

Abstract
Macromolecules such as proteins are attracting increasing interest for molecular imaging. We previously proposed a novel strategy for preparing macromolecules labeled with a PET radionuclide, 11C, using a cell-free translation system with 11C-methionine. However, macromolecules tend to exhibit slower kinetics, thus requiring a longer scanning time. Here, we expand our strategy using 18F, which has a longer half-life, with the cell-free translation system with 4-18F-fluoro-l-proline (18F-FPro). We evaluated 18F-interleukin-8 (18F-IL-8) produced by this method in vitro and in vivo to provide a proof of concept of our strategy. Methods: We tested some fluorinated amino acids to be incorporated into a protein. Trans-18F-FPro was radiolabeled from the corresponding precursor. 18F-IL-8 was produced using the cell-free translation system with trans-18F-FPro instead of natural l-proline with incubation at 37°C for 120 min. An in vitro binding assay of 18F-IL-8 was performed using IL-8 receptor–expressing cells. After intravenous administration of 18F-IL-8, in vivo PET imaging of IL-8 receptor–expressing xenograft-bearing mice was performed using a small-animal PET system. Results: FPro was identified as an amino acid incorporated into the protein. 18F-IL-8 was successfully prepared using the cell-free translation system and trans-18F-FPro with the radiochemical yield of 1.5% (decay-corrected) based on trans-18F-FPro. In vitro binding assays of 18F-IL-8 demonstrated its binding to IL-8 receptor. In vivo PET imaging demonstrated that 18F-IL-8 clearly accumulated in IL-8 receptor–expressing xenografts in mice, unlike trans-18F-FPro. Conclusion: 18F-IL-8 produced by this method binds to IL-8 receptors in vitro, and 18F-IL-8 PET clearly visualizes its target receptor–expressing xenograft in vivo. Therefore, this technique might be useful for labeling macromolecules and performing preclinical evaluations of proteins of interest in vitro and in vivo.
- PET
- 18F
- cell-free protein synthesis
- non-natural amino acid
- interleukin-8
PET is a noninvasive molecular imaging technique that visualizes physiologic and pathologic functions using PET radiopharmaceuticals. PET is a highly sensitive and quantitative technique, providing functional information in living subjects from small animals to humans. Although small organic compounds labeled with PET radionuclides, such as 18F-fludeoxyglucose, are widely used in clinical practice, bioactive macromolecules such as proteins are attracting increasing interest as PET probes for molecular imaging in order to characterize their pharmacokinetics, potentially diagnose diseases, and evaluate therapeutic efficacy (1,2). Because bioactive macromolecules possess high binding affinity and specificity for their targets in vivo, radiolabeled bioactive macromolecules are promising candidates for PET imaging radiotracers.
Recent biotechnologic advances have enabled the simple preparation of various proteins using cell-free translation systems that contain enzymes and essential factors for protein synthesis derived from Escherichia coli, wheat germ, insect, and mammalian cells (3,4); these methods are now commercially available for research purposes. We previously proposed a unique approach for preparing 11C-labeled proteins using a cell-free translation system with a natural amino acid, 11C-l-methionine (5,6). The advantage of this method is its simplicity regarding preparation of proteins of interest labeled with 11C using only their template DNA. Furthermore, carbon is generally included in proteins as a component of natural amino acids, permitting the use of the same scaffold of original proteins without any additions. However, the main limitation of this method is the short half-life of 11C (20.4 min). In general, macromolecules tend to exhibit slower pharmacokinetics than small molecules, thus requiring a longer scanning time. Therefore, 11C labeling may be inappropriate for proteins with slow pharmacokinetics.
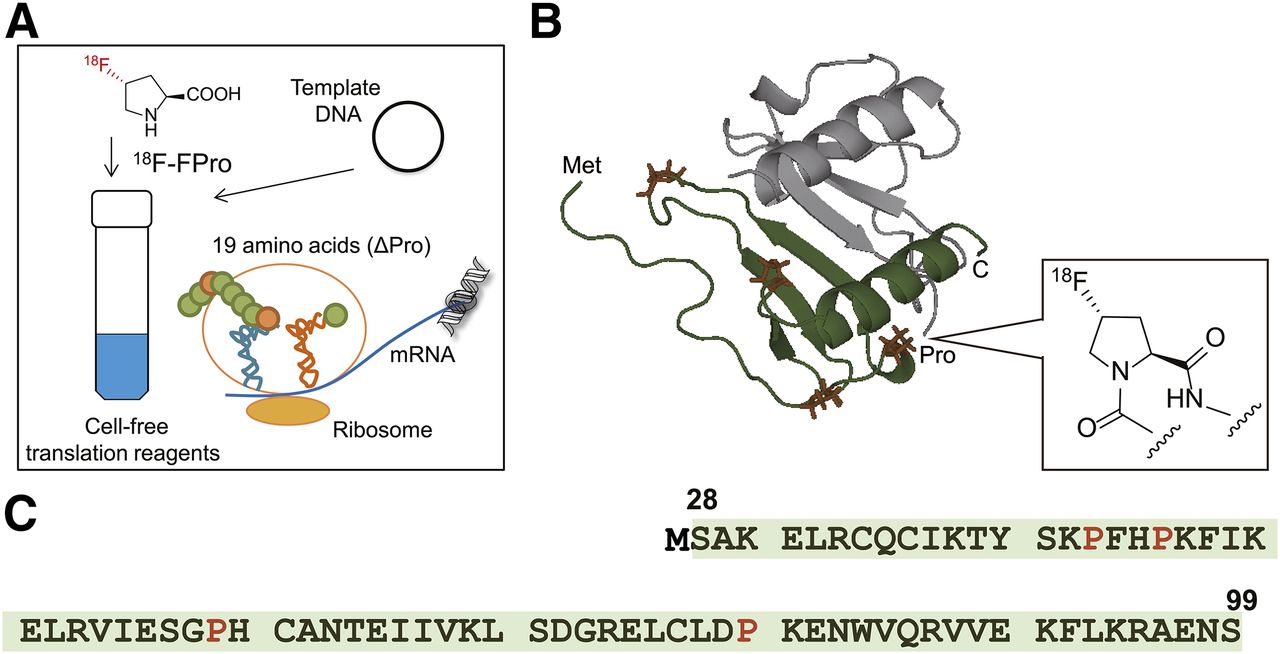
Accordingly, we expanded our approach by preparing radiolabeled proteins with 18F, which has a longer half-life (109.8 min), making it much easier to use for in vivo imaging (7), and using a cell-free translation system with an 18F-labeled amino acid (Fig. 1A). However, incorporation of nonnatural amino acid into proteins is generally unlikely because of the high substrate specificity of aminoacyl-transfer RNA (tRNA) synthase (8). Although some approaches have been proposed to incorporate nonnatural amino acids into proteins using cell-free translation systems with chemically synthesized amber suppressor tRNA-nonnatural amino acid, unnatural base pair, engineered orthogonal amber suppressor tRNA/aminoacyl-tRNA synthase pair, and an amino acid depleted system (9–12), it is unlikely that they are suitable for labeling of PET radionuclides because of their complexity and time-consuming procedure. Therefore, searching for nonnatural amino acids that can be directly incorporated into proteins by a cell-free translation system derived from E. coli without any additional components and easily labeled with 18F led to the identification of 4-fluoro-l-proline (FPro) being incorporated into proteins instead of natural l-proline (Pro). Here, we report the successful preparation of 18F-labeled interleukin-8 (18F-IL-8) as a model using a commercially available cell-free translation system with 18F-FPro and investigated the usefulness of 18F-IL-8 in vivo as a PET tracer to provide a proof of concept of our strategy.

(A) Schematic illustration of proposed 18F-labeled protein preparation system using a cell-free translation system and trans-4-18F-fluoro-l-proline (trans-18F-FPro). Three-dimensional structure of 18F-IL-8 (B) and amino acid sequence of IL-8 (C). Cartoon representation of 18F-IL-8 structure (28–99) showing dimeric structure (PDBID: 1IL-8) (35). Green represents 1 monomer unit of dimeric structure. 18F-FPro was site-specifically incorporated into l-proline residue (orange) instead of natural l-proline. Because IL-8 is a secreted protein (signal sequence, 1–20 amino acids), a methionine (MET) residue was added to N terminus of each IL-8 sequence to initiate translation (Met-28–99). Colors of cartoon of IL-8 structure correspond to highlighted amino acids sequences (green highlights and orange letters).
MATERIALS AND METHODS
Materials
The PURExpress Δ(aa, tRNA) kit, PURExpress Disulfide Bond Enhancer, and murine RNase inhibitor were purchased from New England BioLabs. All amino acids were obtained from Wako Chemicals. pET-28a and pF5K CMV-neo Flexi vectors were purchased from Novagen and Promega, respectively. The IL-8 gene was custom-synthesized by Genescript. IL-8 gene was subcloned into pET-28a expression vector. The IL-8 receptor α (IL-8 RA) gene (GenBank accession no. AB464598) was obtained from Promega. IL-8RA gene was subcloned into pF5K CMV-neo Flexi vector for the expression. 125I-labeled IL-8 (human, recombinant) was obtained from Perkin Elmer and Analytic Science (specific activity, 81.4 TBq/mmol). (2S, 4S)-4-fluoro-l-proline (cis-FPro) and (2S, 4R)-4-fluoro-l-proline (trans-FPro) were purchased from Watanabe Chemical Industries. O-fluoroethyl-l-tyrosine was purchased from ABX GmbH. O-fluoromethyl-L-tyrosine was synthesized as previously described (13).
Synthesis of IL-8–Containing FPro Using PURExpress
We synthesized IL-8 using PURExpress, which is based on the PURESYSTEM previously used (5), because PURExpress can easily be customized with respect to the components of the reaction solution. IL-8–containing FPro or fluorinated tyrosine derivatives were synthesized by the addition of pET-28a with IL-8, 19 amino acids, and FPro or fluorinated tyrosine derivatives instead of Pro or tyrosine, respectively, to the kit reaction solution containing E. coli 70S ribosome, nucleoside triphosphates, E. coli tRNA, and an energy recycling system. In addition, we added RNase inhibitor and disulfide bond enhancer to protect against RNA degradation and enhance protein folding, respectively. After incubation at 37°C for 120 min, IL-8 was purified using a strong cation exchange spin column as described previously (5). To confirm IL-8 production, proteins were resolved in NuPAGE Novex 4%–12% Bis-Tris gel using NuPAGE MES SDS Running Buffer (Life Technologies), transferred to a polyvinylidene fluoride membrane, and probed with anti–IL-8 antibody (1:1000; MBL) and antirabbit secondary antibody (1:20,000; Thermo Fisher Scientific). Blots were developed with ECL Western Blotting Detection Reagents (GE Healthcare) and exposed in x-ray film (Fujifilm). After Zip TipC18 (Millipore) treatment, mass spectrometry was performed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (AXIMA Performance; Shimadzu) to confirm FPro incorporation. The amounts of synthesized IL-8, FPro-IL-8, and 18F-IL-8 were determined using an IL-8 enzyme-linked immunosorbent assay kit (Toyobo).
IL-8 Receptor Binding Assay
Competitive binding assays were performed on HEK293 cells transiently expressing IL-8 RA in 24-well dishes. The cells were washed with binding medium (RPMI1640, 1% bovine serum albumin, 25 mM 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid [HEPES]), and 200 μL of 0.06 nM 125I-IL-8 were added along with increasing concentrations of unlabeled synthesized wild-type IL-8 and its FPro variants by PURExpress. After incubation at 37°C for 1 h, the cells were washed 3 times with assay buffer (phosphate-buffered saline plus Mg2+ and Ca2+), harvested with radioimmunoprecipitation assay buffer, and measured in a γ-counter (AccuFLEX γ7000; Aloka). All experiments were performed in triplicate. The half maximal inhibitory concentration (IC50) values were determined by nonlinear regression analysis (GraphPad Prism, version 5.0; GraphPad Software).
Radiosynthesis of Trans-4-18F-Fluoro-l-Proline
No-carrier-added 18F-fluoride was produced by the 18O(p, n)18F reaction on enriched 18O-H2O (Taiyo Nippon Sanso) with a HM-12 cyclotron (Sumitomo Heavy Industries). The specific activity of 18F-fluoride was usually in the range of 100–400 GBq/μmol at the end of bombardment. The precursor of trans-4-18F-fluoro-l-proline (trans-18F-FPro) was prepared as described previously (14). Trans-4-18F-FPro was radiosynthesized using the corresponding tosylate precursor (3 mg). Aqueous 18F− contained in K2CO3 solution (1.0–1.5 GBq, 0.5 mL) and Kryptofix222 (16 mg) were placed in a vial. Water was removed by azeotropic evaporation with acetonitrile. After being dried, the activated 18F-KF/Kryptofix222 was reacted with the precursor (3 mg) in dimethyl sulfoxide (0.7 mL) at 110°C for 10 min. Then, 1 M NaOH (0.4 mL) was added to the solution, followed by additional deprotection of the methyl ester group at 110°C for 10 min. After neutralization by the addition of 1 M HCl (0.5 mL), the resultant solution was diluted with water (6 mL) and subsequently extracted with an activated Sep-Pak tC18 cartridge (Waters). After being dried in ethanol by azeotropic evaporation at 110°C, 2 M HCl (0.4 mL) was added to the solution and reacted at 110°C for 5 min to remove the tert-butoxycarbonyl protective group. After neutralization with 4 M potassium acetate (0.2 mL), the resultant solution was dried by azeotropic evaporation with acetonitrile (3 mL) at 110°C and dissolved in ethanol (0.6 mL). After filtration with a Syringe Filter (Whatman), the crude product was purified by semipreparative reversed-phase high-performance liquid chromatography (column: YMC-Pack Polyamine II, s-5 μm, 250 × 10 mm [YMC Co., Ltd.]; mobile phase: CH3OH/H2O = 65/35; 4 mL/min; ultraviolet: 230 nm). After extraction of the purified product with a Sep-Pak tC18 cartridge and ethanol evaporation, the purified product was dissolved in distilled water for the protein synthesis reaction. The overall synthesis time was approximately 60 min. The radiochemical purity exceeded 95% (Supplemental Fig. 1; supplemental materials are available at http://jnm.snmjournals.org). The decay-corrected radiochemical yields of trans-18F-FPro were 30%–40%. Because of the small size of the ultraviolet peak, the specific activity of trans-18F-FPro was estimated to 74 GBq/mmol as a lower limit on specific activity using a calibration curve for the sensitivity of the ultraviolet peak, although the average specific activity of 18F ligands with general ultraviolet absorbance is at least more than 37 GBq/μmol at the end of the synthesis (Supplemental Fig. 1).
Radiosynthesis of IL-8–Containing Trans-18F-FPro Using PURExpress
For 250 μL scale, no-carrier-added trans-18F-FPro (11.4–151.7 MBq, 50 μL) and pET-28a with IL-8 (500 ng) dissolved in distilled water were added to the described kit reaction solution except for Pro. The reaction mixture was reacted at 37°C for 15, 30, 60, 90, and 120 min. To confirm 18F-IL-8 production, proteins were resolved in 4%–12% gradient gels and transferred to a polyvinylidene fluoride membrane, and the gel and transferred membrane were subsequently exposed to a BAS-MS2025 imaging plate (Fujifilm) for 90 min. Autoradiograms were obtained using a BAS-5000 phosphor imaging instrument (Fujifilm). After radioactive decay, the membrane was subjected to Western blotting analysis as described above. The radiochemical purity of 18F-IL-8 was determined by analyzing the gel autoradiograms. After incubation at 37°C for 120 min, 18F-IL-8 was purified as described above, and radioactivity was measured in a γ-counter (AccuFLEX γ7000) to determine the radiochemical yield. The eluted 18F-IL-8 was diluted in water for animal experiments.
In Vitro Binding of 18F-Labeled IL-8
In vitro binding of 18F-IL-8 to IL-8 receptor was performed on Chinese hamster ovary (CHO) cells (2.0 × 105 cells/well) transfected with pF5K CMV-neo containing control or IL-8 RA vector in 24-well dishes. The cells were washed with binding medium (RPMI1640, 1% bovine serum albumin, and 25 mM HEPES), and 200 μL of 18F-IL-8 (14.8 kBq/mL) were added. After incubation at room temperature for 1 h, the cells were washed 3 times with assay buffer, harvested with 0.1 M NaOH, and measured in a γ-counter. All experiments were performed in triplicate.
Small-Animal PET Studies
All animal experimental protocols described herein were approved by the Laboratory Animal Care Committee of Tohoku University. In vivo PET studies were performed using male SLC:ICR mice for the biodistribution study and male immunodeficiency mice (BALB/c-ν: CAnN.Cg-Foxn1Nu/CrlCrlj) with IL-8 RA stably expressing CHO xenografts for the receptor imaging study. Mice were inoculated under the arm by subcutaneous injections of 106 cells, and IL-8–expressing CHO xenografts were allowed to grow for 4 wk. 18F-IL-8 or trans-18F-FPro was administered intravenously (0.2–1.1 MBq/mouse, ∼0.35 mL in distilled water containing NaCl). Dynamic PET images were acquired using a G4 PET/x-ray scanner (PerkinElmer) at 10 min after radiotracer injection (18F-IL-8 or trans-18F-FPro) for 120 min under 1.5% isoflurane anesthesia. Images were analyzed by AMIDE software (15).
RESULTS
Synthesis of IL-8–Containing FPro by PURExpress
Western blot analysis demonstrated that the cell-free reaction mixtures containing FPro stereoisomers including cis-FPro and trans-FPro, template DNA, and PURExpress reagents without natural Pro successfully produced IL-8 (Fig. 2A), but not fluoro-l-tyrosine derivatives, instead of natural tyrosine (Supplemental Fig. 2). There were no significant differences in the amounts of synthesized IL-8 (∼25 μg, 125-μL reaction scale) determined by enzyme-linked immunosorbent assay between FPro stereoisomers. In addition, MALDI-TOF MS analysis showed that the molecular weight of IL-8 containing each FPro stereoisomer was 73 Da heavier than wild-type IL-8, which is equivalent to 4 fluorine (molecular weight, 19) minus 4 hydrogen (molecular weight, 1) (Fig. 2B). Both cis- and trans-FPro were incorporated into the protein as previously reported (14,16).
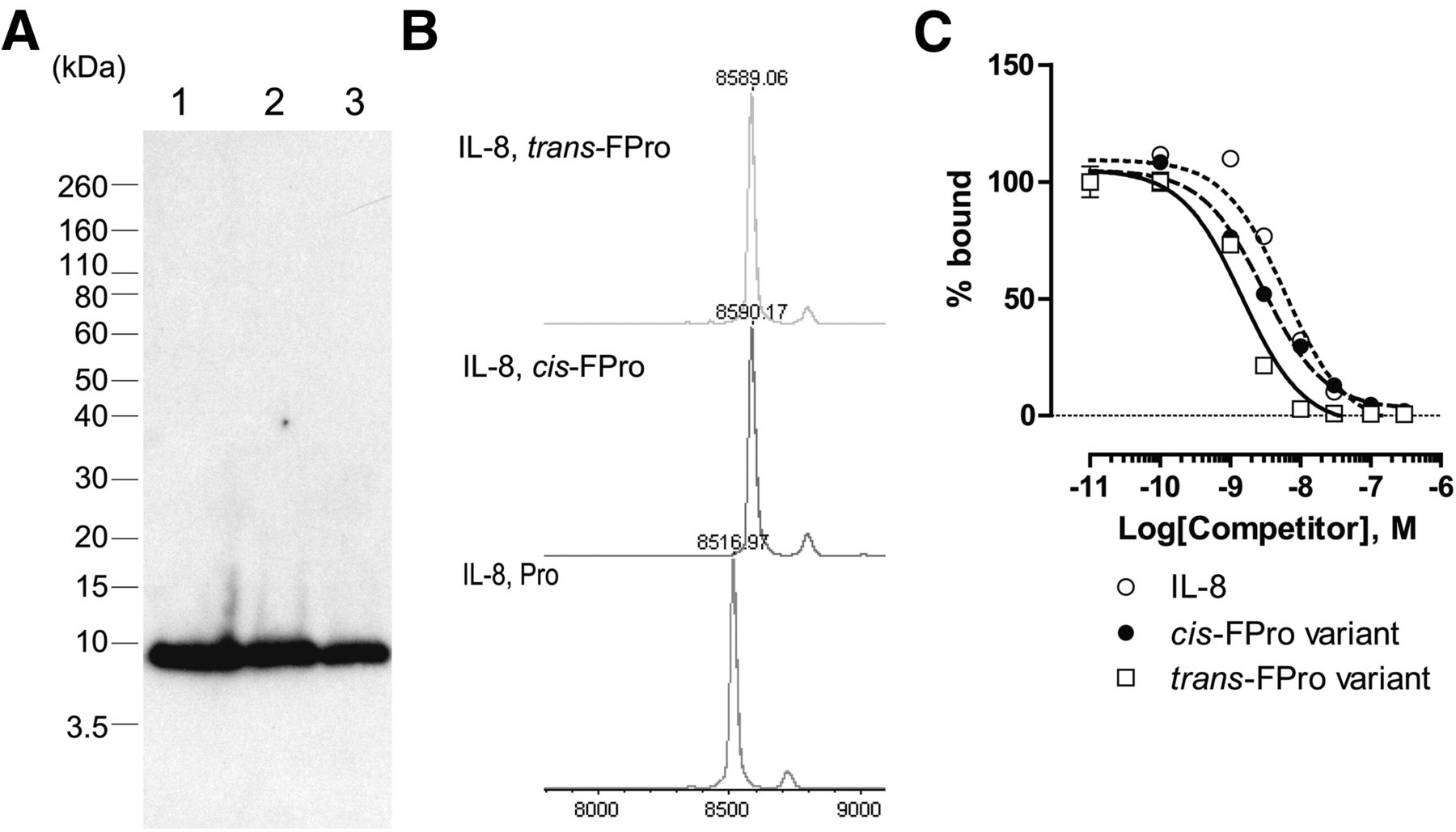
IL-8 synthesis using PURExpress system and nonnatural amino acids such as FPro. (A) Western blot analysis of IL-8. Lane 1: PURExpress with Pro and IL-8 DNA template. Lane 2: PURExpress with cis-FPro and IL-8 DNA template. Lane 3: PURExpress with trans-FPro and IL-8 DNA template. (B) MALDI-TOF MS. Competitive binding of wild-type IL-8 and its cis-FPro and trans-FPro variants to IL-8 RA–expressing cells (C). 125I-IL-8 was used as a radioactive tracer.
Binding Assay of HEK293 Cells Expressing IL-8 Receptor
In vitro competitive binding analysis of 125I-IL-8 to IL-8 RA–expressing HEK293 cells was performed. Wild-type IL-8 and its FPro variants completely inhibited 125I-IL-8 binding to IL-8 RA (Fig. 2C). In addition, the trans-FPro IL-8 variant bound to IL-8 RA with higher affinity (IC50 = 1.5 ± 0.1 nM) than the cis-FPro IL-8 variant (IC50 = 3.0 ± 0.3 nM) and wild type (IC50 = 6.0 ± 1.5 nM). Therefore, we chose trans-FPro, which possesses higher binding affinity for IL-8 RA, as an 18F-labeled amino acid source for further investigation.
Radiosynthesis of IL-8–Containing Trans-18F-FPro by PURExpress
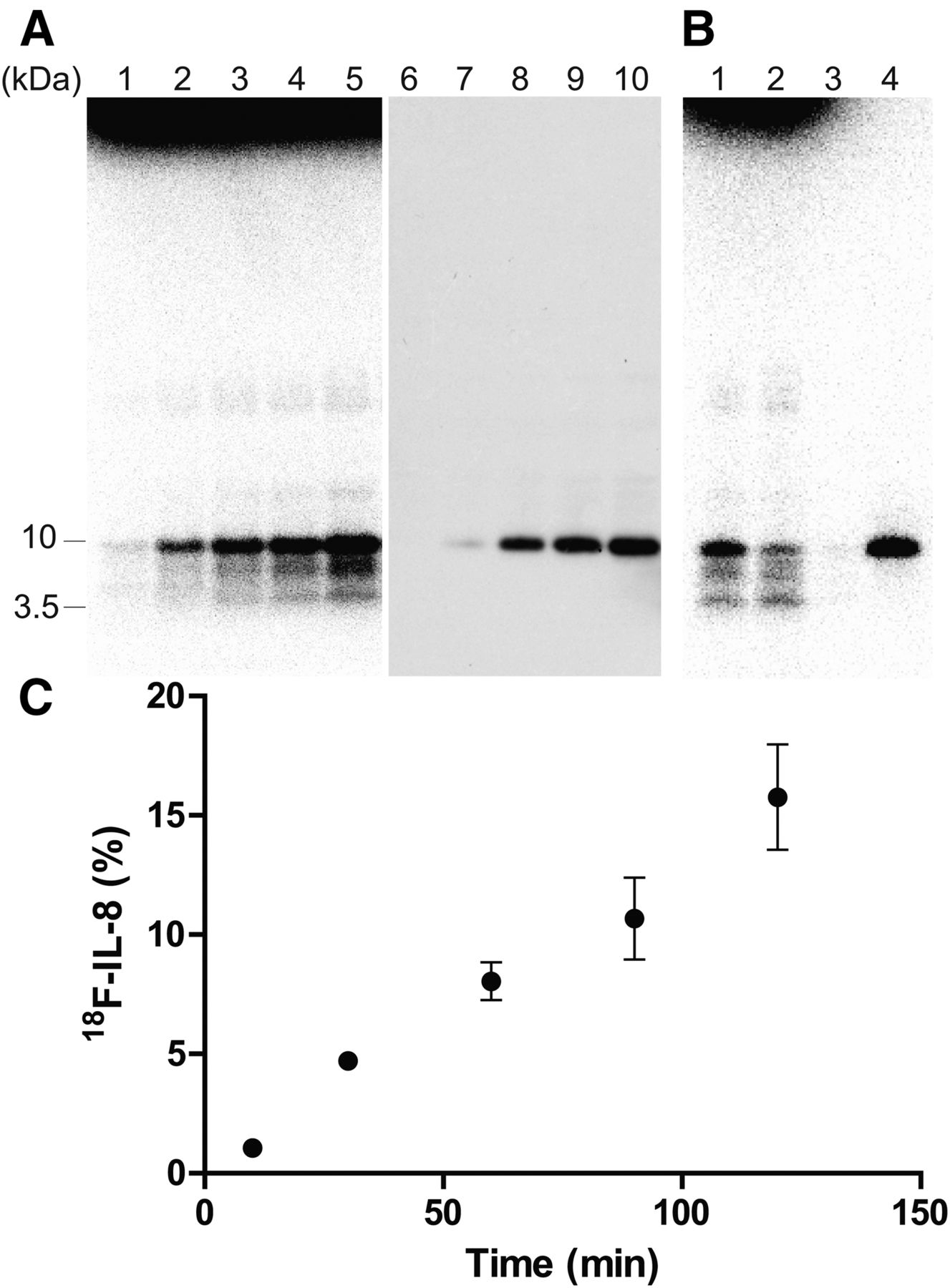
We subsequently determined the optimal reaction time for 18F-IL-8 synthesis. Although there were some radioactive bands derived from incomplete products of 18F-IL-8, a radioactive band at approximately 10 kDa was prominent, which is consistent with the results of Western blotting of IL-8 after radioactive decay (Fig. 3A). 18F-IL-8 was purified by a cation exchange spin column within 30 min like 11C-IL-8 (5), with resulting radiochemical purity exceeding 92% as determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis autoradiography (Fig. 3B). However, 18F-IL-8 production did not saturate within 120 min (Fig. 3C), indicating 18F-FPro may be slowly incorporated into the protein synthesis because it is a nonnatural amino acid. The decay-corrected radiochemical yield based on 18F-FPro was less than 1.5% at the total synthesis time of 150 min. The specific activity of 18F-IL-8 was estimated to 0.3 MBq/μg (2.6 GBq/μmol) at the end of synthesis by the preliminary analysis of IL-8 mass after radioactive decay, which was lower than conventional methods (17,18).

Radiosynthesis of 18F-IL-8 using PURExpress system and trans-4-18F-fluoro-l-proline. (A) Gel autoradiography and Western blot analysis. Lanes 1, 2, 3, 4, and 5: incubation for 15, 30, 60, 90, and 120 min, respectively. (B) Purification of 18F-IL-8 by cation exchange spin column. Lane 1: crude reaction solution. Lane 2: reaction solution passed through cation exchange spin column. Lane 3: washing solution with 0.1 M NaCl-HEPES (pH 7.6). Lane 4: eluted solution from spin column with 1 M NaCl-HEPES (pH 7.6). (C) Optimization of reaction time for 18F-IL-8 synthesis.
18F-IL-8 Binding to IL-8 RA
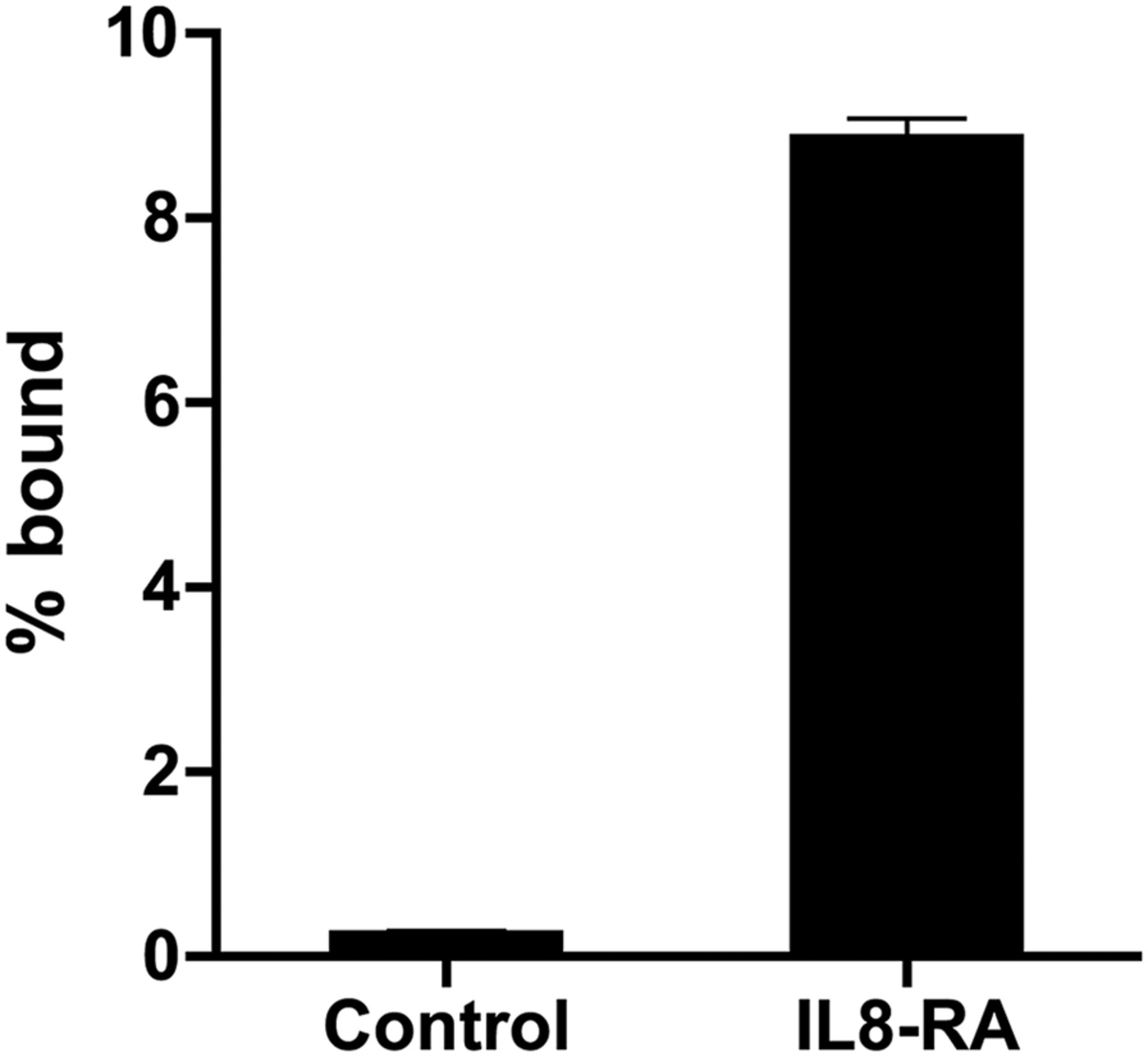
18F-IL-8 specifically bound to IL-8 RA–expressing CHO cells and not control CHO cells (Fig. 4).

In vitro binding of 18F-IL-8 to CHO cells transfected with pF5K CMV-neo containing control or IL-8 RA vector. Percentage bound (% bound) was obtained by taking radioactivity bound to cells divided by that added to wells.
IL-8 RA Imaging in Xenograft-Bearing Mice
In normal mice, 18F-IL-8 exhibited the largest tracer uptake in the kidneys and then gradually accumulated in the bladder (Fig. 5). No remarkable tracer uptake was observed during the PET scan except in the kidneys and bladder.
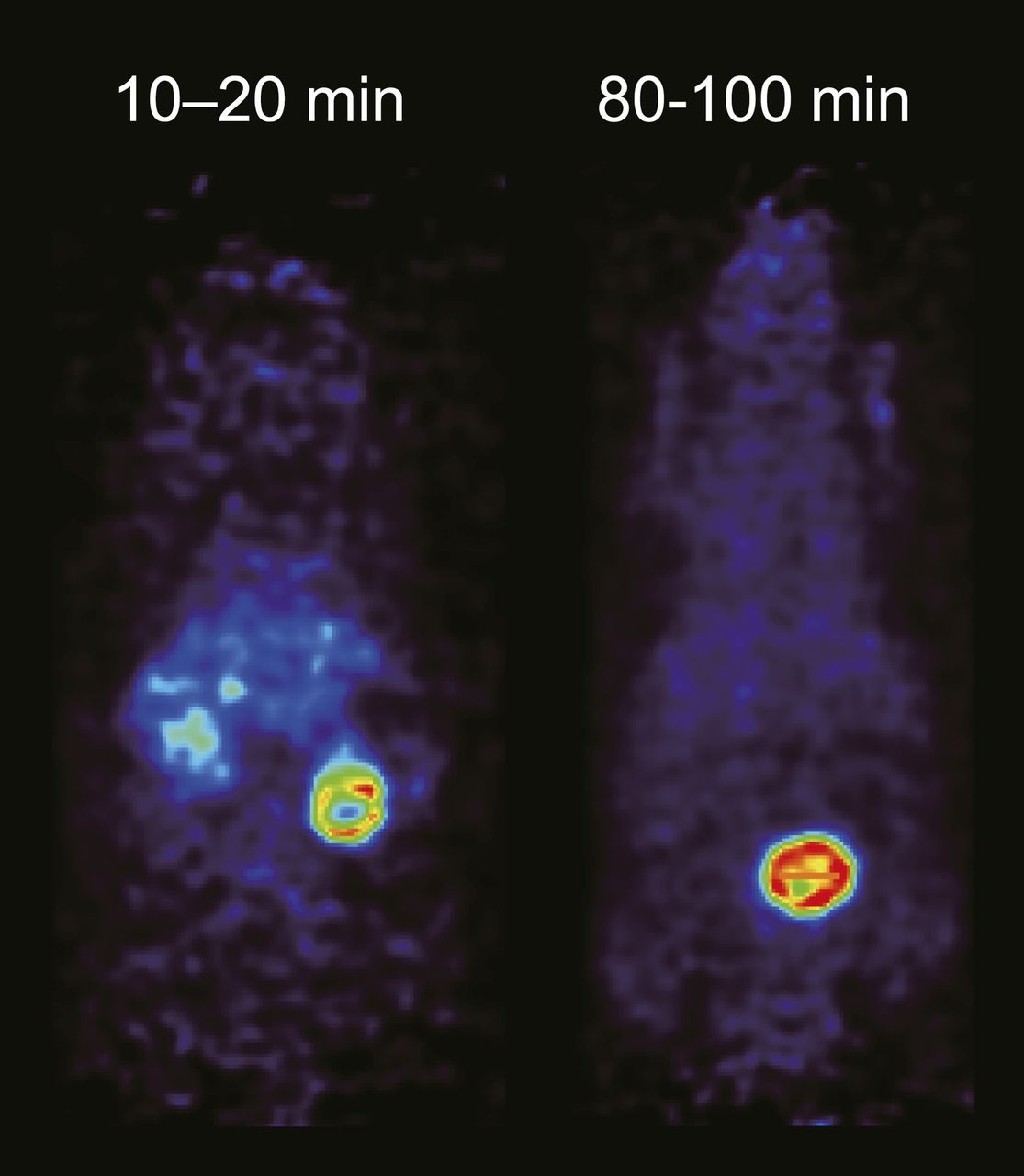
18F-IL-8 PET images from 10 to 20 min and 80 to 100 min in normal mice after intravenous administration of 18F-IL-8.
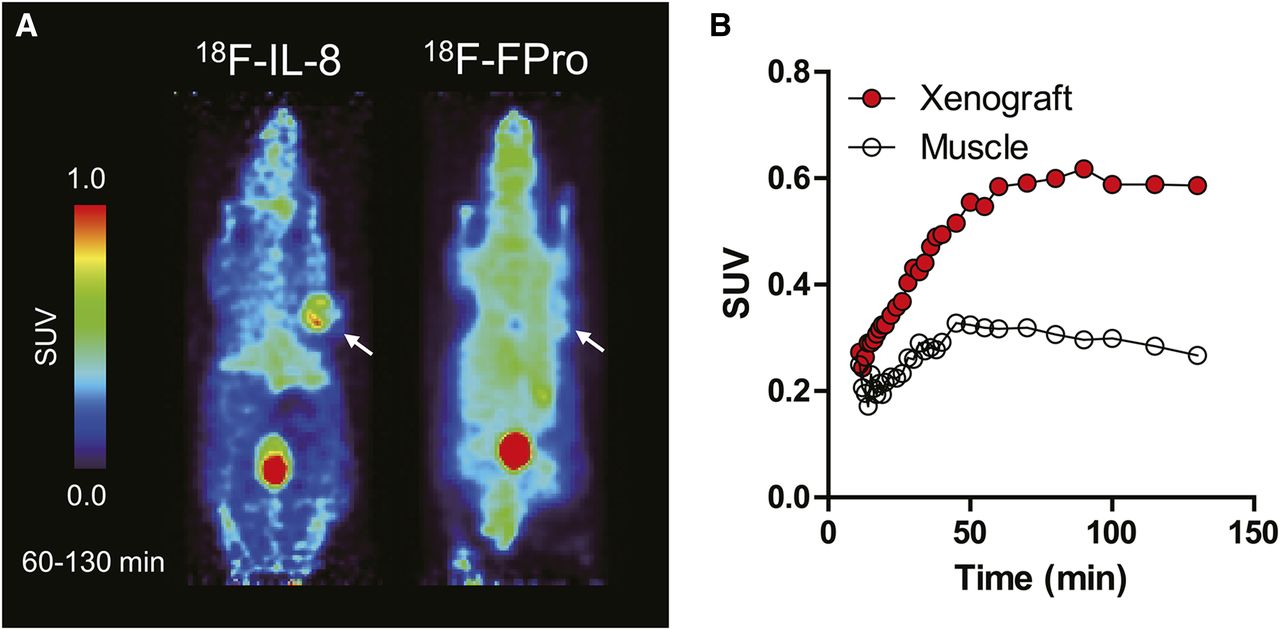
The PET images summed from 60 to 130 min after intravenous administration of 18F-IL-8 and trans-18F-FPro are shown in Figure 6A. 18F-IL-8 clearly showed tracer uptake in IL-8 RA xenografts with high contrast, whereas trans-18F-FPro showed a low signal-to-background ratio, making it difficult to distinguish IL-8 RA–expressing xenografts from other areas. The time–activity curves of 18F-IL-8 are shown in Figure 6B. 18F-IL-8 rapidly accumulated in the IL-8 RA–expressing xenografts, and tracer uptake plateaued at 60 min. The tracer uptake in the xenograft was approximately twice that in muscle at 60 min.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) 18F-IL-8 and trans-4-18F-fluoro-l-proline PET images from 60 to 130 min in IL-8 RA–expressing xenograft-bearing mice after intravenous administration of radiotracers. (B) Time–activity curves of 18F-IL-8 in IL-8 RA–expressing xenografts and muscle.
DISCUSSION
We previously reported a novel technique for labeling macromolecules such as peptides and proteins using a cell-free translation system and 11C-labeled l-methionine (5,6). However, macromolecules generally exhibit slower pharmacokinetics than small molecules, thus requiring a longer scanning time. To overcome these limitations, we expanded our strategy for PET-labeling macromolecules to use 18F, which has a longer half-life, consequently allowing imaging several hours after intravenous injection (17,19). In the present study, we successfully prepared 18F-IL-8, which showed high affinity for IL-8 RA as a model protein, synthesized by a cell-free translation system with trans-18F-FPro and clearly visualized IL-8 RA–expressing xenografts in vivo.
Because nonnatural amino acids are generally not recognized by aminoacyl-tRNA synthase due to its high substrate specificity, incorporation of nonnatural amino acids into proteins has been challenged using cell-free translation systems with additional various engineered molecules (8–12). Initially we unsuccessfully screened unlabeled fluorinated amino acids such as O-fluoromethyl-l-tyrosine, which have been used for radiolabeling (13,20), to be directly incorporated into proteins in a cell-free translation system derived from E. coli. instead of natural amino acids without any additional components. However, in the present study we demonstrated that both FPro isomers were successfully incorporated into the protein using the cell-free translation system instead of Pro (Fig. 2). FPro is a nonnatural amino acid used for protein folding research as well as a radiotracer for abnormal collagen synthesis and amino transport system A (21–23). Therefore, 18F-FPro is a good candidate radiolabeled amino acid source in a cell-free translation system. Pro residues affect protein folding and stability via the cis/trans isomerization of peptide bonds (24). The incorporation of FPro, which has 2 stereoisomers, into proteins may affect the binding affinity to targets. Interestingly, both FPro IL-8 variants exhibited higher affinity for IL-8 RA than the wild type (Fig. 2). Trans-FPro IL-8 possessed higher affinity than the cis-FPro variant in the case of IL-8 despite the lack of a substantial difference in the efficiency of incorporation into IL-8 protein between stereoisomers. As the effect of FPro incorporation on binding affinity appears to depend on proteins, cis-FPro may be an appropriate choice depending on the proteins even though trans-FPro is better than cis-FPro in this case.
Regarding the successful radiosynthesis of proteins using a cell-free translation system and 18F-FPro, there are some limitations. The first limitation is the lower radiochemical yield than conventional labeling methods for 18F (1). The decay-corrected radiochemical yields of trans-18F-FPro were high (30%–40%), but the maximum radiochemical yield of 18F-IL-8 was 1.5% (decay-corrected) based on 18F-FPro (0.45%–0.60% for decay-corrected radiochemical yield based on starting 18F-fluoride at the total synthesis time of 240 min). A possible reason for low radiochemical yield is considered to be simply unreacted 18F-FPro due to the slow reaction time. In addition, a large amount of radioactivity of 18F-FPro in the cell-free translation system showed a reduction in the radiochemical yield of 18F-IL-8 (data not shown), indicating another potential reason might be radioactive decomposition. Radical scavengers such as ascorbic acid might be a solution for low radiochemical yield even if they interfere with the reaction. A second limitation is the slow synthesis rate. As shown in Figure 4, 18F-IL-8 production did not plateau within 120 min (non–decay-corrected), which causes radioactive decay of the product and consequently low specific activity. 18F-FPro apparently has these limitations because it is not effectively recognized by aminoacyl-tRNA synthase and incorporated into proteins. Further studies are required to improve the synthesis rate and labeling efficiency. As another limitation of our method, incorporation sites of 18F-FPro depend on the sequences of interesting proteins. Nonnatural amino acids including fluorinated amino acids can be effectively and enzymatically incorporated into proteins by engineered orthogonal amber suppressor tRNA/aminoacyl-tRNA synthase pair in bacteria as well as cell-free translation systems (25–28). By combining other 18F-labeled amino acids for preparation of macromolecules labeled with 18F, allowing site-specific incorporation of nonnatural amino acid into the N terminus of interesting proteins, this technique might overcome these limitations.
IL-8 is a chemokine of approximately 80 amino acids (8.5 kDa) synthesized and secreted from several types of cells in response to inflammatory stimuli (29). Neutrophils can quickly respond to infection or injury and accumulate first in inflamed tissues. IL-8 binds to IL-8 receptors (i.e., IL-8 RA and IL-8 RB) expressed in neutrophils with high affinity (dissociation constant = 1 nM) (30). Therefore, positron-labeled IL-8 is a candidate PET tracer for inflammation imaging. As shown in Figure 5, 18F-IL-8 exhibited good pharmacokinetics—that is, fast clearance from normal tissues in mice via urinary excretion. IL-8 RA (chemokine [C-X-C motif] receptor 1) is highly selective for IL-8 in humans, but it is not expressed in mice or rats (31), making it difficult to evaluate the target engagement of 18F-IL-8 to IL-8 RA in vivo. Therefore, we used human IL-8 RA–expressing xenograft bearing–mice to determine whether 18F-IL-8 produced by the cell-free protein synthesis system works in vivo. 18F-IL-8 rapidly accumulated in IL-8 RA–expressing xenografts and exhibited a good signal-to-background ratio in vivo; in contrast, trans-18F-FPro exhibited low contrast as reported previously (32–34). Although full validation was required to characterize 18F-IL-8 by doing inhibition/blocking study for the development of 18F-IL-8 as a PET tracer, 18F-IL-8 produced by this technique was biologically active in vitro and in vivo.
CONCLUSION
The present study is proof of concept of our strategy for the preparation of proteins labeled with PET radionuclides—that is, 18F (which has a longer half-life)—using a cell-free translation system with trans-18F-FPro. This method can be performed with as little as the template DNA to prepare the desired 18F-labeled proteins. We successfully expanded our strategy from 11C to 18F, which is suitable for macromolecules with slow pharmacokinetics, requiring a longer scanning time. A protein radiotracer, 18F-IL-8, clearly visualized its target receptor–expressing xenograft in vivo. Therefore, this technique may be useful for labeling macromolecules and the preclinical evaluation of proteins of interest both in vitro and in vivo.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This study was supported by grants-in-aid for scientific research (23791387, 24790527, and 24659255) from the Japan Society of Promotion Science and the Ministry of Education, Culture, Sports, Science, and Technology, Japan. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank all of the staff at the Cyclotron Radioisotope Center of Tohoku University for the HM-12 cyclotron operation.
Footnotes
Published online Jan. 7, 2016.
- © 2016 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- Received for publication June 26, 2015.
- Accepted for publication December 1, 2015.