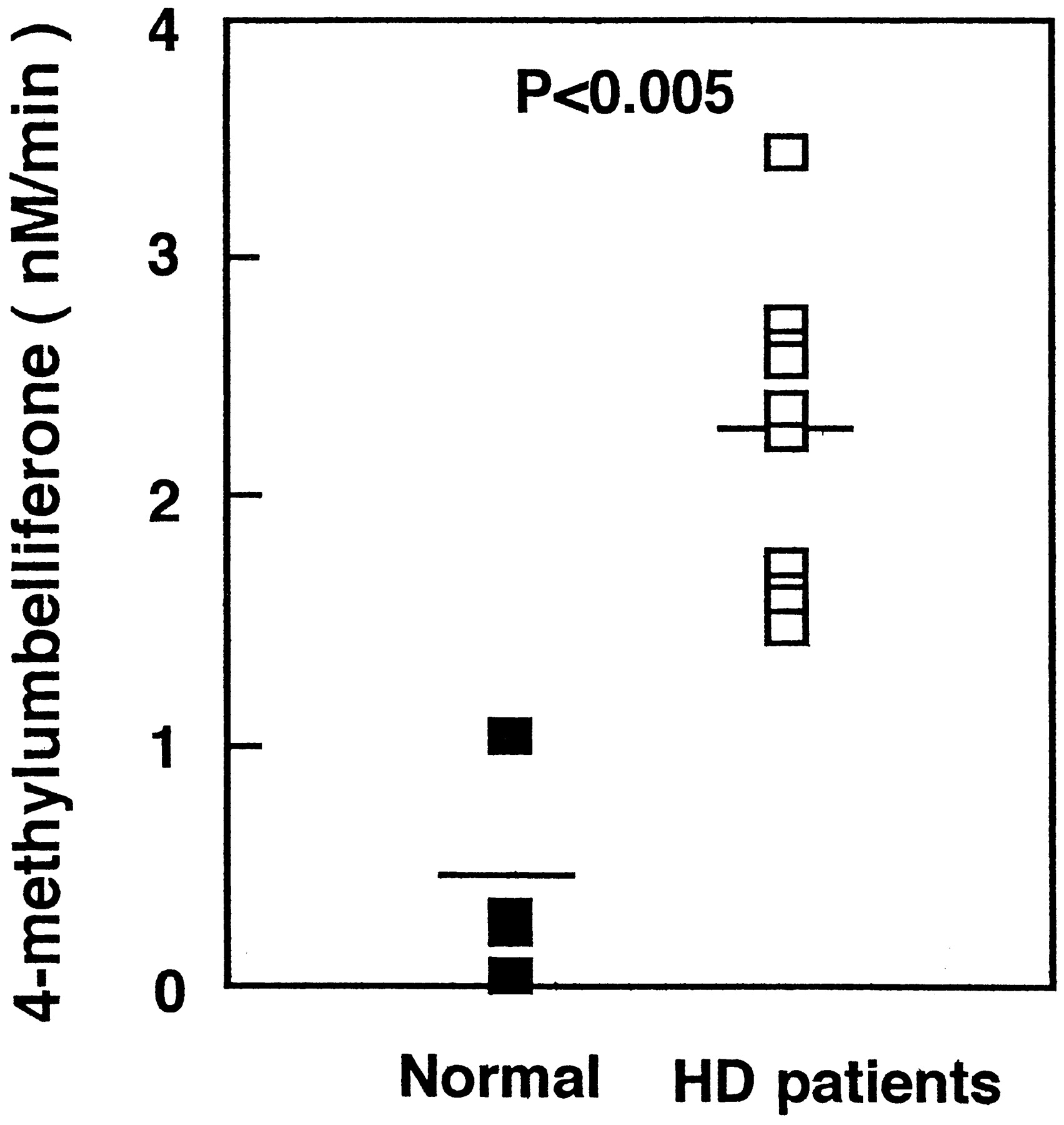
β-Glucuronidase activity in serum of healthy volunteers (n = 4) and patients on hemodialysis (n = 10).
The serum was prepared by centrifugation for 10 min at 900g. The β-glucuronidase activity in each plasma was assayed by liberation of 4-methylumbelliferone from 4-methylumbelliferyl-glucuronide.
Supernatants, which were obtained from the culture medium after neutrophils from two human healthy volunteers who had been stimulated with or without ionomycin/cytochalasin B, were incubated with the methanol extract of rat plasma containing luteolin and luteolin monoglucuronide. Deglucuronidation activity in the supernatants was investigated by HPLC analysis. Figure 3A shows the HPLC chromatogram of the methanol extract of rat plasma used in this assay. The ratio of luteolin to luteolin monoglucuronide in the methanol extracts after treatment with the supernatants from stimulated neutrophils increased from 0.44 to 1.73 and 3.69, respectively (Fig.3B). These results indicate that luteolin monoglucuronide can be a substrate for β-glucuronidase released from neutrophils.

A, HPLC chromatogram of the methanol extract of rat plasma 30 min after oral administration of luteolin (50 μmol/kg).
Peak a, luteolin monoglucuronide; peak b, luteolin. B, the ratio of luteolin to luteolin monoglucuronide of the methanol extract of plasma of rat orally administered luteolin after treatment with or without the supernatant that contains β-glucuronidase released from the stimulated human neutrophils. Neutrophils were isolated from venous blood of two healthy volunteers (1, 5.4 × 106 cells; 2, 7.6 × 106 cells). The reaction mixtures were analyzed by HPLC. The ratio of luteolin to luteolin monoglucuronide was determined by the peak areas on HPLC chromatograms.
The β-glucuronidase activity in rat plasma after LPS treatment increased significantly from 1.38 ± 0.86 nM/min (mean ± S.D.) to 10.5 ± 3.0 nM/min (mean ± S.D.) (P< 0.005; Fig. 4A). The ratio of luteolin to luteolin monoglucuronide in plasma of LPS-treated rats also increased after luteolin administration (Fig. 4B). A significant increase was observed 0.5 and 1 h after luteolin administration [0.23 ± 0.02 to 0.78 ± 0.26 (mean ± S.D.), P < 0.005; 0.25 ± 0.03 to 0.74 ± 0.41 (mean ± S.D.), P < 0.01, respectively]. No difference in the plasma concentration of luteolin was observed between LPS-treated rats and nontreated rats. However, the concentration of luteolin monoglucuronide decreased by about half in the LPS-treated rats (data not shown). Although there is a possibility that β-glucuronidase for deglucuronidation is more active than UDP-glucuronyltransferase for glucuronidation in the liver when inflammation occurs, the results suggest that β-glucuronidase released from human granulocytes, including neutrophils or certain injured cells, appears in the blood or body fluids and hydrolyzes glucuronide conjugates of flavonoids to free aglycones at the inflammation site. The tissue fluid at the inflammation site has a low pH, which is favorable for β-glucuronidase, because β-glucuronidase exhibits activity at a very low pH (pH 4–5).

A, β-glucuronidase activity in plasma of rats treated with or without LPS.
The blood was collected from the abdominal aorta into EDTA-treated tubes 4.5 h after i.v. injection of LPS (750 μg/kg). β-Glucuronidase activity was assayed with 4-methylumbelliferone, as described in the legend to Fig. 2. The values are mean ± S.D. (n = 3; ∗∗∗, P < 0.005). B, the ratio of luteolin to luteolin monoglucuronide in plasma of LPS-treated rats at different times after luteolin administration. Luteolin (50 μmol/kg) was administered to rats by gastric intubation 3.5 h after LPS injection. The ratio of luteolin to luteolin monoglucuronide was determined by the peak areas on HPLC chromatograms. The values are mean ± S.D. (n = 3; ∗∗∗,P < 0.005; ∗∗, P < 0.01).
Manach et al. (1998) have reported that the conjugated derivatives of quercetin significantly delayed the Cu2+-induced oxidation of lipoproteins. However, the magnitude of this inhibition was about one-half that measured with the aglycone. It has been demonstrated that quercetin-conjugated metabolites, such as quercetin-3′ (or 4′)-glucuronide, inhibit xanthine oxidase and lipoxygenase at 0.25 μM (Ki). This concentration is within the expected range after a meal rich in flavonols (Manach et al., 1998; Day et al., 2000). However, it is not clear at the present time whether or not the cells take up the flavonoid glucuronides.
In inflammatory responses such as those which are involved in the pathogenesis of atherosclerosis, cancer, rheumatoid arthritis, diabetic nephropathy, etc., neutrophils generate O⨪2 and release lysosomal enzymes such as β-glucuronidase. Because the released β-glucuronidase can hydrolyze glucuronide conjugates of flavonoids to free aglycones, flavonoids in the free form become more antioxidant. They come into the cells and exert biological effects at inflammation sites. The flavonoids might play a role in the prevention of diseases mentioned above.
Footnotes
-
This work was supported by Special Coordination Funds of the Ministry of Education, Culture, Sports, Science, and Technology, the Japanese Government, and a grant from The Danone Institute of Japan. We thank Oryza Oil and Fat Chemical Co., Ltd. for providing luteolin.
- Abbreviations used are::
- HPLC
- high-performance liquid chromatography
- LPS
- lipopolysaccharide
- Received March 12, 2001.
- Accepted August 20, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}