
Abstract
Imaging the competition between D2/3 radioligands and endogenous dopamine is so far the only way to measure dopamine release in the living human brain. The dopamine D2 receptor exists in a high (D2high) and a low-affinity state for dopamine. Under physiological conditions, dopamine is expected to bind to D2high only. [11C]-(+)-4-propyl-9-hydroxynaphthoxazine ((+)-PHNO) is the first D2/3 agonist radioligand for positron emission tomography (PET) imaging in humans. Since [11C]-(+)-PHNO is expected to bind preferentially to D2high, it should be particularly vulnerable to competition with endogenous dopamine. Nine healthy subjects participated in two PET scans, one after administration of d-amphetamine and one after placebo. [11C]-(+)-PHNO PET test re-test variability was determined in 11 healthy subjects. Binding potentials (BPs) were calculated for caudate, putamen, ventral striatum, and globus pallidus. d-Amphetamine led to a significant decrease of [11C]-(+)-PHNO BPs in caudate (−13.2%), putamen (−20.8%), and ventral striatum (−24.9%), but not in globus pallidus (−6.5%). d-Amphetamine-induced displacement correlated with serum d-amphetamine levels in all regions but caudate. This is the first report on competition between endogenous dopamine and a D2/3 agonist radioligand in humans. [11C]-(+)-PHNO PET might be a superior measure for release of endogenous dopamine than PET employing conventional D2/3 antagonist radioligands.
Similar content being viewed by others

INTRODUCTION
Measuring changes in radioligand binding caused by fluctuations in synaptic dopamine is at present the only way to draw inference on changes in dopamine concentrations in the living human brain. Reductions in radioligand binding to dopamine D2/3 receptors measured with positron emission tomography (PET) or single photon emission computer tomography (SPECT) have been shown after several behavioral (Koepp et al, 1998; de la Fuente-Fernandez et al, 2002; Pruessner et al, 2004; Zald et al, 2004; Volkow et al, 2006) and pharmacological manipulations (Farde et al, 1992; Volkow et al, 1994; Tedroff et al, 1996; de la Fuente-Fernandez et al, 2004), which raise endogenous dopamine levels. In particular, the well-replicated finding of greater reductions in [11C]-raclopride and [123I]IBZM binding after d-amphetamine administration in patients with schizophrenia when compared to healthy controls (Laruelle et al, 1996, 1999; Breier et al, 1997; Abi-Dargham et al, 1998) has generated considerable interest in this research strategy. A critical shortcoming of these studies is the ceiling-effect found with conventional antagonist-radiotracers (Laruelle, 2000), and although competitive inhibition of radioligand binding at the D2/3 receptor by dopamine is believed to be one of the main mechanisms underlying reductions in radioligand-binding, the exact nature of the process is still not fully elucidated (for review see Ginovart, 2005).
It is well established in vitro that D2 receptors exist in two interconvertible affinity states for their natural agonist dopamine, the high-affinity state (D2high; Kd for dopamine 1.5±0.2 nM) and the low-affinity state (D2low; Kd for dopamine in the micromolar range; Sibley et al, 1982). Since D2high mediates signal transduction at the postsynaptic neuron, the high-affinity state is believed to be the functionally important one (Zahniser and Molinoff, 1978; George et al, 1985; Leff, 1995). As a result of the large difference in Kd values between D2high and D2low, under physiological conditions, dopamine is expected to bind to D2high only. So far, all D2/3 radioligands used in human PET or SPECT studies are radio-labelled antagonists, which do not differentiate between D2high and D2low. Several research groups have recently reported on development and experimental use of newly developed D2/3 agonist radioligands (Zijlstra et al, 1993a, 1993b; Shi et al, 1999, 2004; Hwang et al, 2000; Mukherjee et al, 2000, 2004; Finnema et al, 2005; Narendran et al, 2004). Similar to dopamine, D2/3 agonist radioligands are expected to bind mainly to D2high. As a consequence, they should be particularly sensitive to competition with endogenous dopamine. This has recently been confirmed in experiments in pigs (Cumming et al, 2003), rodents (Cumming et al, 2002), non-human primates (Narendran et al, 2004; Seneca et al, 2006), and cats (Ginovart et al, 2006a).
(+)-4-Propyl-9-hydroxynaphthoxazine, (+)-(PHNO), is a full agonist at D2/3 receptors (Brown et al, 1997). Labelled with carbon-11 (Wilson et al, 2005), it is a PET ligand with excellent signal-to noise ratio and favorable kinetics for PET imaging in humans (Ginovart et al, 2006b; Willeit et al, 2006). As [11C]-(+)-PHNO is expected to bind to D2high only, in theory, all binding sites should be vulnerable to competition with endogenous dopamine, and its binding should be reduced to a considerably larger extent than binding of an antagonist-radioligand by pretreatment with d-amphetamine, a potent releaser of central nervous dopamine. In a PET study performed in cats, Ginovart et al (2006a) have shown not only that [11C]-(+)-PHNO is more vulnerable towards d-amphetamine effects than the D2/3 antagonist radioligand [11C]raclopride, but also that [11C]-(+)-PHNO binding is reduced to an even larger extent than binding of the D2 agonist ligand [11C]NPA.
In this study, we aimed to investigate the effects of endogenous dopamine on striatal [11C]-(+)-PHNO binding in healthy human subjects undergoing two [11C]-(+)-PHNO PET scans, one at placebo conditions and another one after pretreatment with d-amphetamine. Test–retest variability of [11C]-(+)-PHNO PET was determined in an independent study sample.
MATERIALS AND METHODS
Study Protocol
This study consisted of two parts. Part one was designed to determine test–retest reliability of [11C]-(+)-PHNO PET imaging, part two investigated the effects of d-amphetamine on [11C]-(+)-PHNO binding. For the test-retest protocol, subjects underwent two [11C]-(+)-PHNO PET scans at drug-naive conditions at least 1 week apart from each other. For the d-amphetamine protocol, subjects underwent one [11C]-(+)-PHNO PET scan 2 h after oral intake of d-amphetamine, and another [11C]-(+)-PHNO PET scan after oral intake of a placebo. Conditions were randomly counterbalanced, scans took place at least 1 week apart from each other.
Study Subjects and Safety Procedures
This study has been approved by the local Ethics Committee and the Canadian Ministry of Health, Therapeutic Products Research Department. Twenty-three healthy volunteers (nine females, 14 males; mean age: 33±9 years, range: 18–49 years) were recruited by advertisements or word of mouth. Written informed consent was obtained after full explanation of study procedures and risks. Routine blood and urine tests, an electrocardiogram (ECG) and a physical exam were performed before inclusion. Psychiatric disorders were assessed using the MINI-Plus structured interview (Sheehan et al, 1998). Subjects with serious or unstable medical or neurological conditions, axis-one psychiatric diagnoses, substance abuse other than caffeine or nicotine within 6 months before baseline visit were not included into the study. On a day of a PET examination, smokers (n=5) were asked to consume no more than their usual amount of cigarettes, and all participants were asked to abstain from alcohol intake 24 h before PET scans, and from caffeine-containing beverages 12 h before scans. A standardized light breakfast was served before d-amphetamine/placebo intake.
Standard urine tests for psychotropic substances were performed at inclusion and before PET scans. Pregnancy was excluded using serum HCG analysis at inclusion and standard urine pregnancy tests before each scan. Blood pressure measurements and continuous ECG monitoring were performed during all scans. Participants had a physical exam and standard ECGs immediately after scans. To document safety of [11C]-(+)-PHNO PET procedures, the first 12 subjects underwent an additional physical exam, ECG, routine blood and urine analysis the day after PET scans.
Eleven subjects (one female) completed the test–retest protocol, nine subjects (five females) the d-amphetamine protocol. Three subjects (all females) dropped out during or after the first [11C]-(+)-PHNO PET scan (two because of nausea, one subject had moved away).
d-Amphetamine Administration
Two hours before radiotracer injection, participants were administered either two or three capsules containing d-amphetamine (Dexedrine® tablets, Glaxo Smith Kline, Mississauga, ON) or an equal number of identical capsules containing inactive lactose-powder. Participants and research personnel were blind to the content of the capsules. According to body weight, 25, 30, or 35 mg d-amphetamine were administered, resulting in a dose of 0.38–0.45 mg/kg body weight (mean±SD dose: 27.8±3.02 mg; mean±SD dose per kg body weight: 0.42±0.02 mg). Five millilitres of blood were drawn immediately before PET scans, centrifuged, and stored at −80°C for determination of serum d-amphetamine levels.
Subjective drug effects were measured using the Drug Effects Questionnaire (DEQ; Justice and de Wit, 2000) and stimulant-subscales of the Subjective States Questionnaire (SSQ; White et al, 2002). Both scales are visual analog-scales previously shown to be sensitive to d-amphetamine effects (Justice and de Wit, 2000; White et al, 2002). Scales were administered before d-amphetamine/placebo intake and 60 and 230 min thereafter. Heart rate and blood pressure were measured at 15, 30, 60, 90, and 180 min after d-amphetamine administration, after tracer injection, and after PET scans.
[11C]-(+)-PHNO Synthesis
Radiosynthesis of [11C]-(+)-PHNO has been described in detail elsewhere (Wilson et al, 2005). Briefly, [11C]propionyl chloride was reacted with 9-hydroxynaphthoxazine to generate a [11C]amide, which was subsequently reduced by lithium aluminium hydride. Purification by HPLC and formulation gave radiochemically pure [11C]-(+)-PHNO as a sterile, pyrogen-free solution suitable for human studies.
Image Acquisition
All PET images were acquired on a CPS-HRRT high-resolution neuro-PET camera system (Siemens Medical Imaging, Knoxville, TN) with an in-plane resolution of approximately 2.8 mm full-width at half-maximum (FWHM). Participants were scanned in supine position using a custom-made thermoplastic facemask together with a head-fixation system (Tru-Scan Imaging, Annapolis). Transmission scans were acquired before emission scans using a single photon point source, 137Cs (T=30.2 years, Eγ=662 keV) and used for attenuation correction. A saline solution of 355.2±44 MBq [11C]-(+)-PHNO with a specific activity at time of injection of 42.65±13.2 GBq/μmol was injected as a bolus into an intravenous line placed in an antecubital vein. The line was flushed with 10 ml saline immediately after tracer injection and subsequently removed. Emission data were acquired in list mode over 90 min, raw data were reconstructed by filtered-back projection to yield dynamic images with 15 1-min frames and 15 5-min frames. Proton-density (PD) magnetic resonance images (MRIs) were obtained on a General Electric Medical System Signa 1.5T MRI scanner (General Electric Medical Systems, Milwaukee, WI).
Image Analysis
All PET images were analyzed using the in-house automated image analysis software ROMI. Exact procedures used in ROMI are described elsewhere (Rusjan et al, 2006a). In brief, a PD-MRI template in Montreal Neurologic Institute/International Consortium for Brain Mapping (MNI/ICBM) standard brain space was co-registered to PD-MRI images using nonlinear iterative co-registration algorithms implemented in SPM2 (http://www.fil.ion.ucl.ac.uk/spm/). Transformation matrices were then applied to a standardized template in MNI/ICBM space containing predefined regions of interest (ROIs) for caudate (CAU), putamen (PUT), ventral striatum (VST), globus pallidus (GP), and cortical cerebellum (CER). Definition of the VST in the template followed the method of Mawlawi et al (2001). After spatial co-registration to PD-MRIs, the template was refined using gray matter probability-maps obtained from MRIs using SPM2. Since GP is imaged with a tone halfway between gray and white matter structures, a special algorithm using a predefined volume for GP (Spinks et al, 2005) was applied to refine the GP-ROI (Rusjan et al, 2006b). PD-MRIs were co-registered to summed PET images, the spatial transformation matrix was then applied to the refined ROIs. Data from both hemispheres were pooled to obtain average radioactivity concentrations in the volumes of interest. Regional radioactivity was determined for each frame, corrected for decay, and plotted vs time to obtain time–activity curves (TACs). The simplified reference tissue model (SRTM; Lammertsma et al, 1996) was applied to derive binding potentials (BPs) for each region of interest using PMOD software (Version 2.6.1; PMOD Technologies Ltd, Zurich, Switzerland). Cortical cerebellum served as reference region since it is virtually devoid of dopamine D2 and D3 receptors in humans (Camps et al, 1989; Hall et al, 1996; Levant, 1998). The use of the SRTM with a cerebellar input function has recently been validated using kinetic modelling and shown to provide adequate quantification of D2/3 receptors with [11C]-(+)-PHNO in humans (Ginovart et al, 2006b).
To validate the automated image analysis software for [11C]-(+)-PHNO, 12 PET scans of six participants were analyzed using ROMI software and in a conventional manual way. For manual analysis, MRI scans were co-registered to PET scans using Analyze 5.0 software (Biomedical Imaging Resource, Mayo Clinic, Rochester, MN). Regions of Interest for CAU, PUT, VST, GP, and CER were drawn on PD-MRI images and subsequently transferred onto PET images. Typically, five axial PET slices were used for CER (around the outermost border of cerebellar cortex, sparing midline structures), 7–8 axial slices for CAU and PUT, and 6–7 axial slices for GP. Delineation of the VST followed the method described by Mawlawi et al (2001).
For an analysis of d-amphetamine effects without a priori anatomical hypothesis, parametric maps of d-amphetamine and placebo scans were constructed using PMOD software and subsequently analyzed using SPM2. For each scan, the SRTM was applied voxelwise using CER as reference region to create parametric maps with a voxel size of 2 × 2 × 2 mm (x–y–z). A template was constructed using a mean image of naïve BP maps. Individual parametric maps were spatially normalized to the template by Nearest Neighbor interpolation algorithm. Effects of d-amphetamine administration were assessed voxel-wise using paired t-test procedures implemented in SPM2.
Statistical Analysis
Differences in the magnitude of change between placebo and d-amphetamine and between scan one and scan two in d-amphetamine/placebo and test/re-test parts of the study were analyzed using repeated measures analysis of variance (RM-ANOVA). Binding potentials in the four ROIs were the dependent variables, the repetition factor was termed ‘condition’ (ie, d-amphetamine vs placebo and scan one vs scan two). The respective study part (d-amphetamine/placebo vs test/retest) was the between-subject variable. The significance of the ‘study part*condition’ interactions are reported. Paired-samples t-tests (two-tailed) were used for post hoc comparisons. Correlations between d-amphetamine plasma levels and d-amphetamine-induced reductions in [11C]-(+)-PHNO BPs, and correlations between results obtained with ROMI software and manual image analysis were analyzed using Pearson Product Moment correlations. Paired-samples t-tests (two-tailed) were used to analyze subjective and physiological effects of d-amphetamine.
Test-retest variability was calculated as [(BPscan one−BPscan two)/BPscan one × 100], d-amphetamine-induced reductions in [11C]-(+)-PHNO BPs were calculated as the percentage reduction in BP obtained after drug treatment when compared to placebo [(BPplacebo−BPd-amphetamine)/BPplacebo × 100].
All tests were performed using the statistical software package SPSS, Release 12.0.1 (SPSS Inc., Chicago, IL).
RESULTS
Physiological Effects/Safety of [11C]-(+)-PHNO
Injection of [11C]-(+)-PHNO did not lead to any significant changes in blood pressure, heart rate, or ECG at any time in the study. Similarly, there were no relevant findings in physical or neurological exams or in routine blood and urine analyses during the study. However, as described previously (Willeit et al, 2006), participants described mild and self-limited (duration 2–3 min) side effects (slight nausea or abdominal sensations of warmth) in one-third of the scans (14 of 43 scans). One subject wished to interrupt the scan; another one had a single episode of vomiting and did not reassume scanning thereafter because of data loss. Full data sets were acquired in 20 subjects.
Physiological and Subjective Effects of d-Amphetamine Administration
In good agreement with previous data [http://us.gsk.com/products/assets/us_dexedrine.pdf], administration of d-amphetamine resulted in serum levels of 53.2±24.4 ng/ml (range: 11.6–73.8 ng/ml) two hours post d-amphetamine intake (p.i.), that is, immediately before the PET scan. There was a significant increase in systolic (baseline: 115±12 mm Hg; peak 90 min p.i.: 132±23 mm Hg) and diastolic (baseline: 73±7 mm Hg; peak 60 min p.i.: 77±13 mm Hg) blood pressure and in DEQ-ratings (baseline: 5.2±1.8; 60 min p.i.: 6.6±2.6; 230 min p.i.: 8.3±8.4) after d-amphetamine intake. Heart rate and SSQ did not differ significantly between conditions.
Imaging Results
Binding potentials obtained with the automated image analysis software ROMI and conventional manual analysis showed excellent correlations in all investigated ROIs: CAU: r=0.973, p<0.001; PUT: r=0.993, p<0.001; VST: r=0.966, p<0.001; GP: r=0.916, p<0.001.
Indicating a lack of blood-flow effects on the free and non-specific tracer compartment, TACs obtained in d-amphetamine and placebo scans for the reference region CER were congruent (Figure 1). As described in detail elsewhere (Ginovart et al, 2006b; Willeit et al, 2006), tracer kinetics in GP differed from those in CAU, PUT, and VST: TACs peaked at a lower level and showed a slower washout.

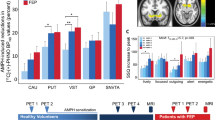
Time–activity curves (mean±SEM) for caudate, putamen, ventral striatum, and globus pallidus 2 h after oral intake of placebo (Plac; empty triangles) or d-amphetamine (Amph; filled triangles). Y-axis represents standardized uptake values (SUV; calculated as: regional radioactivity concentration/(injected radioactivity/body weight) for [11C]-(+)-PHNO. The congruent curves in cerebellum (empty diamonds: placebo; filled diamonds: d-amphetamine) indicate that administration of d-amphetamine did not have relevant influence on free and non-specific [11C]-(+)-PHNO binding in this study.
In the test–retest group, there were no significant differences in [11C](+)-PHNO BPs between scan one and scan two in any of the ROIs (paired t-test; Table 1). Using absolute, that is, unsigned values of the differences between scan one and scan two, test–retest variability of [11C]-(+)-PHNO BPs was 8.7±8% for CAU, 9.9±8% for PUT, 18.6±19% for VST, and 21.3±16% for GP, respectively.
There was a significant decrease in [11C]-(+)-PHNO BPs in d-amphetamine scans vs placebo (Table 1, Figure 2) in CAU, PUT, and VST. Changes in GP did not reach level of significance. Indicating significant differences for within-subject changes in [11C]-(+)-PHNO BPs between d-amphetamine/placebo and the test/re–test part of the study, the interaction term ‘condition*study-part’ was significant in all ROIs with exception of GP: CAU: F(1)=6.248, p=0.022; PUT: F(1)=11.637, p=0.003; VST: F(1)=5.501, p=0.031; GP: F(1)=1.125, p=0.303.

[11C]-(+)-PHNO binding potentials (BPs) in healthy subjects for the regions of interest caudate (CAU), putamen (PUT), ventral striatum (VST), and globus pallidus (GP) 2 h after intake of placebo capsules or capsules containing 27.8±3 mg (mean±SD) d-amphetamine. ***Two-tailed p⩽0.001; n.s.: two-tailed p=0.224.
Serum d-amphetamine levels correlated significantly with d-amphetamine induced reductions in [11C]-(+)-PHNO BPs in all ROIs but the CAU (CAU: r=−0.0116, p=0.77; PUT: r=0.682, p=0.043; VST: r=0.669, p=0.049; GP: r=0.948, p<0.001). Notably, an increase rather than a decrease in post-d-amphetamine [11C]-(+)-PHNO BPs in the GP measured in the three subjects with lowest serum d-amphetamine levels contributed visibly (Figure 3) to the high correlation coefficient found in this ROI.

Correlations between serum d-amphetamine levels (Amph) and reductions in [11C]-(+)-PHNO binding potentials (Δ[11C]-(+)-PHNO BPs) calculated as ((BP placebo−BP d-amphetamine)/BP placebo) × 100. Note scaling of the Y axis in scatter-plot for GP. CAU, caudate; PUT, putamen; VST, ventral striatum; GP, globus pallidus.
Binding Potential Maps
Overall appearance of parametric BP maps was similar to summated [11C]-(+)-PHNO PET images, dopamine D2/3 receptor rich regions were clearly delineated. Highest [11C]-(+)-PHNO BPs were found in GP and the ventral portion of the neostriatum. The region with statistically most significant d-amphetamine-induced displacement of [11C]-(+)-PHNO binding was a bilateral cluster located in the medio-ventral portion of the striatum (Figure 4). Peak voxels were identified at MNI coordinates x=−14, y=16, z=−6 (T=12.52, puncorrected<0.001), x=4, y=14, z=−8 (T=12.01, puncorrected<0.001), and x=18, y=16, z=−8 (T=11.71, puncorrected<0.001). The cluster followed the contours of the putamen in dorso-caudal direction (Figures 4 and 5).

Statistical parametric map showing areas of significant d-amphetamine induced reductions (paired samples t-test) in [11C]-(+)-PHNO binding potentials. Most significant displacement is found in a bilateral area located in the ventral striatum.

Colored areas show percent decrease in [11C]-(+)-PHNO binding potentials (BPs) in d-amphetamine scans as compared to placebo scans. Image represents mean image of nine individual subtraction images calculated as ((BPmap placebo−BPmap d-amphetamine)/BPmap placebo) × 100.
DISCUSSION
This study is, to our knowledge, the first to demonstrate in vivo competition of endogenous dopamine with the new D2/3 agonist radioligand [11C]-(+)-PHNO in humans. Administration of d-amphetamine led to a significant reduction in [11C]-(+)-PHNO BPs in neostriatal ROIs. Largest reductions were found in the VST (Figure 2), a brain region that is particularly sensitive for the actions of psychostimulants and critical for their reinforcing properties (Di Chiara, 1999; Kiyatkin and Brown, 2003; Wise, 2004; Sellings et al, 2006). This result was confirmed in a voxel-wise analysis of parametric BP maps. The specificity of the measured signal-change is supported by significant correlations between changes in [11C]-(+)-PHNO BPs and serum d-amphetamine levels in all ROIs but CAU (Figure 3). Congruent TACs derived in the reference region CER during the d-amphetamine and placebo condition (Figure 1) show that changes in free and non-specific tracer compartments did not contribute significantly to the findings.
This being the first study on D2/3 agonist displacement in humans, the present results will be discussed in light of human experiments using D2/3 antagonist radioligands and data employing agonist radioligands in animals. Several studies have examined the effect of d-amphetamine administration on D2/3 antagonist radioligand binding in humans (Breier et al, 1997; Farde et al, 1992; Laruelle et al, 1995; Abi-Dargham et al, 1998; Cardenas et al, 2004; Riccardi et al, 2006). Owing to methodological differences such as d-amphetamine administration route (most studies used intravenous d-amphetamine administration), tracer administration (eg bolus vs bolus/constant infusion), control conditions (placebo vs no intervention), ROI delineation and differences in scanner equipment and sample composition, results of these studies are not directly comparable to the present ones. However, reductions compared to baseline described after similar doses of d-amphetamine and similar post-intake scanning intervals lie generally between approximately 10 and 15% for the whole striatum (Breier et al, 1997; Cardenas et al, 2004; Farde et al, 1992; Laruelle et al, 1997; Abi-Dargham et al, 1998). Average reductions in [11C]raclopride BPs described in five studies giving separate figures for the VST (Drevets et al, 2001; Leyton et al, 2002; Martinez et al, 2005; Munro et al, 2006; Oswald et al, 2005) are approximately 13% for VST, 6% for CAU, and 11% for PUT. A study performed at our PET Centre using a similar dose of oral d-amphetamine and the same post-intake scanning interval found reductions of 13% in striatal [11C]raclopride BPs (Cardenas et al, 2004). However, this latter study reported considerably higher serum d-amphetamine levels and it used a different PET scanning system. A recent study using a similar dose of oral d-amphetamine and the antagonist D2/3 radioligand [18F]fallypride found reductions of 5.6, 11.2, and 6.6% in CAU, PUT, and VST, respectively (Riccardi et al, 2006), reductions that are sensibly smaller than the 13.2, 20.8, and 24.9% reductions found in these corresponding structures with [11C]-(+)-PHNO. A notable aspect of our results is that [11C]-(+)-PHNO BPs measured in placebo scans were consistently lower than the ones in the test–retest condition. One possible explanation for this result is enhanced competition with endogenous dopamine, released in expectation of a real drug, a finding that has already been described using [11C]raclopride and PET (de la Fuente-Fernandez et al, 2002). However, a direct comparison of [11C]-(+)-PHNO with [11C]raclopride, possibly employing a within-subject design, will help to quantify the difference between agonist and antagonist imaging in competition experiments. All together, it seems that reductions measured with [11C]-(+)-PHNO in our present study are somewhat larger than the ones generally seen with D2/3 antagonist radioligands.
Several animal PET studies have recently reported on the effects of d-amphetamine on in vivo D2/3 agonist radioligand binding in the brain. Experiments have been carried out using [11C]NPA (Narendran et al, 2004), [11C]MNPA (Seneca et al, 2006), and [11C]-(+)-PHNO (Galineau et al, 2006; Ginovart et al, 2006a; Wilson et al, 2005; Narendran et al, 2006). Some of the studies used parallel imaging with the antagonist radioligand [11C]raclopride (Narendran et al, 2004, 2006; Seneca et al, 2006; Ginovart et al, 2006a). These studies show clearly that the d-amphetamine-induced displacement of agonist radioligand binding is one to two thirds larger than that which is measured with [11C]raclopride. A head-to-head comparison of d-amphetamine induced displacement in cats showed an extrapolated maximal reduction of 68% for [11C]raclopride BPs as compared to 96% for [11C]-(+)-PHNO BPs (Ginovart et al, 2006a), and direct comparisons between [11C]NPA and [11C]-(+)-PHNO show that the latter ligand is more sensitive to endogenous dopamine than the former one (Ginovart et al, 2006a). Somewhat in contrast to these findings are results obtained in rats (Wilson et al, 2005) that show maximal reductions of 38% in [11C]-(+)-PHNO binding even with high doses of d-amphetamine. One important difference of this study to the aforementioned experiments is that it was performed in un-anesthetized animals. A preliminary report (McCormick et al, 2006) shows that administration of volatile anaesthetics enhances the effect of d-amphetamine on [11C]-(+)-PHNO binding. In sum, animal evidence shows clearly that [11C]-(+)-PHNO is more vulnerable to competition with endogenous dopamine than [11C]raclopride, and it suggests that, besides higher doses of d-amphetamine administered to animals, anesthesia may in part explain the greater magnitude of [11C]-(+)-PHNO displacement observed in animal studies as compared to the present study.
A peculiarity of [11C]-(+)-PHNO PET is the high BPs measured in GP. Tracer kinetics in GP proved to be different from those in neo-striatal regions, and no significant post-amphetamine reductions were found in the GP. Peak-uptake in GP was lower than in VST, CAU or PUT, and radioligand-washout considerably slower (Figure 1). Since equilibrium is reached later in the GP and since it is more sustained, activity measured in GP throughout the later part of the scanning session contributes substantially more to BP measures derived with SRTM than late-scan activity in neo-striatal ROIs (Ginovart et al, 2006b). As a direct consequence, the more noisy late parts of TACs increase variability of BP measurements in the GP. As shown in Figure 1, peak uptake in VST was slightly lower than in CAU and PUT. Although not to the extent seen in GP, tracer washout from VST seemed to be relatively slow, leading to a more extended equilibrium with greater weight of the late, more ‘noisy’ parts of the TACs in VST as well. This might explain in part the high variability observed in GP and VST, and it may have contributed to the lack of significant post-d-amphetamine reductions in the GP. On the other hand, three individuals with very low d-amphetamine levels showed an increase rather than a decrease in post-d-amphetamine [11C]-(+)-PHNO BPs in GP (Figure 2). As seen easily in Figure 3, this increase contributed substantially to the highly significant correlation between serum d-amphetamine-levels and [11C]-(+)-PHNO BPs in GP. As of yet, it is unclear whether this finding is caused by any real physiological processes in response to low-dose d-amphetamine, or whether this is a spurious finding relating to the high test-retest variability of [11C]-(+)-PHNO PET in the GP. However, a recent study in baboons (Narendran et al, 2006) shows prominent (60%) d-amphetamine induced displacement of [11C]-(+)-PHNO also in GP. It might be worth noting that in the present human study, displacement in GP was significant as well (21.9±9%, t(5)=−5.254, p=0.003) if the three subjects displaying serum d-amphetamine levels more than two SE below the mean were not included into the analysis.
It is unknown up to date what causes the particular binding pattern of [11C]-(+)-PHNO in GP and the relatively high binding to ventral striatal structures. Evidence points towards a greater role of D3 over D2 receptors in [11C]-(+)-PHNO binding in GP, and to a greater role of D3 in VST as compared to CAU and PUT. First, anatomical distribution of [11C]-(+)-PHNO uptake, with high BPs in GP and ventral portions of the neo-striatum, regions where post-mortem studies have shown relatively high densities of D3 receptors (Seeman et al, 2006; Gurevich and Joyce, 1999; Murray et al, 1994), is compatible with a significant contribution of D3 receptors to the captured signal. Second, some studies point towards a higher in vitro (Freedman et al, 1994) and in vivo (Narendran et al, 2006) affinity of [3H]-(+)-PHNO for D3 over D2 receptors. On the other hand, there is evidence suggesting that the affinity of [11C]-(+)-PHNO for D2high is considerably higher than the affinity for D3 receptors (Seeman et al, 2005), and in contrast to pretreatment with the partial D3 receptor agonist BP897 in baboons (Narendran et al, 2006), pre–treatment with the D3 antagonist SB-277011 did not significantly reduce [11C]-(+)-PHNO binding in the cat striatum (Ginovart et al, 2006a). However, reductions in [11C]-(+)-PHNO BPs after d-amphetamine in this study were largest in VST, but smallest in GP. It is thus unlikely that binding to D3 receptors alone accounts for both observations. Other factors, such as regional differences in the amount of dopamine released, should be considered in the interpretation of this finding. In sum, while not conclusive so far, evidence points to an important contribution of D3 receptor binding to the high BPs measured in GP.
Although competitive inhibition of radioligand binding is the hypothesis most commonly put forward to explain reductions in radioligand binding after d-amphetamine administration, several observations suggest that other mechanisms such as receptor internalization (Sun et al, 2003) or changes in receptor affinity (Ginovart et al, 2004) could contribute to this effect (for review see Ginovart, 2005; Laruelle, 2000). It is thus a limitation of the present study—as of all other competition studies performed with D2/3 antagonist radioligands in humans—that the methodology does not allow to identify the exact mechanism leading to decreased radioligand binding after d-amphetamine administration. Another limitation of the present study is that we did not apply correction for partial volume effects (PVEs). Since ROIs such as GP and VST are relatively small for resolutions reached by current PET scanning systems, and since they are adjacent to receptor-rich regions such as PUT, correction for PVEs can be expected to add to the reliability of d-amphetamine-induced BP changes in this regions. However, a recent study using [11C]raclopride showed that correction for PVEs resulted in higher estimates of the d-amphetamine effect in the VST of healthy subjects (Martinez et al, 2005), suggesting that, if anything, we might have underestimated the actual d-amphetamine effects on [11C]-(+)-PHNO BPs in the VST.
A peculiarity of this study was that we observed nausea (in one case vomiting) after injection of [11C]-(+)-PHNO. Nausea and emesis are typical unwanted drug effects during treatment with dopamine agonists. Injected radioligand mass was by no means higher than what is usually administered in PET studies using antagonist radioligands. Still, the close temporal contiguity between tracer injection and nausea is suggestive for a pharmacological effect of [11C]-(+)-PHNO, and, possibly owing to the presence of spare D2high states, some of the pharmacological effects of dopamine agonists, such as inhibition of prolactin secretion (Meller et al, 1991), have been shown to occur at low receptor occupancies. According to the tracer principle, a radioligand should not perturb the biological system it is measuring. Although nausea might be interpreted as an indication that the tracer principle was violated in the present study, evidence suggests that this is unlikely to be the case. First, estimation of central receptor occupancy during [11C]-(+)-PHNO PET according to the method described by Hume et al (1998) results in a mean±SD occupancy of 1.6±0.5% when using an ED50 value of 7.7 nmol/kg as measured in vivo in cats (Ginovart; unpublished observation). This is similar to what has been described for [11C]raclopride PET (Nordström et al, 1992). Second, the ED50 of (+)-PHNO for inducing emesis in animal experiments is two orders of magnitude smaller than the ED50 for inducing motor effects or stereotyped behavior (Martin et al, 1984). Third, therapeutic (+)-PHNO plasma levels, even at the low end, are at least three orders of magnitude higher than what is measured after a single injection of approximately 2 μg total tracer mass as used in our study (Coleman et al, 1990; Ginovart et al, 2006b). Finally, (+)-PHNO-induced emesis is readily prevented by pretreatment with peripherally acting dopamine receptor antagonists in animals (Martin et al, 1984; Nomoto et al, 1987) and humans (Grandas et al, 1987). In sum, nausea is most likely a peripheral effect caused by even minute doses of (+)-PHNO acting at dopamine receptors outside the blood brain barrier in the area postrema (Carpenter, 1990). Pretreatment with a peripheral dopamine receptor antagonist may be a viable strategy to avoid [11C]-(+)-PHNO induced nausea in future studies.
SUMMARY
This study is the first to demonstrate in vivo competition between endogenous dopamine and a D2/3 agonist radioligand in humans. With exception of GP, the study showed clear-cut d-amphetamine effects in all striatal ROIs, and d-amphetamine-induced reductions in [11C]-(+)-PHNO binding were larger than those reported for D2/3 antagonist radioligands in the literature. Our data suggest that, in spite of relatively high test-retest variability, [11C]-(+)-PHNO might be a superior radioligand for investigating alterations in pre-synaptic dopamine release in patients with schizophrenia and other psychiatric disorders.
References
Abi-Dargham A, Gil R, Krystal J, Baldwin RM, Seibyl JP, Bowers M et al (1998). Increased striatal dopamine transmission in schizophrenia: confirmation in a second cohort. Am J Psychiatry 155: 761–767.
Breier A, Su TP, Saunders R, Carson RE, Kolachana BS, de Bartolomeis A et al (1997). Schizophrenia is associated with elevated amphetamine-induced synaptic dopamine concentrations: evidence from a novel positron emission tomography method. Proc Natl Acad Sci USA 94: 2569–2574.
Brown DJ, Luthra SK, Brady F, Prenant C, Dijkstra D, Wikström H et al (1997). Labeling of the D2 agonist -(+)-PHNO using [11C]-propionyl chloride. XIIth International Symposium. Radiopharmaceutical Chemistry, Uppsala, Sweden. Wiley: Chichester, UK. pp 565–566.
Camps M, Cortes R, Gueye B, Probst A, Palacios JM (1989). Dopamine receptors in human brain: autoradiographic distribution of D2 sites. Neuroscience 28: 275–290.
Cardenas L, Houle S, Kapur S, Busto UE (2004). Oral D-amphetamine causes prolonged displacement of [11C]raclopride as measured by PET. Synapse 51: 27–31.
Carpenter DO (1990). Neural mechanisms of emesis. Can J Physiol Pharmacol 68: 230–236.
Coleman RJ, Quinn NP, Traub M, Marsden CD (1990). Nasogastric and intravenous infusions of (+)-4-propyl-9-hydroxynaphthoxazine (PHNO) in Parkinson's disease. J Neurol Neurosurg Psychiatry 53: 102–105.
Cumming P, Gillings NM, Jensen SB, Bjarkam C, Gjedde A (2003). Kinetics of the uptake and distribution of the dopamine D(2,3) agonist (R)-N-n-propylnorapomorphine in brain of healthy and MPTP-treated Gottingen miniature pigs. Nucl Med Biol 30: 547–553.
Cumming P, Wong DF, Dannals RF, Gillings N, Hilton J, Scheffel U et al (2002). The competition between endogenous dopamine and radioligands for specific binding to dopamine receptors. Ann N Y Acad Sci 965: 440–450.
de la Fuente-Fernandez R, Phillips AG, Zamburlini M, Sossi V, Calne DB, Ruth TJ et al (2002). Dopamine release in human ventral striatum and expectation of reward. Behav Brain Res 136: 359–363.
de la Fuente-Fernandez R, Sossi V, Huang Z, Furtado S, Lu JQ, Calne DB et al (2004). Levodopa-induced changes in synaptic dopamine levels increase with progression of Parkinson's disease: implications for dyskinesias. Brain 127: 2747–2754.
Di Chiara G (1999). Drug addiction as dopamine-dependent associative learning disorder. Eur J Pharmacol 375: 13–30.
Drevets WC, Gautier C, Price JC, Kupfer DJ, Kinahan PE, Grace AA et al (2001). Amphetamine-induced dopamine release in human ventral striatum correlates with euphoria. Biol Psychiatry 49: 81–96.
Farde L, Nordstrom AL, Wiesel FA, Pauli S, Halldin C, Sedvall G (1992). Positron emission tomographic analysis of central D1 and D2 dopamine receptor occupancy in patients treated with classical neuroleptics and clozapine. Relation to extrapyramidal side effects. Arch Gen Psychiatry 49: 538–544.
Finnema SJ, Seneca N, Farde L, Shchukin E, Sovago J, Gulyas B et al (2005). A preliminary PET evaluation of the new dopamine D2 receptor agonist [11C]MNPA in cynomolgus monkey. Nucl Med Biol 32: 353–360.
Freedman SB, Patel S, Marwood R, Emms F, Seabrook GR, Knowles MR et al (1994). Expression and pharmacological characterization of the human D3 dopamine receptor. J Pharmacol Exp Ther 268: 417–426.
Galineau L, Wilson AA, Garcia A, Houle S, Kapur S, Ginovart N (2006). In vivo characterization of the pharmacokinetics and pharmacological properties of [11C]-(+)-PHNO in rats using an intracerebral beta-sensitive system. Synapse 60: 172–183.
George SR, Watanabe M, Di Paolo T, Falardeau P, Labrie F, Seeman P (1985). The functional state of the dopamine receptor in the anterior pituitary is in the high affinity form. Endocrinology 117: 690–697.
Ginovart N (2005). Imaging the dopamine system with in vivo [11C]raclopride displacement studies: understanding the true mechanism. Mol Imaging Biol 7: 45–52.
Ginovart N, Galineau L, Willeit M, Mizrahi R, Bloomfield PM, Seeman P et al (2006a). Binding characteristics and sensitivity to endogenous dopamine of [11C]-(+)-PHNO, a new agonist radiotracer for imaging the high-affinity state of D2 receptors in vivo using positron emission tomography. J Neurochem 97: 1089–1103.
Ginovart N, Willeit M, Rusjan PM, Graff A, Bloomfield PM, Houle S et al (2006b). Positron emission tomography quantification of [11C]-(+)-PHNO binding in the human brain. J Cereb Blood Flow Metabo [E-pub ahead of print] doi:10.1038/sj.jcbfm.9600411.
Ginovart N, Wilson AA, Houle S, Kapur S (2004). Amphetamine pretreatment induces a change in both D2-Receptor density and apparent affinity: a [11C]raclopride positron emission tomography study in cats. Biol Psychiatry 55: 1188–1194.
Grandas F, Quinn N, Critchley P, Rohan A, Marsden CD, Stahl SM (1987). Antiparkinsonian activity of a single oral dose of PHNO. Mov Disord 2: 47–51.
Gurevich EV, Joyce JN (1999). Distribution of dopamine D3 receptor expressing neurons in the human forebrain: comparison with D2 receptor expressing neurons. Neuropsychopharmacology 20: 60–80.
Hall H, Halldin C, Dijkstra D, Wikstrom H, Wise LD, Pugsley TA et al (1996). Autoradiographic localisation of D3-dopamine receptors in the human brain using the selective D3-dopamine receptor agonist (+)-PD 128907. Psychopharmacology (Berlin) 128: 240–247.
Hume SP, Gunn RN, Jones T (1998). Pharmacological constraints associated with positron emission tomographic scanning of small laboratory animals. Eur J Nucl Med 25: 173–176.
Hwang DR, Kegeles LS, Laruelle M (2000). N-[(11)C]propyl-norapomorphine: a positron-labeled dopamine agonist for PET imaging of D(2) receptors. Nucl Med Biol 27: 533–539.
Justice AJ, de Wit H (2000). Acute effects of estradiol pretreatment on the response to d-amphetamine in women. Neuroendocrinology 71: 51–59.
Kiyatkin EA, Brown PL (2003). Fluctuations in neural activity during cocaine self-administration: clues provided by brain thermorecording. Neuroscience 116: 525–538.
Koepp MJ, Gunn RN, Lawrence AD, Cunningham VJ, Dagher A, Jones T et al (1998). Evidence for striatal dopamine release during a video game. Nature 393: 266–268.
Lammertsma AA, Bench CJ, Hume SP, Osman S, Gunn K, Brooks DJ et al (1996). Comparison of methods for analysis of clinical [11C]raclopride studies. J Cereb Blood Flow Metab 16: 42–52.
Laruelle M (2000). Imaging synaptic neurotransmission with in vivo binding competition techniques: a critical review. J Cereb Blood Flow Metab 20: 423–451.
Laruelle M, Abi-Dargham A, Gil R, Kegeles L, Innis R (1999). Increased dopamine transmission in schizophrenia: relationship to illness phases. Biol Psychiatry 46: 56–72.
Laruelle M, Abi-Dargham A, van Dyck CH, Gil R, D'Souza CD, Erdos J et al (1996). Single photon emission computerized tomography imaging of amphetamine-induced dopamine release in drug-free schizophrenic subjects. Proc Natl Acad Sci USA 93: 9235–9240.
Laruelle M, Abi-Dargham A, van Dyck CH, Rosenblatt W, Zea-Ponce Y, Zoghbi SS et al (1995). SPECT imaging of striatal dopamine release after amphetamine challenge. J Nucl Med 36: 1182–1190.
Laruelle M, D'Souza CD, Baldwin RM, Abi-Dargham A, Kanes SJ, Fingado CL et al (1997). Imaging D2 receptor occupancy by endogenous dopamine in humans. Neuropsychopharmacology 17: 162–174.
Leff P (1995). The two-state model of receptor activation. Trends Pharmacol Sci 16: 89–97.
Levant B (1998). Differential distribution of D3 dopamine receptors in the brains of several mammalian species. Brain Res 800: 269–274.
Leyton M, Boileau I, Benkelfat C, Diksic M, Baker G, Dagher A (2002). Amphetamine-induced increases in extracellular dopamine, drug wanting, and novelty seeking: a PET/[11C]raclopride study in healthy men. Neuropsychopharmacology 27: 1027–1035.
Martin GE, Williams M, Pettibone DJ, Yarbrough GG, Clineschmidt BV, Jones JH (1984). Pharmacologic profile of a novel potent direct-acting dopamine agonist, (+)-4-propyl-9-hydroxynaphthoxazine [(+)-PHNO]. J Pharmacol Exp Ther 230: 569–576.
Martinez D, Gil R, Slifstein M, Hwang DR, Huang Y, Perez A et al (2005). Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biol Psychiatry 58: 779–786.
Mawlawi O, Martinez D, Slifstein M, Broft A, Chatterjee R, Hwang DR et al (2001). Imaging human mesolimbic dopamine transmission with positron emission tomography: I. Accuracy and precision of D(2) receptor parameter measurements in ventral striatum. J Cereb Blood Flow Metab 21: 1034–1057.
McCormick P, Ginovart N, Vasdev N, Seeman P, Kapur S, Wilson AA (2006). Isoflurane increases both the specific binding ratio and sensitivity to amphetamine challenge of [11C]-(+)-PHNO. Neuroimage 31: T33.
Meller E, Puza T, Miller JC, Friedhoff AJ, Schweitzer JW (1991). Receptor reserve for D2 dopaminergic inhibition of prolactin release in vivo and in vitro. J Pharmacol Exp Ther 257: 668–675.
Mukherjee J, Narayanan TK, Christian BT, Shi B, Dunigan KA, Mantil J (2000). In vitro and in vivo evaluation of the binding of the dopamine D2 receptor agonist (11)C-(R,S)-5-hydroxy-2-(di-n-propylamino)tetralin in rodents and nonhuman primate. Synapse 37: 64–70.
Mukherjee J, Narayanan TK, Christian BT, Shi B, Yang ZY (2004). Binding characteristics of high-affinity dopamine D2/D3 receptor agonists, 11C-PPHT and 11C-ZYY-339 in rodents and imaging in non-human primates by PET. Synapse 54: 83–91.
Munro CA, McCaul ME, Wong DF, Oswald LM, Zhou Y, Brasic J et al (2006). Sex differences in striatal dopamine release in healthy adults. Biol Psychiatry 59: 966–974.
Murray AM, Ryoo HL, Gurevich E, Joyce JN (1994). Localization of dopamine D3 receptors to mesolimbic and D2 receptors to mesostriatal regions of human forebrain. Proc Natl Acad Sci USA 91: 11271–11275.
Narendran R, Hwang DR, Slifstein M, Talbot PS, Erritzoe D, Huang Y et al (2004). In vivo vulnerability to competition by endogenous dopamine: comparison of the D2 receptor agonist radiotracer (−)-N-[11C]propyl-norapomorphine ([11C]NPA) with the D2 receptor antagonist radiotracer [11C]-raclopride. Synapse 52: 188–208.
Narendran R, Slifstein M, Guillin O, Hwang Y, Hwang DR, Scher E et al (2006). Dopamine (D2/3) receptor agonist positron emission tomography radiotracer [(11)C]-(+)-PHNO is a D(3) receptor preferring agonist in vivo. Synapse 60: 485–495.
Nomoto M, Stahl S, Jenner P, Marsden CD (1987). Antiparkinsonian activity of (+)-PHNO in the MPTP-treated common marmoset. Mov Disord 2: 37–45.
Nordström A_L, Farde L, Pauli S, Litton J-E, Halldin C (1992). PET analysis of central [11C]raclopride binding in healthy young adults and schizophrenic patients—reliability and age effects. Hum Psychopharmacol 7: 157–165.
Oswald LM, Wong DF, McCaul M, Zhou Y, Kuwabara H, Choi L et al (2005). Relationships among ventral striatal dopamine release, cortisol secretion, and subjective responses to amphetamine. Neuropsychopharmacology 30: 821–832.
Pruessner JC, Champagne F, Meaney MJ, Dagher A (2004). Dopamine release in response to a psychological stress in humans and its relationship to early life maternal care: a positron emission tomography study using [11C]raclopride. J Neurosci 24: 2825–2831.
Riccardi P, Li R, Ansari MS, Zald D, Park S, Dawant B et al (2006). Amphetamine-induced displacement of [18F] fallypride in striatum and extrastriatal regions in humans. Neuropsychopharmacology 31: 1016–1026.
Rusjan P, Mamo D, Ginovart N, Hussey D, Vitcu I, Yasuno F et al (2006a). An automated method for the extraction of regional data from PET images. Psychiatry Res 147: 79–89.
Rusjan PM, Mizrahi R, Ginovart N, Graff A, Willeit M, Vitcu I et al (2006b). Validation of a method for automatic quantification of radioactivity in the globus pallidus in [11C]-(+)-PHNO PET images. Neuroimage 31: T96.
Seeman P, Ko F, Willeit M, McCormick P, Ginovart N (2005). Antiparkinson concentrations of pramipexole and PHNO occupy dopamine D2(high) and D3(high) receptors. Synapse 58: 122–128.
Seeman P, Wilson A, Gmeiner P, Kapur S (2006). Dopamine D2 and D3 receptors in human putamen, caudate nucleus, and globus pallidus. Synapse 60: 205–211.
Sellings LH, McQuade LE, Clarke PB (2006). Characterization of dopamine-dependent rewarding and locomotor stimulant effects of intravenously-administered methylphenidate in rats. Neuroscience 14: 1457–1468.
Seneca N, Finnema SJ, Farde L, Gulyas B, Wikstrom HV, Halldin C et al (2006). Effect of amphetamine on dopamine D2 receptor binding in nonhuman primate brain: a comparison of the agonist radioligand [11C]MNPA and antagonist [11C]raclopride. Synapse 59: 260–269.
Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E et al (1998). The Mini-International Neuropsychiatric Interview (MINI): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry 59 (Suppl 20): 22–33; quiz 34–57.
Shi B, Narayanan TK, Christian BT, Chattopadhyay S, Mukherjee J (2004). Synthesis and biological evaluation of the binding of dopamine D2/D3 receptor agonist, (R,S)-5-hydroxy-2-(N-propyl-N-(5′-(18)F-fluoropentyl)aminotetralin ((18)F-5-OH-FPPAT) in rodents and nonhuman primates. Nucl Med Biol 31: 303–311.
Shi B, Narayanan TK, Yang ZY, Christian BT, Mukherjee J (1999). Radiosynthesis and in vitro evaluation of 2-(N-alkyl-N-1′-11C-propyl)amino-5-hydroxytetralin analogs as high affinity agonists for dopamine D-2 receptors. Nucl Med Biol 26: 725–735.
Sibley DR, De Lean A, Creese I (1982). Anterior pituitary dopamine receptors. Demonstration of interconvertible high and low affinity states of the D-2 dopamine receptor. J Biol Chem 257: 6351–6361.
Spinks R, Nopoulos P, Ward J, Fuller R, Magnotta VA, Andreasen NC (2005). Globus pallidus volume is related to symptom severity in neuroleptic naive patients with schizophrenia. Schizophr Res 73: 229–233.
Sun W, Ginovart N, Ko F, Seeman P, Kapur S (2003). In vivo evidence for dopamine-mediated internalization of D2-receptors after amphetamine: differential findings with raclopride versus spiperone. Mol Pharmacol 63: 456–462.
Tedroff J, Pedersen M, Aquilonius SM, Hartvig P, Jacobsson G, Langstrom B (1996). Levodopa-induced changes in synaptic dopamine in patients with Parkinson's disease as measured by [11C]raclopride displacement and PET. Neurology 46: 1430–1436.
Volkow ND, Wang GJ, Fowler JS, Logan J, Schlyer D, Hitzemann R et al (1994). Imaging endogenous dopamine competition with [11C]raclopride in the human brain. Synapse 16: 255–262.
Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Childress AR et al (2006). Cocaine cues and dopamine in dorsal striatum: mechanism of craving in cocaine addiction. J Neurosci 26: 6583–6588.
White TL, Justice AJ, de Wit H (2002). Differential subjective effects of D-amphetamine by gender, hormone levels and menstrual cycle phase. Pharmacol Biochem Behav 73: 729–741.
Willeit M, Ginovart N, Kapur S, Houle S, Hussey D, Seeman P et al (2006). High-affinity states of human brain dopamine D2/3 receptors imaged by the agonist [11C]-(+)-PHNO. Biol Psychiatry 59: 389–394.
Wilson AA, McCormick P, Kapur S, Willeit M, Garcia A, Hussey D et al (2005). Radiosynthesis and evaluation of [11C]-(+)-4-propyl-3,4,4a,5,6,10b-hexahydro-2H-naphthooxazin-9 -ol as a potential radiotracer for in vivo imaging of the dopamine D2 high-affinity state with positron emission tomography. J Med Chem 48: 4153–4160.
Wise RA (2004). Dopamine, learning and motivation. Nat Rev Neurosci 5: 483–494.
Zahniser NR, Molinoff PB (1978). Effect of guanine nucleotides on striatal dopamine receptors. Nature 275: 453–455.
Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS et al (2004). Dopamine transmission in the human striatum during monetary reward tasks. J Neurosci 24: 4105–4112.
Zijlstra S, van der Worp H, Wiegman T, Visser GM, Korf J, Vaalburg W (1993a). Synthesis and in vivo distribution in the rat of a dopamine agonist: N-([11C]methyl)norapomorphine. Nucl Med Biol 20: 7–12.
Zijlstra S, Visser GM, Korf J, Vaalburg W (1993b). Synthesis and in vivo distribution in the rat of several fluorine-18 labeled N-fluoroalkylaporphines. Appl Radiat Isot 44: 651–658.
Acknowledgements
This work was supported in part by the Canadian Institutes for Health Research (Grant #74702 to AAW). Funding of the PET camera system CPS-HRRT was supported by the Canada Foundation for Innovation, the Ontario Innovation Trust and the Ontario Research and Development Challenge Fund. SK was supported by a CRC Chair in Schizophrenia and Therapeutic Neuroscience. We thank Penny Barsoum, Anna Carella, Armando Garcia, Doug Hussey, Alvina Ng, Nicole Praschak-Rieder, CM Shammi, and Winston Stableford for their indispensable technical and logistic assistance, and Peter Bloomfield for physics support. We also thank Laurie Zawertailo and Usoa Busto for their intellectual input during planning of this study. This work was presented in part at the meeting of the Society of Biological Psychiatry, 2006, Toronto, ON.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Willeit, M., Ginovart, N., Graff, A. et al. First Human Evidence of d-Amphetamine Induced Displacement of a D2/3 Agonist Radioligand: A [11C]-(+)-PHNO Positron Emission Tomography Study. Neuropsychopharmacol 33, 279–289 (2008). https://doi.org/10.1038/sj.npp.1301400
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1301400
Keywords
This article is cited by
-
Effect of scan-time shortening on the 11C-PHNO binding potential to dopamine D3 receptor in humans and test–retest reliability
Annals of Nuclear Medicine (2023)
-
The relationship between glutamate, dopamine, and cortical gray matter: A simultaneous PET-MR study
Molecular Psychiatry (2022)
-
EANM guideline for harmonisation on molar activity or specific activity of radiopharmaceuticals: impact on safety and imaging quality
EJNMMI Radiopharmacy and Chemistry (2021)
-
The influence of conditioned stimuli on [11C]-(+)-PHNO PET binding in tobacco smokers after a one week abstinence
Scientific Reports (2021)
-
D3 dopamine receptors and a missense mutation of fatty acid amide hydrolase linked in mouse and men: implication for addiction
Neuropsychopharmacology (2020)
