
Abstract
MMP-11 is a bad prognosis paracrine factor in invasive breast cancers. However, its mammary physiological function remains largely unknown. In the present study we have investigated MMP-11 function during postnatal mammary gland development and function using MMP-11-deficient (MMP-11−/−) mice. Histological and immunohistochemical analyses as well as whole-mount mammary gland staining show alteration of the mammary gland in the absence of MMP-11, where ductal tree, alveolar structures and milk production are reduced. Moreover, a series of transplantation experiments allowed us to demonstrate that MMP-11 exerts an essential local paracrine function that favors mammary gland branching and epithelial cell outgrowth and invasion through adjacent connective tissues. Indeed, MMP-11−/− cleared fat pads are not permissive for wild-type epithelium development, whereas MMP-11−/− epithelium transplants grow normally when implanted in wild-type cleared fat pads. In addition, using primary mammary epithelial organoids, we show in vitro that this MMP-11 pro-branching effect is not direct, suggesting that MMP-11 acts via production/release of stroma-associated soluble factor(s). Finally, the lack of MMP-11 leads to decreased periductal collagen content, suggesting that MMP-11 has a role in collagen homeostasis. Thus, local stromal MMP-11 might also regulate mammary epithelial cell behavior mechanically by promoting extracellular matrix stiffness. Collectively, the present data indicate that MMP-11 is a paracrine factor involved during postnatal mammary gland morphogenesis, and support the concept that the stroma strongly impact epithelial cell behavior. Interestingly, stromal MMP-11 has previously been reported to favor malignant epithelial cell survival and promote cancer aggressiveness. Thus, MMP-11 has a paracrine function during mammary gland development that might be harnessed to promote tumor progression, exposing a new link between development and malignancy.
Similar content being viewed by others

Introduction
Matrix metalloproteinases (MMPs) mediate a number of physiological and pathological processes such as matrix degradation, tissue remodeling, inflammation and tumor progression.1, 2 MMP-11, a secreted MMP previously called stromelysin-3, was first identified in breast carcinomas. MMP-11 is the paradigm of tumor proteinases not expressed in the malignant epithelial cells but in the non-malignant adjacent mesenchymal/stromal cells.3, 4 High MMP-11 levels are associated with poor patient outcome in various cancers,5, 6, 7, 8, 9 a large depth of invasion and metastases.10, 11 Preclinical studies in mice show that MMP-11 promotes tumors by favoring cancer cell survival in a tissue environment initially not permissive.12, 13
Moreover, MMP-11 is expressed and secreted by adipocytes and has a negative regulatory function during adipogenesis.4, 6, 14 Interestingly, in human tumors, invasive cancer cells induce MMP-11 expression by proximal adipocytes, which in turn leads to the accumulation of peritumor fibroblast-like cells, known to favor cancer cell survival and tumor progression.6, 14 However, how adipocyte-related MMP-11 acts at the physiological level remains largely unknown. As stromal cells are potent mediators in the proper development and maintenance of mammary ductal morphogenesis,15, 16, 17, 18, 19, 20 we hypothetized that MMP-11 might participate in this biological process.
Although the rudimental mammary ductal tree is formed prior to birth, the main mammary gland development occurs postnatally, first during puberty through ductal structure growth including terminal end bud (TEB) appearance, ductal elongation and branching, invasion and filling of the fat pad, and second during pregnancy through growth and differentiation of functional mature alveoli. Reciprocal interactions between the epithelium and the connective tissues dictate the terminal branching pattern of the ducts and contribute to mammary gland morphogenesis, although the details of these events are not fully understood.14, 21, 22, 23, 24, 25
In the present study, we investigated the impact of MMP-11 on mammary gland postnatal development using wild-type and MMP-11 knockout (MMP-11−/−) mice.26 We studied the relative impact of MMP-11 on epithelial and stromal cell compartments. We show that stromal MMP-11 is required for optimal ductal morphogenesis and function. Furthermore, we show in vitro that this pro-branching MMP-11 function is not a direct effect. Finally, we show that MMP-11 positively regulates periductal collagen homeostasis. This study provides the first evidence that MMP-11 has a paracrine role in postnatal mammary gland development.
Results
MMP-11 is required for optimal postnatal mammary gland morphogenesis but does not influence the intrinsic organization of the mammary epithelium
The MMP-11 status of mice was established by PCR genotyping (Supplementary Figure S1A). As expected, in virgin wild-type mammary gland, MMP-11 was expressed at a low level (RT–PCR; Supplementary Figure S1B). Using in situ hybridization, we observed that whereas no MMP-11 was detected in the mammary epithelium, MMP-11 was present in the stroma. Intense punctual staining was restricted to periductal adipose tissue in accordance with previous mouse4, 5, 27, 28 and human3, 4 studies (Supplementary Figure S1C, right panel). Embryonic limb buds were used as a positive control (Supplementary Figure S1C, left panel). Moreover, there were no obvious differences between wild-type and MMP-11−/− mammary gland weight and volume at 4, 6 and 12 weeks of age (Supplementary Figures S1D and E, respectively).
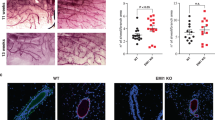
To investigate whether loss of MMP-11 affects postnatal development, we examined the inguinal #4 mammary gland phenotype of 3-, 4-, 6- and 12-week-old littermate mice by whole-mount carmine staining (Figure 1A). The initial rudimental ductal trees appeared normal in MMP-11−/− glands in 3-week-old pre-pubertal mice. During puberty, TEBs appear and bifurcate repeatedly to form the ductal tree to develop into a mature gland. Disruption of TEBs is often associated with delayed ductal outgrowth and impaired branching morphogenesis.29, 30 Accordingly, at 4 weeks of age, many distal branches of the wild-type ducts were tipped with TEBs (32.6±8.35 per gland), indicating that active ductal morphogenesis was in progress. However, in the MMP-11−/− glands, although morphogenesis process was also initiated, there were less TEBs (22.0±5.1 per gland; P=0.04; Figure 1B). Nevertheless, carmine-stained MMP-11−/− TEBs exhibit a normal structure larger than ducts, with staining densification at their apical part as already reported31 (Figure 1Ab). This MMP-11−/− phenotype was more pronounced at 6 weeks of age, and the number of TEBs was reduced compared with wild-type (13.6±6.4 vs 39.6±14.3; P=0.006; Figure 1B). Additionally, mammary gland complexity, which represents the extent of ductal branching via TEB dichotomy and/or duct side branching, was quantified by determining the number of ductal branches intersecting a line arbitrarily drawn ahead of the lymph node and perpendicularly to the long axis of the mammary gland, as previously described.32 At 6 weeks of age, mammary gland complexity was 13.6±1.1 for wild-type mice and 4.8±2.0 for MMP-11−/− animals (P<0.001; Figure 1C). Similarly, at 12 weeks of age, the ductal complexity was lower in MMP-11−/− than in wild-type mice (9.8±1.1 vs 15.0±3.3; P<0.006; Figure 1C). It has been reported that, at the end of pubertal development, the ductal tree almost reaches the border of fat pad and that only very few TEBs remain present.25 This phenotype was observed at 12 weeks of age in the wild-type mammary glands, but the MMP-11−/− ductal structures did not fill the entire fat pad, and more TEBs were always present (29.0±12.1 vs 14.2±5.3; P<0.05) (Figures 1A and B). Finally, older MMP-11−/− mice (12–30-week-old) did not show more expanded ductal tree (not shown), indicating that mammary gland development is not delayed but defective.

Mammary ductal morphogenesis is impaired in MMP-11−/− mice, although TEB and ductal epithelial architecture is not obviously altered. (A) Whole-mount carmine staining of wild-type (upper panels) and MMP-11−/− (lower panels) mammary glands at 3, 4, 6 and 12 weeks of age; LN: lymph node; magnification: (a) × 5; (b) × 25. (B) TEB quantification at 4, 6 and 12 weeks of age. (C) Quantification of ductal complexity represented by the number of branches intersecting a line (blue) drawn ahead of the lymph node and perpendicular to the long axis of the mammary gland at 6 and 12 weeks of age. The ductal trees are less developed. Moreover, the gland does not reach the distal limits of the fat pad, indicating that the ductal elongation and invasive potential are weaker. Purple dotted lines delineate the fat pad limits. Five pairs of littermates were used for each condition. (D) HE staining of 6-week-old MMP-11−/− epithelial duct showing TEB at the distal tip. MMP-11−/− TEB exhibits a normal structure associating an outer layer of cap cells (cc) to several layers of body cells (bc). Magnification: × 100 and × 400. (E) Mammary gland sections from 6-week-old wild-type (upper panels) and MMP-11−/− (lower panels) littermates were analyzed by immunofluorescence staining for α-smooth muscle actin (α-SMA) and E-cadherin as indicated. E-cadherin is a marker for luminal epithelial cells, whereas α-SMA is expressed by myoepithelial cells. Both wild-type and MMP-11−/− mammary TEBs and ducts are lined by inner luminal epithelial cells (red) and an outer layer of myoepithelial cells (green). Cell nuclei are colored in blue using DAPI; magnification: × 400.
TEBs comprise a dynamic mass of luminal body cells surrounded by a motile cap cell layer.25, 31 The ducts are hollow, lined with a layer of luminal epithelial cells and then a layer of myoepithelial cells.2531 To determine whether there are cellular organizational defects in MMP-11−/− epithelial TEBs and ducts, we performed hematoxilin–eosin (HE) staining. Histological analysis showed that TEBs and ducts exhibited normal structure in the absence of MMP-11 (Figures 1D and 3a). Accordingly, immunofluorescence analysis showed E-cadherin, a marker of the cuboidal luminal epithelial cells, at the cell–cell contacts in the TEBs and ducts and α-smooth muscle actin (α-SMA), a marker of myoepithelial cells, in the myoepithelial cell layer lining the TEBs and ducts regardless of MMP-11 status (Figure 1E).
Thus, pubertal duct morphogenesis is impaired in MMP-11−/− mammary glands, suggesting that MMP-11 functions in a non-redundant manner to allow the epithelium to fully expand and spread throughout the stromal microenvironment. However, the absence of MMP-11 does not affect epithelial cellular architecture, suggesting that the MMP-11−/− phenotype is not due to an intrinsic defect of the epithelial cell compartment.
MMP-11 is required for optimal postnatal mammary gland function
The mammary alveolus structures develop gradually during pregnancy and reach their mature forms by parturition. To determine whether MMP-11 affects the functional differentiation of the mammary gland, wild-type or MMP-11−/− mice were mated for the first time at 12 weeks of age. The ductal/alveolar structures were examined during early pregnancy (day 10 after conception), late pregnancy (day 18 after conception) and lactation (day 3 post partum) by whole-mount carmine staining (Figure 2a; data not shown). The alveoli developed in both genotypes, but their density was reduced in the absence of MMP-11. HE histological analysis (Figure 2b) showed that the lactating wild-type mammary gland was composed almost entirely of alveoli greatly distended with milk. The interlobular connective tissue was reduced. By contrast, MMP-11−/− terminal alveolus number was reduced and their structure defective. They were not filled by milk and the milk-secreting vacuoles exhibited various sizes, whereas they were homogeneous in wild-type mice.

Mammary alveolar morphogenesis and differentiation are impaired in MMP-11−/− mice. (a) Whole-mount carmine staining of wild-type (upper panel) and MMP-11−/− (lower panel) mammary gland at pregnancy (10th day) and lactation (3rd day); magnification: × 5 and × 25; purple dotted lines delineate the fat pad limits. Five pairs of littermates were used for each condition. (b) HE staining of lactating mammary gland sections: MMP-11−/− lactating glands are hypotrophic and greatly disorganized compared with wild-type gland. Three images from three MMP-11−/− mice are shown. Magnification: × 200. (c) Body weight of pups fed by wild-type or MMP-11−/− females from 1 day to 28 days postnatally. Pups fed by MMP-11−/− females show retarded growth until post-weaning (∼20 days after parturition) when they rapidly catch up with wild-type pups. Three litters (18 pups) per genotype were used for this analysis. Data are expressed as mean±s.d.
Mammary gland function was assessed by measuring, from 1–28 days after parturition, the body weight of pups fed by wild-type or MMP-11−/− first-time littermate mothers (six pups/female). All pups were alive at the end of the test. However, from days 5–18, the pups fed by the MMP-11−/− mothers had lower body weights compared with those fed by the wild-type mothers. However, the body weight of the two sets of pups rapidly caught up after weaning (around day 20) (Figure 2c). These results indicated that nursing was altered in the absence of MMP-11. This probably does not result from impaired milk quality, but rather from decreased milk production due to altered alveoli.
Thus, the absence of MMP-11 leads to alveolus hypotrophy and reduced mammary gland functionality.
MMP-11 has a role in the mammary gland stromal compartment
The mammary stroma is mainly composed of adipocytes and collagen-rich extracellular matrix that are essential for the regulation of mammary gland development.15, 16, 17 HE staining showed that the size of the adipocytes appeared larger (Figure 3a), consistent with previous studies showing adipocyte hypertrophy in the MMP-11−/− abdominal fat, and that MMP-11 is a negative regulator of adipogenesis.4, 33

The MMP-11−/− mammary stroma is dramatically altered. (a) HE staining of 6-week-old wild-type and MMP-11−/− mammary gland sections as indicated. The bilayered mammary ducts (left panels) showing an inner layer of luminal cells and an outer layer of myoepithelial cells are similar irrespective of the MMP-11 status. A low number of fibroblasts (f) are similarly distributed around the ducts. There are very few immune cells in both genotypes. Adipose tissue (right panel) is mainly composed of adipocytes. The size of the adipocytes appears larger in the MMP-11−/− gland; magnification: × 400. (b) Masson’s trichrome staining of mammary gland sections shows that collagen (blue) is more abundant in wild-type than in MMP-11−/− periductal stroma (red arrows); the stroma located around the TEBs is less abundant and has a similar content regardless of MMP-11 status (green arrows); magnification: × 200. (c) Sirius red staining of collagen (red) confirms the Masson’s results concerning ducts (red arrows) and TEBs (green arrows); magnification: × 400.
Collagen content was estimated using Masson’s trichrome and sirius red staining. In both genotypes, collagen was around the ductal and TEB structures (Figures 3b and c). However, in the MMP-11−/− glands, the width of periductal collagen was reduced compared with the wild type (red arrows). As previously reported,22 the peri-TEB collagen depots were less dense compared with periductal depots. No clear differences were observed between wild type and MMP-11−/− peri-TEB collagen levels (green arrows).
Moreover, HE analysis showed similar fibroblast pattern irrespective of the MMP-11 status. They were mainly present in the periductal support tissue and represented a small population (Figure 3a). Accordingly, mammary gland QRT–PCR analysis did not allow the detection of the expression of vimentin, a fibroblast marker (positive control: 3T3L1 fibroblasts). Finally, careful histological analysis showed similar low number of immune cells in both wild-type and MMP-11−/− mammary glands (Figure 3a; data not shown). Accordingly, CD11b, an immune cell marker, was expressed at the same low level using QRT–PCR (wild type: 1.17±0.9; MMP-11−/−: 1.21±0.19; NS; positive control: conventional spleen dentritic cells). Thus, the absence of MMP-11 does not obviously have an impact on fibroblasts or immune cells.
Collectively, these data show that the two main components of the mammary stroma are greatly altered by the absence of MMP-11, suggesting that these alterations might be responsible for impaired postnatal mammary morphogenesis and function.
MMP-11 mammary stroma promotes in vivo ductal morphogenesis
To investigate the respective functional potential of MMP-11−/− mammary epithelia and stroma in forming the mature ductal structure, epithelium from 3-week-old pre-pubertal MMP-11−/− mice was transplanted into cleared fat pads of littermate wild-type mice and vice versa, as previously described34, 35 (Supplementary Figures S2A and B). Transplants were analyzed at 3, 6 and 9 weeks after transplantation for ductal outgrowth. Three and 6 weeks post transplantation, the MMP-11−/− epithelium formed number of TEBs and ducts in the wild-type fat pads (Figure 4a). Interestingly, at these times, the wild-type mammary epithelium was less developed in the MMP-11−/− fat pads and showed a lower number of ducts that were also shorter in length (Figure 4b). These differences were amplified at 9 weeks post transplantation. Thus, during puberty, the MMP-11−/− transplants formed a normal number of ducts that invaded the entire wild-type fat pad. By contrast, the wild-type transplants formed only sparse ducts that did not invade the entire fat pad when grown in a MMP-11−/− environment. As expected, ductal morphogenesis in the MMP-11−/− control glands (controlateral non-operated #4 glands) were impaired at all ages studied compared with the wild-type control (controlateral non-operated #4 glands) (Supplementary Figure S2C).

MMP-11−/− epithelial transplants develop normally into wild-type cleared fat pad. (a, b) Whole-mount carmine staining of transplanted 3-week-old mammary glands at 3, 6 and 9 weeks post-operation (p.o.). (a) MMP-11−/− epithelia (MMP-11−/− E) undergo proper ductal outgrowth within the wild-type cleared fat pad stroma (WT S). (b) By contrast, wild-type epithelia (WT E) generate less ducts when transplanted into MMP-11−/− cleared fat pads (MMP-11−/− S). Moreover, the gland does not reach the fat pad border, indicating that the ductal elongation and invasive potential are also weaker; magnification: × 5; LN: lymph node.
These results confirm that the MMP-11−/− epithelia have no intrinsic functional deficiencies and support the concept that stromal MMP-11 exerts pro-branching and pro-invasive functions either directly or indirectly.
MMP-11 pro-branching function is not a direct effect
To address this question, we investigated MMP-11 pro-branching function, using a three-dimensional culture system to allow the accurate in vitro modeling of the branching process.36, 37 We isolated primary mammary epithelial organoids from the mammary glands of 14-week-old virgin wild-type mice. The organoids were cultured in matrigel three-dimensional gels. Organoids were characterized using immunofluorescence analysis (Figure 5a). No staining was obtained using antibodies directed against vimentin and CD31, which are markers of stromal cells and endothelial cells as shown using 3T3L1 and BEND cells, respectively (Figure 5b). These results indicated that organoids did not contain these two cell types. By contrast, antibodies directed against keratins K8/18 and K14, normally expressed by epithelial cells, gave positive staining indicating that organoids are of epithelial nature (Figures 5a and c).

Mammary organoids are composed of epithelial cells. (a) Confocal analysis of immunofluorescence staining: no staining was obtained using antibodies directed against CD31 and vimentin, indicating that organoids do not contain endothelial cells or fibroblasts. However, antibodies against keratin K8/18 and K14 gave positive staining, establishing the epithelial nature of organoids. Moreover, K14 localizes more at the periphery of organoids, whereas K8/18 is more homogenous. (b) Positive immunofluorescence controls were 3T3L1 mouse fibroblasts for vimentin (ATCC). BEND mouse endothelial cells for CD31 (generous gift from Kari Alitalo, Haartman Institute, Helsinki, Finland). (c) Immunofluorescence experiment on one branched organoid (TGFα-treated; day 6) shows a similar spatial arrangement concerning K14 myoepithelial cells. Cell nuclei are colored in blue using DAPI.
The mammary organoids were first treated for 4 days with culture media containing buffer alone (negative control), buffer with TGFα (positive control) or buffer with active or inactive forms of mouse MMP-11 recombinant protein produced in the laboratory38 (rMMP-11a and rMMP-11i, respectively; Supplementary Figure S3). Two MMP-11 concentrations were used (1 μg/ml and 10 μg/ml) and organoids were analyzed for branching morphogenesis. The medium was changed every day because MMP-11 is susceptible to autolysis. Non-treated organoids exhibited restricted branching (Figure 6a; negative controls). As expected, TGFα allowed multicellular epithelial bud elongation out from the organoids, as previously reported37 (Figure 6b). Compared with controls, the active rMMP-11a did not induce mammary organoid branching (Figures 6c and e). Treatment with inactive rMMP-11i had no effect (Figures 6d and f). Finally, similar results were obtained using recombinant MMP-11 at 1 μg/ml or 10 μg/ml for a longer time (7 days; Figures 6c–f). These results indicated that MMP-11 does not directly promote ductal branching morphogenesis.

Organoid response to active rMMP-11a recombinant protein at various concentrations and times. Evolution of organoids in Matrigel, after various treatments for 4 and 7 days. (a) Untreated organoids (buffer alone) fail to undergo morphological changes, except lumen expansion (negative control). (b) In contrast, TGFα stimulation initiates bud formation (positive control). (c, e) Morphology of organoids treated with buffer containing active recombinant MMP-11 (rMMP-11a) was not modified irrespective of the concentration used, indicating that MMP-11 has no significant direct effect on organoid branching. (d, f) Inactive recombinant MMP-11 (rMMP-11i) had no effect; original magnification: × 10.
Thus, MMP-11 promotes ductal branching morphogenesis in an indirect manner presumably by inducing production/release of stroma-associated soluble factor(s) necessary to mediate its function.
Discussion
In the present study, we demonstrate in vitro and in vivo that stromal MMP-11 is required for proper postnatal mammary gland morphogenesis and function. Indeed, the absence of MMP-11 leads to reduced ductal trees and alveolar density, altered alveolus organization and ultimately lower milk production.
The epithelial architecture of TEBs and ducts has an essential function in the overall mammary gland development. Thus, TEB disruption is often associated with delayed ductal outgrowth and impaired branching.29, 30 We show that the absence of MMP-11 does not intrinsically affect the cellular organization of the mammary epithelium compartment. Initial pre-pubertal rudimental ductal trees were similar regardless of the MMP-11 status. This was further confirmed at the functional level as the MMP-11−/− pre-pubertal mammary epithelium develops normally when implanted into wild-type cleared host mammary fat pads. These data suggest that MMP-11 is involved in epithelium morphogenesis and growth in a paracrine manner. It has been shown that epithelial–stromal interactions are essential for TEB and duct morphogenesis and the functional differentiation of the mammary gland.19, 22, 25, 39, 40, 41, 42, 43 Strongly supporting our hypothesis, we observed that wild-type pre-pubertal epithelia transplanted into MMP-11−/− cleared fat pads do not fully mature and show reduced ductal outgrowth and morphogenesis. Thus, local stromal MMP-11 is important for correct postnatal mammary gland development.
To further study the impact of MMP-11, we used a surrogate in vitro three-dimensional culture model using primary epithelial mammary organoids to investigate the branching process, as previously described.36, 37, 40 Treatment with active rMMP-11a recombinant protein does not allow organoid branching, showing that MMP-11 acts in an indirect manner, presumably via stromal soluble factor(s) whose number and nature remain to be studied. In this context, mammary adipocytes have been shown to be essential for the regulation of mammary gland development.15, 16, 17 Interestingly, we have previously shown that MMP-11 expressed by adipocytes exerts a negative effect on adipogenesis,4, 6, 14, 33 suggesting that adipocyte-related factor(s) might be involved in the MMP-11 function during mammary gland development. In this context, we observed in vitro that wild-type adipocyte culture medium favors the occurrence of organoid protrusions (Supplementary Figure S4). Moreover, this protrusion inducability was significantly reduced for the MMP-11−/− adipocyte culture medium. Interestingly, treatment of MMP-11−/− adipocytes with active rMMP-11a recombinant protein rescues their altered function for inducing mammary organoid protrusions. This protrusion process shows discrepancies with the previously reported organoid branching36, 37 and remains to be studied in detail.
Another interesting result is that MMP-11 influences fat pad collagen homeostasis. The mammary extracellular matrix normally contains high amounts of collagen.15, 16, 17, 22, 39, 44, 45. We show that width of the periductal collagen layer is lower in MMP-11−/− compared with wild-type glands. Thus, MMP-11 is associated with the presence of more collagen. To date, only collagen VI, an adipocyte-related extracellular matrix component, has been identified as a MMP-11 substrate. MMP-11 is required for correct collagen VI folding and therefore for fat tissue cohesion and adipocyte function.46 Indeed, specific proteolysis is sometimes required for correct collagen formation/folding 47.48 Irrespective of the mechanism, by modulating pericellular collagen composition, MMP-11 regulates matrix stiffness and can thereby influence epithelial cell behavior, as previously reported.34, 49
Several other MMPs are produced by mammary glands by fibroblasts only (that is, MMP-2 and MMP-3), epithelial and stromal cells (that is, MMP-14), epithelial cells only (that is, MMP-7) or predominantly by local immune cells (that is MMP-9).22, 25 However, to date, only a few have been investigated for their participation in postnatal mammary gland development. How they work at the cellular and/or molecular levels is not well defined.22, 50, 51 While MMP-9 has no effect, MMP-2−/− mice, like MMP-14-overexpressing mice, show excessive branching, whereas MMP-3−/− mice show defective branching. Thus, MMPs might exert inverse ductal branching functions. In this context, MMP-11 promotes ductal branching. Finally, MMP-11 function is not rescued by the compensatory activity of other MMPs in MMP-11−/− mice, indicating that MMP-11 has a non-redundant role in postnatal mammary gland development.
Both TEB dichotomy and ductal side branching globally result in an increased epithelial compartment and require invasion of the epithelium through the surrounding connective fat pad separated by sparse and mature basement membranes, respectively. These processes are accompanied by intense stroma remodeling. Similar epithelial expansion and invasion, and stromal remodeling are characteristics of malignant tumors.52 Interestingly, stroma-mediated MMP-11 function favors breast cancer agressiveness.4, 6 In this context, the present data strongly implicate MMP-11 as one factor that normally has a role in mammary gland development and is harnessed to promote tumor agressiveness.
In conclusion, our data show that MMP-11 has a role in postnatal mammary gland morphogenesis and differentiation. In a paracrine manner, MMP-11 might regulate epithelial cell behavior both biochemically via inducing molecular processes and/or mechanically by modulating the degree of extracellular matrix flexibility. Deciphering the cellular and molecular mechanisms behind the stromal–epithelial cell heterotypic crosstalk/interaction is therefore of great interest because it might provide new targets for improving cancer diagnosis/prognosis and for the design of innovative therapeutic strategies.
Materials and methods
Animals
We have previously generated MMP-11−/− mice with 129/Svj D3 genetic background.26 Protocols were approved by the Animal Welfare and Research Committee/AWRC at IGBMC institute, Strasbourg, France. All experiments were performed using inguinal #4 mammary glands of littermate MMP-11−/− or wild-type females obtained by heterozygous mouse crossing. Genomic DNA extracted from mouse tails (∼1–2 weeks of age) was analyzed by PCR amplification using sense primer A: 5′-TTCTAACATCCCTCTGGGCTC-3′ (exon 6) or primer B: 5′-GCCGCTTTTCTGGATTCATCG-3′ (neomycin gene) and antisense primer C: 5′-GTGGAAACGCCAATAGTCTC-3′ (exon 7) (wild-type (primers A+C); MMP-11−/− (primers B+C)).
Total RNA RT–PCR and QRT–PCR analysis
Reverse transcription was carried out with a superScrip system (Invitrogen, Carlsbad, CA, USA) on total RNA extracted from mammary gland (TRI reagent, Invitrogen). For MMP-11 PCR, DNA primer sequences were sense primer D: 5′-CCGAAGGGGCATCCAGCACC-3′ and antisense primer E: 5′-GCATCCACAGGGCTGGGCAG-3′. GAPDH was used as control: sense primer F: 5′-ACTGGCATGGCCTTCCGTGTTC-3′ and reverse primer G: 5′-TCTTGCTCAGTGTCCTTGCTGG-3′.
QRT–PCR analysis was performed on mammary gland total RNA using the following primers:
Vimentin: 5′-TGCGCCAGCAGTATGAAA-3′; 5′-GCCTCAGAGAGGTCAGCAAA-3′.
CD11b: 5′-CGTCCCCAGTGTGAGTGTC-3′; 5′-TTTCCCAATGTAGCCAGACTC-3′
Positive control: HPRT: 5′-TCCTCCTCAGACCGCTTTT-3′; 5′-CCTGGTTCATCATCGCTAATC-3′.
In situ hybridization analysis
Tissue sections (15-μm thick) were hybridized using digoxigenin (DIG)-labeled sense or antisense mouse MMP-11 cDNA (nucleotides 179–1505; DIG RNA Labeling Kit; Boehringer Mannheim Corp., Indianapolis, IN, USA), a polyclonal sheep anti-DIG Fab fragment conjugated with alkaline phosphatase (DIG Nucleic Acid Detection Kit; Boehringer Mannheim Corp.) and alkaline phosphatase solution containing levamisole as described.26 Slides were counterstained with nuclear fast red, dehydrated and mounted using Permount (Sigma-Aldrich, St Louis, MO, USA).
Whole-mount analysis of mammary glands
Carmine-alum staining was performed as described.19 Briefly, dissected mammary glands were spread onto glass slides, fixed with Carnoy’s fixative (60% ethanol, 30% chloroform and 10% glacial acetic acid) for overnight, hydrated, stained overnight in 0.2% carmine and 0.5% AlK(SO4)2, dehydrated in graded solutions of ethanol, cleared in Histoclear (Shandon Inc., Pittsburgh, PA, USA) and mounted.
Histological and immunofluorescence analysis
Paraffin sections (5-μm thick) were used for HE, Masson’s trichrome or Sirius red staining according to the manufacturer’s instructions. Immunofluorescence experiments were performed, as previously described,19, 30 using primary antibodies directed against E-cadherin (Zymed Laboratories Inc., San Francisco, CA, USA), anti-αSMA (Sigma-Aldrich), anti-keratin 14 (Eurogentec, Angers, France), anti-keratin 8/18 (Spring Biosciences, Pleasanton, CA, USA), anti-CD31 (BD Bioscience, Le Pont de Claix, France), anti-vimentin (Fitzgerald Industries International, Acton, MA, USA) at recommended concentrations. After washing, slides were incubated with fluorescent Alexa-488- and Cy3-conjugated secondary antibodies (Jackson Immuno Research, West Grove, PA, USA). Then DAPI-Mounting medium was used and sections were observed under fluorescence microscope (Invitrogen) or confocal fluorescence microscope (Leica Microsystem, Wetzlar, Germany). Image analysis was performed using ImageJ (National Institute of Health, Bethesda, MD, USA).
Cleared fat pad transplantation system
As previously described,19, 53 reciprocal mammary epithelial transplantations were done between 3-week-old WT and MMP-11−/− littermate females. Mice were anaesthetized by intraperitoneal injection of drug (xylazine/ketamine) and inguinal mammary glands were exposed through small incisions in the skin of the lower abdomen and along the right hind leg. For fat pad clearing, the connection between glands #4 and #5 was disrupted by ligation and endogenous epithelia were removed by removing the region between the nipple and the line above the lymph node. Removed glands (∼2 mm3) were cut into small pieces and transplanted into the cleared fat pads of recipient mice (Supplementary Figures S2A and B). The left inguinal (#4) mammary glands were used as controls. Transplanted mice were killed 3, 6 and 9 weeks after the operation. At least three sets of transplantation (four WT and four MMP-11−/− mice per set) were performed for each experimental condition and reproducible results were obtained from all of them.
MMP-11 recombinant proteins
Active and inactive (Glu220Ala) MMP-11 recombinant proteins (rMMP-11a and rMMP-11i, respectively) were produced as previously described.38, 54 Their enzymatic activities were controlled using the indirect quantitative colorimetric substrate assay with a1-PI.55 Briefly, MMPs (50–400 ng) were incubated at 22 °C with a1-PI (final concentration 30 μg/ml 50 mM Tris (pH 7.5), 200 mM NaCl, 10 mM CaCl2, 0.1% Triton X-100). The a1-PI cleavage was stopped before addition of a-Chymotrypsin (a-CT; final concentration 2.4 μg/ml). Chromogenic synthetic substrate of a-CT (N-succinyl-Ala-Ala-Pro-Phe-p-nitroanilide) was added (final concentration 140 μg/ml). The reaction was stopped by phenylmethylsulfonyl fluoride and the optical density at 405 nm was determined (Yellow; Beckman DU640 spectrophotometer, Fullerton, CA, USA).
In vitro culture of mouse mammary epithelial organoids
As described,56 the #4 inguinal mammary glands were removed from 14-week-old virgin mice. The lymph nodes were removed. The mammary glands minced with a razor blade were incubated in a collagenase/trypsin mixture. After centrifugation (1000 r.p.m., 10 min), the pellet was re-suspended in DMEM/F12 and treated with DNase (2 U/μl). The epithelial pieces were separated from the single cells through a series of differential centrifugation. The final pellet containing organoids was re-suspended in Matrigel and distributed in 96-well culture plates containing an underlay Matrigel layer (Growth Factor Reduced Matrigel, BD Biosciences, San Jose, CA, USA). After 24 h incubation at 37 °C into serum-free basal medium (DMEF/F12, 1% ITS (insulin/transferrin/selenium), 1% penicillin/streptomycin), appropriate medium (alone or containing rMMP-11) was added and changed daily. Experiments were carried out in triplicates and three independent sets of experiments were performed for each condition. Positive control corresponds to organoids treated with human TGF alpha (9 nM) (Sigma) as previously described.37
Statistical analysis
Results are represented as means±standard deviation (s.d.). Paired and two-tailed Student’s t-tests or analysis of variance. A P-value <0.05 was considered statistically significant (*P<0.05; **P<0.01; ***P<0.001).
References
Brinckerhoff CE, Matrisian LM . Matrix metalloproteinases: a tail of a frog that became a prince. Nat Rev Mol Cell Biol 2002; 3: 207–214.
Maskos K, Bode W . Structural basis of matrix metalloproteinases and tissue inhibitors of metalloproteinases. Mol Biotechnol 2003; 25: 241–266.
Basset P, Bellocq JP, Wolf C, Stoll I, Hutin P, Limacher JM et al. A novel metalloproteinase gene specifically expressed in stromal cells of breast carcinomas. Nature 1990; 348: 699–704.
Andarawewa KL, Motrescu ER, Chenard MP, Gansmuller A, Stoll I, Tomasetto C et al. Stromelysin-3 is a potent negative regulator of adipogenesis participating to cancer cell-adipocyte interaction/crosstalk at the tumor invasive front. Cancer Res 2005; 65: 10862–10871.
Basset P, Bellocq JP, Lefebvre O, Noel A, Chenard MP, Wolf C et al. Stromelysin-3: a paradigm for stroma-derived factors implicated in carcinoma progression. Crit Rev Oncol Hematol 1997; 26: 43–53.
Motrescu ER, Rio MC . Cancer cells, adipocytes and matrix metalloproteinase 11: a vicious tumor progression cycle. Biol Chem 2008; 389: 1037–1041.
Wasenius VM, Hemmer S, Kettunen E, Knuutila S, Franssila K, Joensuu H . Hepatocyte growth factor receptor, matrix metalloproteinase-11, tissue inhibitor of metalloproteinase-1, and fibronectin are up-regulated in papillary thyroid carcinoma: a cDNA and tissue microarray study. Clin Cancer Res 2003; 9: 68–75.
Kren L, Goncharuk VN, Krenova Z, Stratil D, Hermanova M, Skrickova J et al. Expression of matrix metalloproteinases 3, 10 and 11 (stromelysins 1, 2 and 3) and matrix metalloproteinase 7 (matrilysin) by cancer cells in non-small cell lung neoplasms. Clinicopathologic studies. Cesk Patol 2006; 42: 16–19.
Zhao ZS, Chu YQ, Ye ZY, Wang YY, Tao HQ . Overexpression of matrix metalloproteinase 11 in human gastric carcinoma and its clinicopathologic significance. Hum Pathol 2010; 41: 686–696.
Pedersen G, Saermark T, Kirkegaard T, Brynskov J . Spontaneous and cytokine induced expression and activity of matrix metalloproteinases in human colonic epithelium. Clin Exp Immunol 2009; 155: 257–265.
Kwon YJ, Hurst DR, Steg AD, Yuan K, Vaidya KS, Welch DR et al. Gli1 enhances migration and invasion via up-regulation of MMP-11 and promotes metastasis in ERalpha negative breast cancer cell lines. Clin Exp Metastasis 2011; 28: 437–449.
Noel AC, Lefebvre O, Maquoi E, VanHoorde L, Chenard MP, Mareel M et al. Stromelysin-3 expression promotes tumor take in nude mice. J Clin Invest 1996; 97: 1924–1930.
Wu E, Mari BP, Wang F, Anderson IC, Sunday ME, Shipp MA . Stromelysin-3 suppresses tumor cell apoptosis in a murine model. J Cell Biochem 2001; 82: 549–555.
Tan J, Buache E, Chenard MP, Dali-Youcef N, Rio MC . Adipocyte is a non-trivial, dynamic partner of breast cancer cells. Int J Dev Biol 2011; 55: 851–859.
Couldrey C, Moitra J, Vinson C, Anver M, Nagashima K, Green J . Adipose tissue: a vital in vivo role in mammary gland development but not differentiation. Dev Dyn 2002; 223: 459–468.
Landskroner-Eiger S, Park J, Israel D, Pollard JW, Scherer PE . Morphogenesis of the developing mammary gland: stage-dependent impact of adipocytes. Dev Biol 2010; 344: 968–978.
Ching S, Kashinkunti S, Niehaus MD, Zinser GM . Mammary adipocytes bioactivate 25-hydroxyvitamin D(3) and signal via vitamin D(3) receptor, modulating mammary epithelial cell growth. J Cell Biochem 2011; 112: 3393–3405.
Pavlovich AL, Manivannan S, Nelson CM . Adipose stroma induces branching morphogenesis of engineered epithelial tubules. Tissue Eng Part A 2010; 16: 3719–3726.
Ucar A, Vafaizadeh V, Jarry H, Fiedler J, Klemmt PA, Thum T et al. miR-212 and miR-132 are required for epithelial stromal interactions necessary for mouse mammary gland development. Nat Genet 2010; 42: 1101–1108.
McCave EJ, Cass CA, Burg KJ, Booth BW . The normal microenvironment directs mammary gland development. J Mammary Gland Biol Neoplasia 2010; 15: 291–299.
Naylor MJ, Ormandy CJ . Mouse strain-specific patterns of mammary epithelial ductal side branching are elicited by stromal factors. Dev Dyn 2002; 225: 100–105.
Khokha R, Werb Z . Mammary gland reprogramming: metalloproteinases couple form with function. Cold Spring Harb Perspect Biol 2011; 3: a004333.
Hovey RC, Aimo L . Diverse and active roles for adipocytes during mammary gland growth and function. J Mammary Gland Biol Neoplasia 2010; 15: 279–290.
Hennighausen L, Robinson GW . Signaling pathways in mammary gland development. Dev Cell 2001; 1: 467–475.
Green KA, Lund LR . ECM degrading proteases and tissue remodelling in the mammary gland. Bioessays 2005; 27: 894–903.
Masson R, Lefebvre O, Noel A, Fahime ME, Chenard MP, Wendling C et al. In vivo evidence that the stromelysin-3 metalloproteinase contributes in a paracrine manner to epithelial cell malignancy. J Cell Biol 1998; 140: 1531–1541.
Lefebvre O, Wolf C, Limacher JM, Hutin P, Wendling C, LeMeur M et al. The breast cancer-associated stromelysin-3 gene is expressed during mouse mammary gland apoptosis. J Cell Biol 1992; 119: 997–1002.
Witty JP, Wright JH, Matrisian LM . Matrix metalloproteinases are expressed during ductal and alveolar mammary morphogenesis, and misregulation of stromelysin-1 in transgenic mice induces unscheduled alveolar development. Mol Biol Cell 1995; 6: 1287–1303.
Sternlicht MD . Key stages in mammary gland development: the cues that regulate ductal branching morphogenesis. Breast Cancer Res 2006; 8: 20129.
Kurley SJ, Bierie B, Carnahan RH, Lobdell NA, Davis MA, Hofmann I et al. p120-catenin is essential for terminal end bud function and mammary morphogenesis. Development 2012; 139: 1754–1764.
Mailleux AA, Overholtzer M, Schmelzle T, Bouillet P, Strasser A, Brugge JS . BIM regulates apoptosis during mammary ductal morphogenesis, and its absence reveals alternative cell death mechanisms. Dev Cell 2007; 12: 221–234.
Cannata D, Lann D, Wu Y, Elis S, Sun H, Yakar S et al. Elevated circulating IGF-I promotes mammary gland development and proliferation. Endocrinology 2010; 151: 5751–5761.
Lijnen HR, Van HB, Frederix L, Rio MC, Collen D . Adipocyte hypertrophy in stromelysin-3 deficient mice with nutritionally induced obesity. Thromb Haemost 2002; 87: 530–535.
Deome KB, Faulkin LJ Jr., Bern HA, Blair PB . Development of mammary tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice. Cancer Res 1959; 19: 515–520.
Edwards PA . The use of transplanted mammary gland to study cancer signalling pathways. Adv Exp Med Biol 2000; 480: 163–167.
Sternlicht MD, Sunnarborg SW, Kouros-Mehr H, Yu Y, Lee DC, Werb Z . Mammary ductal morphogenesis requires paracrine activation of stromal EGFR via ADAM17-dependent shedding of epithelial amphiregulin. Development 2005; 132: 3923–3933.
Fata JE, Mori H, Ewald AJ, Zhang H, Yao E, Werb Z et al. The MAPK(ERK-1,2) pathway integrates distinct and antagonistic signals from TGFalpha and FGF7 in morphogenesis of mouse mammary epithelium. Dev Biol 2007; 306: 193–207.
Kannan R, Ruff M, Kochins JG, Manly SP, Stoll I, El Fahime M et al. Purification of active matrix metalloproteinase catalytic domains and its use for screening of specific stromelysin-3 inhibitors. Protein Expr Purif 1999; 16: 76–83.
Hinck L, Silberstein GB . Key stages in mammary gland development: the mammary end bud as a motile organ. Breast Cancer Res 2005; 7: 245–251.
Nelson CM, Vanduijn MM, Inman JL, Fletcher DA, Bissell MJ . Tissue geometry determines sites of mammary branching morphogenesis in organotypic cultures. Science 2006; 314: 298–300.
Ewald AJ, Brenot A, Duong M, Chan BS, Werb Z . Collective epithelial migration and cell rearrangements drive mammary branching morphogenesis. Dev Cell 2008; 14: 570–581.
Simian M, Hirai Y, Navre M, Werb Z, Lochter A, Bissell MJ . The interplay of matrix metalloproteinases, morphogens and growth factors is necessary for branching of mammary epithelial cells. Development 2001; 128: 3117–3131.
Mori H, Lo AT, Inman JL, Alcaraz J, Ghajar CM, Mott JD et al. Transmembrane/cytoplasmic, rather than catalytic, domains of Mmp14 signal to MAPK activation and mammary branching morphogenesis via binding to integrin beta1. Development 2013; 140: 343–352.
Keely PJ, Wu JE, Santoro SA . The spatial and temporal expression of the alpha 2 beta 1 integrin and its ligands, collagen I, collagen IV, and laminin, suggest important roles in mouse mammary morphogenesis. Differentiation 1995; 59: 1–13.
Dhimolea E, Maffini MV, Soto AM, Sonnenschein C . The role of collagen reorganization on mammary epithelial morphogenesis in a 3D culture model. Biomaterials 2010; 31: 3622–3630.
Motrescu ER, Blaise S, Etique N, Messaddeq N, Chenard MP, Stoll I et al. Matrix metalloproteinase-11/stromelysin-3 exhibits collagenolytic function against collagen VI under normal and malignant conditions. Oncogene 2008; 27: 6347–6355.
Aigner T, Hambach L, Soder S, Schlotzer-Schrehardt U, Poschl E . The C5 domain of Col6A3 is cleaved off from the Col6 fibrils immediately after secretion. Biochem Biophys Res Commun 2002; 290: 743–748.
Lamande SR, Morgelin M, Adams NE, Selan C, Allen JM . The C5 domain of the collagen VI alpha3(VI) chain is critical for extracellular microfibril formation and is present in the extracellular matrix of cultured cells. J Biol Chem 2006; 281: 16607–16614.
Wolf K, Friedl P . Extracellular matrix determinants of proteolytic and non-proteolytic cell migration. Trends Cell Biol 2011; 21: 736–744.
Ha HY, Moon HB, Nam MS, Lee JW, Ryoo ZY, Lee TH et al. Overexpression of membrane-type matrix metalloproteinase-1 gene induces mammary gland abnormalities and adenocarcinoma in transgenic mice. Cancer Res 2001; 61: 984–990.
Wiseman BS, Sternlicht MD, Lund LR, Alexander CM, Mott J, Bissell MJ et al. Site-specific inductive and inhibitory activities of MMP-2 and MMP-3 orchestrate mammary gland branching morphogenesis. J Cell Biol 2003; 162: 1123–1133.
Lanigan F, O'Connor D, Martin F, Gallagher WM . Molecular links between mammary gland development and breast cancer. Cell Mol Life Sci 2007; 64: 3159–3184.
Khialeeva E, Lane TF, Carpenter EM . Disruption of reelin signaling alters mammary gland morphogenesis. Development 2011; 138: 767–776.
Noel A, Boulay A, Kebers F, Kannan R, Hajitou A, Calberg-Bacq CM et al. Demonstration in vivo that stromelysin-3 functions through its proteolytic activity. Oncogene 2000; 19: 1605–1612.
Holtz B, Cuniasse P, Boulay A, Kannan R, Mucha A, Beau F et al. Role of the S1' subsite glutamine 215 in activity and specificity of stromelysin-3 by site-directed mutagenesis. Biochemistry 1999; 38: 12174–12179.
Lo AT, Mori H, Mott J, Bissell MJ . Constructing three-dimensional models to study mammary gland branching morphogenesis and functional differentiation. J Mammary Gland Biol Neoplasia 2012; 17: 103–110.
Acknowledgements
We thank Susan Chan and Gérard Gradwohl for helpful discussion, and Corinne Wendling, Laure Nivlet and Olivia Wendling for technical assistance. This work was supported by funds from the Institut National de la Santé et de la Recherche Médicale, the Centre National de la Recherche Scientifique, the Hôpital Universitaire de Strasbourg, the Association pour la Recherche sur le Cancer, the Ligue Nationale Française Contre le Cancer (LNCC; Comités du Haut-Rhin, Bas-Rhin et de la Haute-Savoie) and the Institut National du Cancer (INCA PL-BIO ADIPOK 2011–2013). JT was a recipient of a Chinese fellowship of National Natural Science Foundation of China (number: 81102008); EB was a recipient of LNCC and INCA fellowships.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Rights and permissions
This work is licensed under a Creative Commons Attribution 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by/3.0/
About this article
Cite this article
Tan, J., Buache, E., Alpy, F. et al. Stromal matrix metalloproteinase-11 is involved in the mammary gland postnatal development. Oncogene 33, 4050–4059 (2014). https://doi.org/10.1038/onc.2013.434
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2013.434
Keywords
This article is cited by
-
Fibroblast-derived CXCL12 promotes breast cancer metastasis by facilitating tumor cell intravasation
Oncogene (2018)
-
Stromal factors involved in human prostate cancer development, progression and castration resistance
Journal of Cancer Research and Clinical Oncology (2017)
-
Matrix metalloproteinase 11 protects from diabesity and promotes metabolic switch
Scientific Reports (2016)
-
Remodelling the extracellular matrix in development and disease
Nature Reviews Molecular Cell Biology (2014)

{kind=link}
{kind=link}
{kind=link}
{kind=link}