

Abstract
Although T cells can be labeled for noninvasive in vivo imaging, little is known about the impact of such labeling on T-cell function, and most imaging methods do not provide holistic information about trafficking kinetics, homing sites, or quantification. Methods: We developed protocols that minimize the inhibitory effects of 64Cu-pyruvaldehyde-bis(N4-methylthiosemicarbazone) (64Cu-PTSM) labeling on T-cell function and permit the homing patterns of T cells to be followed by PET. Thus, we labeled ovalbumin (OVA) T-cell receptor transgenic interferon (IFN)-γ–producing CD4+ T (Th1) cells with 0.7–2.2 MBq of 64Cu-PTSM and analyzed cell viability, IFN-γ production, proliferation, apoptosis, and DNA double-strand breaks and identified intracellular 64Cu accumulation sites by energy dispersive x-ray analysis. To elucidate the fate of Th1 cell homing by PET, 107 64Cu-OVA-Th1 cells were injected intraperitoneally or intravenously into healthy mice. To test the functional capacities of 64Cu-OVA-Th1 cells during experimental OVA-induced airway hyperreactivity, we injected 107 64Cu-OVA-Th1 cells intraperitoneally into OVA-immunized or nonimmunized healthy mice, which were challenged with OVA peptide or phosphate-buffered saline or remained untreated. In vivo PET investigations were followed by biodistribution, autoradiography, and fluorescence-activated cell sorting analysis. Results: PET revealed unexpected homing patterns depending on the mode of T-cell administration. Within 20 min after intraperitoneal administration, 64Cu-OVA-Th1 cells homed to the perithymic lymph nodes (LNs) of naive mice. Interestingly, intravenously administered 64Cu-OVA-Th1 cells homed predominantly into the lung and spleen but not into the perithymic LNs. The accumulation of 64Cu-OVA-Th1 cells in the pulmonary LNs (6.8 ± 1.1 percentage injected dose per cubic centimeter [%ID/cm3]) 24 h after injection was highest in the OVA-immunized and OVA-challenged OVA airway hyperreactivity–diseased littermates 24 h after intraperitoneal administration and lowest in the untreated littermates (3.7 ± 0.4 %ID/cm3). As expected, 64Cu-OVA-Th1 cells also accumulated significantly in the pulmonary LNs of nonimmunized OVA-challenged animals (6.1 ± 0.5 %ID/cm3) when compared with phosphate-buffered saline–challenged animals (4.6 ± 0.5 %ID/cm3). Conclusion: Our protocol permits the detection of Th1 cells in single LNs and enables temporal in vivo monitoring of T-cell homing over 48 h. This work enables future applications for 64Cu-PTSM–labeled T cells in clinical trials and novel therapy concepts focusing on T-cell–based immunotherapies of autoimmune diseases or cancer.
- 64Cu-PTSM
- murine Th1 cells
- small animal PET
- in vivo cell tracking
Noninvasive in vivo imaging of T-cell trafficking is of prime importance to gain further understanding of the pathogenesis of T-cell–mediated inflammation and to reveal the homing sites, the temporal distribution, and the mode of T-cell action in the context of novel immunotherapies. The use of imaging modalities to follow cell trafficking is also emerging in the fields of regenerative and individualized patient medicine, in which cells of interest must be labeled by appropriate markers to permit their detection in vivo, most likely by optical imaging, ultrasound, PET, or MR imaging. The individual imaging technologies have completely different labeling requirements, and each technique has strengths and weaknesses. For whole-body imaging of cell trafficking, the modalities with the highest potential are PET or MR imaging (1). PET provides exceptional sensitivity in the picomolar range and can reveal dynamic processes with a temporal resolution of several seconds (2). The high sensitivity and approximately 1-mm spatial resolution of the dedicated animal scanner make PET the most appropriate tool to track radiolabeled cells in vivo (3,4). In contrast to PET, MR imaging provides excellent anatomic information but has low sensitivity. For MR imaging, cells can be labeled with superparamagnetic iron oxide particles (SPIOs) that decrease the relaxation times and thus produce negative contrast (5). It is possible to determine the biodistribution of SPIOs-labeled cells in vivo by whole-body MR imaging. However, the sensitivity of MR imaging is approximately 106 times lower than that of PET, limiting its use for the detection of low cell numbers.
The radiolabeling of cells for PET imaging can be achieved by transducing the cells with a reporter gene, such as herpes simplex thymidine kinase, followed by the application of a protein-specific radiolabeled probe, by tagging a radioactive antibody to the cells or by in vitro intracellular labeling with a radiotracer (6–8). All of these methods have distinct benefits and disadvantages. Only a few studies have addressed radioactive T-cell labeling, because this cell population is more prone to be damaged by radiolabeled foreign particles and is more difficult to handle than, for example, cancer or stem cells (9).
Adonai et al. described 64Cu-pyruvaldehyde-bis(N4-methylthiosemicarbazone) (64Cu-PTSM) as an intracellular radiolabel for murine lymphocytes (8). The redox-active carrier molecule 64Cu-PTSM diffuses passively across the cell membrane because of its lipophilicity. Within the cytoplasm, Cu(II)-PTSM is reduced to the unstable Cu(I)-PTSM, resulting in the dissociation of Cu(I) from PTSM. The main step of the cell-labeling process is represented by trapping of the released 64Cu, which is subsequently bound by intracellular proteins (8,10). Because the positron-emitting radioisotope 64Cu has a half-life of 12.7 h, noninvasive cell tracking by PET is feasible for a maximum of 2–3 d. Surprisingly, no data exist concerning the impact of 64Cu-PTSM labeling on the function of lymphocytes.
Here, we evaluated the impact of this label on murine Th1 cells in vitro and in vivo, because Th1 cells enable the visualization of specific homing in the lymph nodes (LNs), the site of MHC-II antigen presentation. We studied the mechanistic properties of the 64Cu-labeling procedures and developed a 64Cu-PTSM–labeling protocol with minimized effects on T-cell survival and functions and permitting the monitoring of in vivo T cell trafficking for up to 48 h.
MATERIALS AND METHODS
64Cu-PTSM Ovalbumin (OVA)-Th1 Cell Labeling
To establish an optimized protocol for the 64Cu-PTSM labeling of mouse Th1 cells, 1 × 106 OVA-Th1 cells were incubated for 2, 3, or 4.5 h without or with 0.7, 1.4, or 2.2 MBq of 64Cu-PTSM in 0.25, 0.5, or 1.0 mL of cell culture medium in 24-well plates at 37°C and 7.5% CO2. For longitudinal in vivo studies, OVA-Th1 cells were always labeled with 0.7 MBq of 64Cu-PTSM for 3 h. To evaluate the impact of the metal copper and PTSM on the OVA-Th1 cells, we used 2 × 106 OVA-Th1 cells in 0.5 mL of cell culture medium (containing 5 U/mL IL-2) in 24-well plates at 37°C and 7.5% CO2. The cells were incubated with 0 μg, 60 pg, or 2 μg of nonradioactive Cu-PTSM or with 0, 0.15, 0.30, or 0.40 μg of PTSM for 2, 3, or 4.5 h. After the incubation, the cells were washed twice and prepared for in vitro and in vivo evaluations.
In Vivo PET Imaging
Specific OVA-Th1 cell trafficking and homing in mice with OVA-induced airway hyperreactivity (AHR) and untreated control mice were investigated after intraperitoneal (in 200 μL of phosphate-buffered saline [PBS]) or intravenous (in 150 μL of PBS) injection of 1 × 107 64Cu-OVA-Th1 cells, which corresponds to an activity of 0.2–0.5 MBq. The 64Cu-OVA-Th1 cells were injected immediately after the PET scan was started. The homing of OVA-Th1 cells in untreated BALB/c mice was assessed noninvasively in vivo by PET over 40 h. Dynamic PET scans were obtained over 180 min, and 20-min static PET scans were acquired 3, 7, 16, 27, and 40 h after Th1 cell injection (intravenous or intraperitoneal). Animals with OVA-induced AHR were imaged (static PET scans) 3, 24, and 48 h after Th1 cell injection (intraperitoneal). Mice were anesthetized 10 min before the PET scans with 1.5% isoflurane in 100% oxygen at a flow rate of 0.7 L/min using a dedicated rodent anesthesia system (Vetland). During the scans, the body temperature of the animals was kept constant at 37°C with a warming pad.
In vivo PET scans were acquired on a small-animal Inveon microPET scanner (Siemens). A 2-dimensional iterative ordered-subset expectation maximization algorithm was used to reconstruct the list mode data. The spatial resolution of the reconstructed PET images was approximately 1.4 mm (3). CT scans were obtained on an Inveon dedicated CT scanner (Siemens). The x-ray voltage was set at 70 keVp, and the current was set to 500 μA. An exposure time of 350 ms and a binning factor of 4 were chosen to yield a reconstructed pixel size of 75 μm. Planar images were acquired from 360 projections over a 360° total rotation in a step-and-shoot mode and reconstructed to obtain a 3-dimensional image. To enable the coregistration of the PET and CT images, we filled glass capillaries with 64Cu-PTSM and fixed them underneath the mouse bed for use in both scanners.
More methods are provided in the supplemental materials (available at http://jnm.snmjournals.org).
RESULTS
Specific T-Cell Function After 64Cu-PTSM Labeling
64Cu-PTSM–radiolabeling protocols were optimized to minimize the impact on Th1 cell function (Fig. 1). OVA-Th1 cells were incubated with 0.7, 1.5, or 2.2 MBq for 2, 3, or 4.5 h, respectively. The radiolabel uptake efficiency and labeling stability were investigated to yield an optimized procedure that consisted of a 3-h incubation of 106 OVA-Th1 cells with 0.7 MBq of 64Cu-PTSM in 500 μL of medium (Supplemental Fig. 1A). This labeling protocol permits an acceptable labeling efficiency of 6.5%–9% (Supplemental Figs. 1A and 1B). Efflux studies revealed that 47% of 64Cu-PTSM remained in the OVA-Th1 cells after 5 h and 14% after 24 h, respectively (Supplemental Fig. 1B).
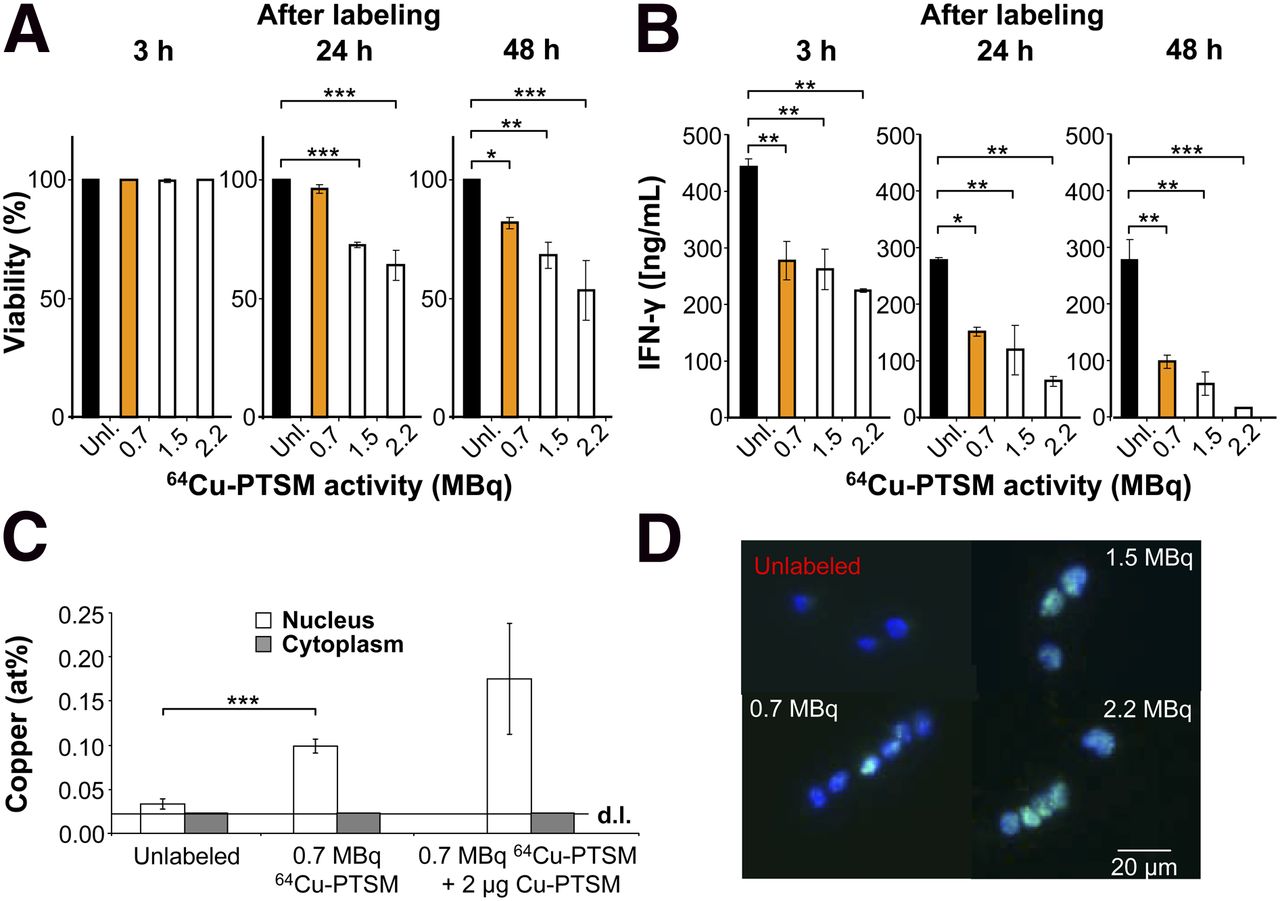
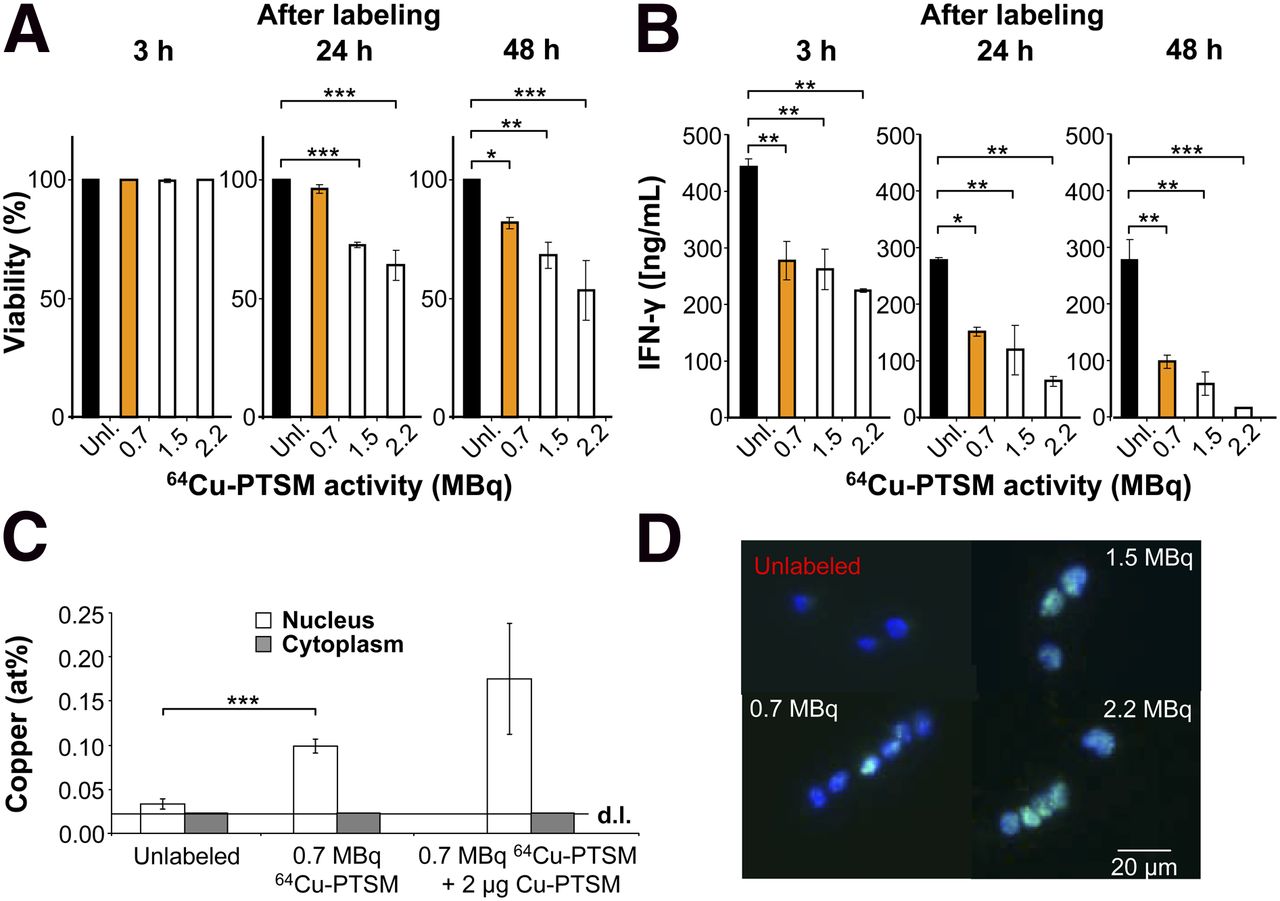
Impact of 64Cu-PTSM labeling on viability, T-cell function, and double-strand breaks. (A) Viability after labeling with 64Cu-PTSM; Dunnet’s test. *P < 0.05. **P < 0.01. ***P < 0.001. Mean ± SD; n = 3; 5 independent experiments. (B) IFN-γ concentration (ng/mL) in supernatants of OVA-Th1 cells that were cultured for 24 h with irradiated antigen-presenting cells and OVA peptide after labeling with 64Cu-PTSM; Dunnet’s test. *P < 0.05. **P < 0.01. ***P < 0.001. Mean ± SD; n = 2; 3 independent experiments. (C) Mole fraction of copper (mean at percentage ± SD) after EDX analysis within nucleus and cytoplasm of OVA-Th1 cells that were unlabeled, labeled with 0.7 MBq of 64Cu-PTSM, or with 0.7 MBq of 64Cu-PTSM and 2 μg of Cu-PTSM. Copper mole fraction in cytoplasm was close to detection limit (d.l.) of 0.02 at percentage; 2-sampled Student t test. ***P < 0.001. (D) Fluorescence microscopy indicates phosphorylated γ-H2AX in OVA-Th1 cells 3 h after labeling with 64Cu-PTSM (n = 2–3) in T-cell nucleus. Green = γ-H2AX antibody; blue = 4′, 6-diamidin-2-phenylindol for nuclear staining.
Following this labeling protocol, trypan blue exclusion demonstrated the unaltered viability of Th1 cells 3 h after labeling with 0.7 MBq of 64Cu-PTSM and only a slight reduction in viability after 24 and 48 h, as compared with the unlabeled control cells (Fig. 1A). Similarly, controls with high concentrations of nonradioactive Cu-PTSM did not show impaired OVA-Th1 cell viability (Supplemental Fig. 1C). A significantly reduced viability was observed when cells were incubated with 1.5 or 2.2 MBq of 64Cu-PTSM (Fig. 1A). Thus, 3 h of labeling of OVA-Th1 cells with 0.7 MBq of 64Cu-PTSM appears to be optimal with respect to viability and labeling efficiency.
With regard to specific Th1 cell functions, we observed reduced interferon (IFN)-γ concentrations in the supernatants of OVA-Th1 cells labeled with 0.7 MBq of 64Cu-PTSM at 3, 24, and 48 h after specific stimulation with antigen-presenting cells and OVA peptide when compared with unlabeled OVA-Th1 cells (Fig. 1B). Increased incubation times further reduced the amount of IFN-γ in the supernatants (Supplemental Fig. 2A). PTSM alone did not impair the OVA-specific IFN-γ secretion of OVA-Th1 cells (Supplemental Fig. 2B), whereas high concentrations of nonradioactive Cu-PTSM strongly suppressed IFN-γ secretion (Supplemental Fig. 2C).
The OVA-specific proliferation rate of 0.7 MBq of 64Cu-PTSM–labeled OVA-Th1 cells, as measured using a 3H-thymidine proliferation assay, indicated a 69% reduction in proliferation 3 h after labeling. Interestingly, incubation with a 70-fold-higher concentration of nonradioactive copper reduced OVA-specific Th1 cell proliferation by 35%, which indicates that the metal itself impaired the proliferation (Supplemental Fig. 2D). However, real-time polymerase chain reaction investigations revealed 3 h after labeling normal IFN-γ messenger RNA levels and 24 h after labeling enhanced IFN-γ messenger RNA levels in 64Cu-OVA-Th1 cells (Supplemental Fig. 3).
Intracellular Localization of 64Cu and DNA Damage
To determine how 64Cu labeling affects OVA-Th1 cell function, we investigated the intracellular concentration and localization of copper in nonlabeled cells and in cells labeled with 0.7 MBq of 64Cu-PTSM or with 0.7 MBq of 64Cu-PTSM plus a 70-fold-higher concentration of nonradioactive Cu-PTSM using electron microscopy and quantitative energy dispersive x-ray (EDX) spectroscopy (Supplemental Fig. 4) (11,12).
Surprisingly, 64Cu was mainly trapped in the cell nucleus, because the copper mole fraction in the nuclei (0.1 ± 0.01 at %) of OVA-Th1 cells exposed to 0.7 MBq of 64Cu-PTSM was higher than that in the cytoplasm. The copper mole fraction of the cytoplasm was close to the detection limit of 0.02%. The copper mole fraction in the nuclei of OVA-Th1 cells labeled with 0.7 MBq of 64Cu-PTSM was 3.3-fold higher than that in the nuclei of unlabeled cells. Importantly, the investigated cells provided similar 64Cu concentrations (Fig. 1C).
We assessed phosphorylated histones of the H2AX family (γ-H2AX) by fluorescence microscopy and fluorescence-activated cell sorting (FACS) to investigate whether 64Cu accumulation within the nuclei leads to radiation-induced DNA double-strand breaks. Fluorescence microscopy analysis of γ-H2AX expression demonstrated double-strand breaks in the nuclei of 64Cu-PTSM–labeled OVA-Th1 cells (Fig. 1D). The relative γ-H2AX expression 3 h after labeling with 0.7 MBq of 64Cu-PTSM was approximately 9-fold higher than that of unlabeled OVA-Th1 cells (Supplemental Fig. 5; Supplemental Table 1).
In Vivo Homing Patterns After Different Administration Routes of 64Cu-OVA-Th1 Cells
The homing of 64Cu-PTSM–labeled OVA-Th1 cells was tracked in vivo by PET and fused with CT images to obtain additional anatomic information. We first analyzed the differences in T-cell homing kinetics and biodistribution after intraperitoneal and intravenous administration of 64Cu-PTSM–labeled OVA-Th1 cells (Fig. 2). Naïve mice were injected intraperitoneally or intravenously with 107 viable 64Cu-OVA-Th1 cells, and 180-min dynamic PET scans were obtained immediately after administration. In addition, 20-min static PET data were acquired 3, 7, 16, 27, and 40 h after 64Cu-OVA-Th1 cell administration (Fig. 2A).

Different administration routes of 64Cu-PTSM–labeled OVA-Th1 cells resulted in distinct homing patterns. (A) Time–activity curves (%ID/cm3) derived from perithymic LNs, lung, and spleen after intraperitoneal and intravenous transfer of 107 64Cu-PTSM–labeled OVA-Th1 cells display dynamics of OVA-Th1 cell homing in vivo within first 180 min (left; dynamic PET measurement); mean ± SEM (n = 3). Differences in homing patterns of 64Cu-OVA-Th1 cells (%ID/cm3) in perithymic LNs, lung, and spleen at 3, 7, 16, 27, and 40 h after intraperitoneal or intravenous administration (static 20-min PET measurements) are shown on right; 2-sampled Student t test. *P < 0.05. **P < 0.01. Mean ± SEM (n = 6). (B) Representative PET/CT images at 7 h (left) and 27 h (right), with focus on perithymic LNs, lung, and spleen (indicated by arrows). i.p. = intraperitoneal; i.v. = intravenous.
Interestingly, intraperitoneally injected OVA-Th1 cells homed to the perithymic LNs within the first 20 min after administration (Figs. 2A and 2B). Given that 107 viable 64Cu-OVA-Th1 cells were injected and neglecting the 64Cu efflux from the OVA-Th1 cells (Supplemental Fig. 1B), it was possible to detect approximately 60,000 64Cu-OVA-Th1 cells in 1 individual LN 20 min after intraperitoneal administration (0.33 ± 0.02 percentage injected dose per cubic centimeter [%ID/cm3]). The activity of 64Cu-OVA-Th1 cells in the perithymic LNs remained constant between 2.0 ± 0.5 %ID/cm3 at 7 h and 2.9 ± 0.5 %ID/cm3 at 40 h after intraperitoneal T-cell transfer. It was not possible to detect specific signals related to 64Cu-OVA-Th1 cells in the perithymic LNs after intravenous administration because the analysis was impaired by the transient adherence of a high number of 64Cu-OVA-Th1 cells in the lung, which produced a high background signal.
Thus, in line with the observations in other cell-trafficking studies (8,13,14), many 64Cu-OVA-Th1 cells nonspecifically accumulated in the lung tissue immediately after intravenous administration (57.4 ± 1.3 %ID/cm3). The signal decreased at a constant rate within the first 180 min to 16.3 ± 0.8 %ID/cm3 and then further to 5.2 ± 0.7 %ID/cm3 at 40 h. In sharp contrast to the intravenous injection, the lung tissue exhibited only background levels of Th1 cell accumulation after intraperitoneal injection (Figs. 2A and 2B).
Within 5 min of intraperitoneal injection, 64Cu-OVA-Th1 cells accumulated rapidly in the spleen (21.1 ± 6.0 %ID/cm3) and decreased to 14.8 ± 2.1 %ID/cm3 by the end of the 180-min dynamic PET scan. Static PET scans revealed values of 6.7 ± 1.8 %ID/cm3 in the spleen at 3 h and 3.2 ± 0.7 %ID/cm3 at 40 h (Figs. 2A and 2B). The increased values in the spleen in the dynamic PET measurement could be the consequence of an increased spillover caused by the intraperitoneally injected 64Cu-OVA-Th1 cells in the peritoneum, intestine, and peripancreatic lymphatic tissue. Mice that underwent dynamic PET scans were anesthetized with 1.5% isoflurane for 3 h, which can cause a reduced intestine movement and motility, yielding a higher accumulation of the 64Cu-OVA-Th1 cells within the intestines (Supplemental Fig. 6) (15). In contrast, animals that were used for the static PET scans were conscious during the first 3 h. Isoflurane anesthesia evolves well-described antiinflammatory effects applicable to suppress cell migration, further explaining why more of the intraperitoneally injected 64Cu-OVA-Th1 cells remained in the peritoneal cavity (16).
In contrast, intravenous injection produced a constant increase in the activity accumulation of 64Cu-OVA-Th1 cells to 19.4 ± 4.0 %ID/cm3 during the 180-min dynamic PET scan. Thereafter, activity of 64Cu-OVA-Th1 cells decreased in the spleen until 40 h after intravenous injection to 11.5 ± 1.7 %ID/cm3. Thus, 16–40 h after intravenous administration approximately 3.5-fold more 64Cu-OVA-Th1 cells homed to the spleen when compared with intraperitoneally injected 64Cu-OVA-Th1 cells.
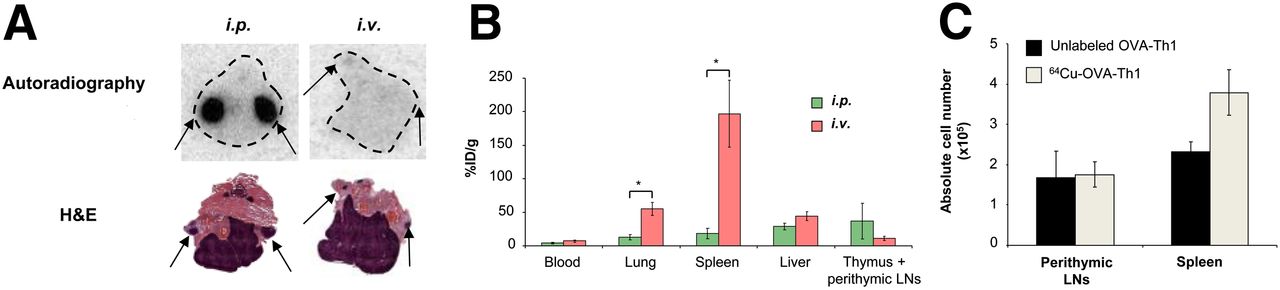
Because PET measurements of the intravenously administered 64Cu-OVA-Th1 cells did not permit the investigation of cell homing to the perithymic LNs, animals were sacrificed 24 h after the intraperitoneal or intravenous administration of 64Cu-OVA-Th1 cells, the thymus with perithymic LNs was dissected, and autoradiography and hematoxylin and eosin staining of the corresponding tissue sections was performed (Fig. 3). The perithymic LNs were confirmed to be the site of 64Cu-OVA-Th1 cell accumulation 24 h after intraperitoneal injection; almost no OVA-Th1 cells homed to the perithymic LNs after intravenous administration (Fig. 3A). Additionally ex vivo biodistribution was performed by γ counting of dissected tissue 40 h after intraperitoneal or intravenous administration of 64Cu-OVA-Th1 cells (Fig. 3B).
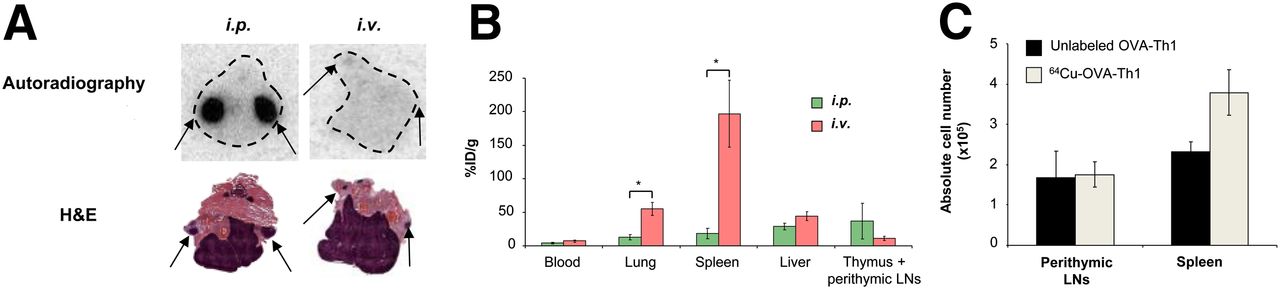
64Cu-PTSM labeling does not impair in vivo OVA-Th1 cell homing. (A) Representative hematoxylin and eosin (H&E)–stained sections and autoradiography images of thymus and perithymic LNs 24 h after intravenous or intraperitoneal 64Cu-OVA-Th1 cell transfer. Autoradiography images revealed perithymic LNs (arrows) to be one of the early homing sites of 64Cu-OVA-Th1 cells after intraperitoneal transfer but not after intravenous administration (n = 3). (B) Biodistribution analysis (%ID/g) of blood, lung, spleen, liver, and thymus (including perithymic LNs) was performed 40 h after intraperitoneal or intravenous 64Cu-OVA-Th1 cell transfer to confirm in vivo PET imaging results; 2-sampled Student t test. *P < 0.05. Mean ± SEM (n = 6). (C) FACS analysis of intraperitoneally administered nonlabeled or 64Cu-PTSM–labeled OVA-Th1 cells revealed almost equivalent numbers of nonlabeled and 64Cu-OVA-Th1 cells within perithymic LNs and increased number of 64Cu-OVA-Th1 cells in spleen. Mean ± SEM (n = 4). i.p. = intraperitoneal; i.v. = intravenous.
To further analyze the homing of intraperitoneally administered OVA-Th1 cells to the perithymic LNs and spleen and the impact of the 64Cu radiolabel on OVA-Th1 cell homing, we subsequently intraperitoneally injected 107 nonlabeled or 64Cu-labeled OVA-Th1 cells into BALB/c mice and performed in vivo PET measurements. The animals were sacrificed after 21 h, the perithymic LNs and spleens were dissected, the lymphocytes were quantified, and FACS analysis was performed to quantify the adoptively transferred OVA-Th1 cells using T-cell receptor–specific antibodies (Fig. 3C; Supplemental Fig. 7). As expected, even without antigen application, intraperitoneally administered OVA-Th1 cells were clearly detected in the perithymic LNs and the spleen. The number of OVA-Th1 cells in the LNs corresponded to previous data published by Schipf et al. (17). No significant differences in the absolute numbers of nonlabeled (1.6 × 106 ± 6.7 × 105) and 64Cu-OVA-Th1 cells (1.8 × 106 ± 3.1 × 105) were found in the perithymic LNs. Slight increased numbers of 64Cu-OVA-Th1 cells (3.8 × 106 ± 5.6 × 105), compared with nonlabeled cells (2.3 × 106 ± 2.4 × 105; Fig. 3C), were detected in the spleen. Thus, 64Cu-PTSM labeling did not affect OVA-Th1 cell–homing properties in vivo.
Specific Homing of 64Cu-OVA-Th1 Cells in OVA-Induced AHR
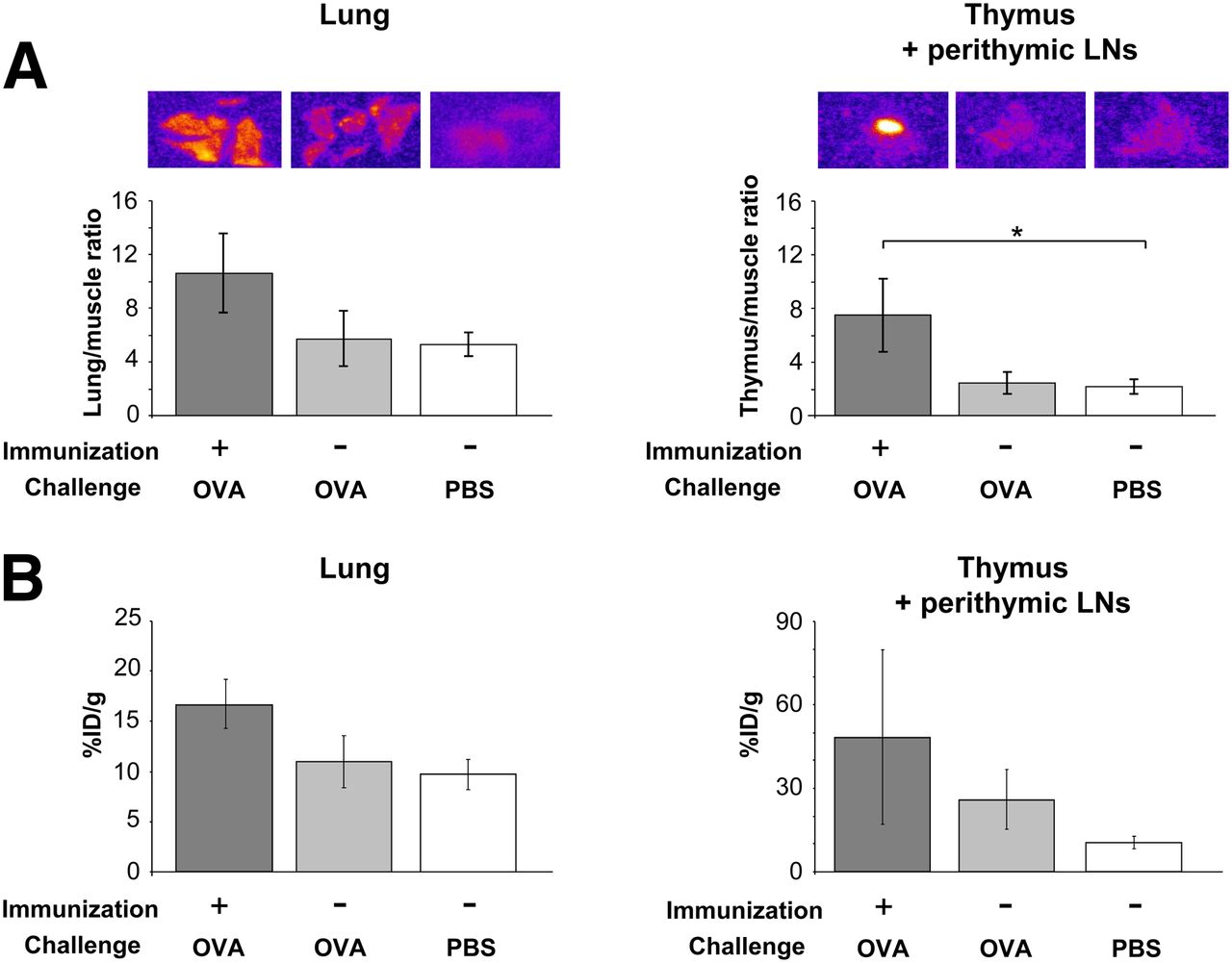
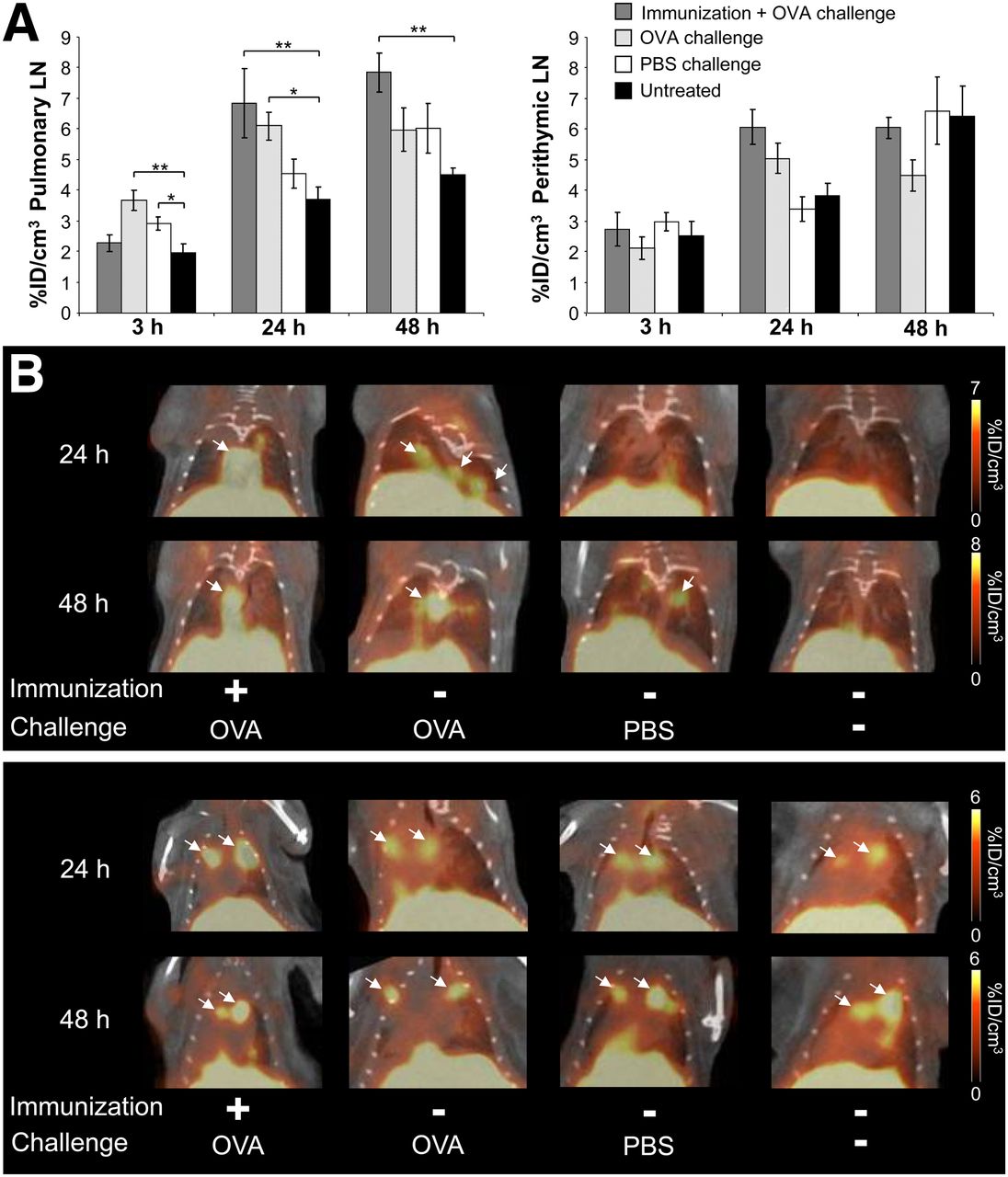
To investigate the functional capacities of 64Cu-OVA-Th1 cells, we tested their ability to home to OVA-specific sites of inflammation using an in vivo model of experimental OVA-induced AHR of the lung (Fig. 4). To determine specific homing, we injected 107 64Cu-OVA-Th1 cells intraperitoneally into OVA-immunized or nonimmunized healthy mice. The mice were challenged twice intranasally with OVA peptide or PBS or they remained untreated. The activity accumulation of 64Cu-OVA-Th1 cells in the pulmonary LNs (6.8 ± 1.1 %ID/cm3) 24 h after injection was highest in the OVA-immunized and OVA-challenged animals and lowest in the untreated littermates (3.7 ± 0.4 %ID/cm3; Fig. 4A, left; Fig. 4B, upper). As expected, 64Cu-OVA-Th1 cells also accumulated significantly in the pulmonary LNs of nonimmunized OVA-challenged animals (6.1 ± 0.5 %ID/cm3), whereas in PBS-challenged animals, the activity values (4.6 ± 0.5 %ID/cm3) 24 h after T-cell transfer were similar to the background of untreated mice. The OVA-specific homing of 64Cu-OVA-Th1 cells in nonprimed but OVA-challenged mice was expected, because the peptide is absorbed and presented in the draining LNs. Specific accumulation in the pulmonary LNs persisted for 48 h only in the OVA-challenged mice that were immunized. These data suggest that the primed mice develop sustained inflammation in the lung. As observed in the lung and the pulmonary LNs, we detected a tendency toward increased OVA-specific accumulation of 64Cu-OVA-Th1 cells in the perithymic LNs of OVA-immunized and OVA-challenged mice 24 h after T-cell administration (Fig. 4A, right; Fig. 4B, lower), a point in time with most severe lung inflammation. Although the perithymic LNs are not the draining LNs of the lungs, they may be affected by the lung inflammation.

64Cu-PTSM–labeled OVA-Th1 cells home specifically in draining lymphatic tissue at sites of OVA-induced AHR. (A) PET/CT measurements of activity accumulation of 64Cu-PTSM–labeled OVA-Th1 cells (%ID/cm3) in pulmonary LNs (left) and perithymic LNs (right) after intraperitoneal transfer of 107 64Cu-OVA-Th1 cells into diseased and control mice. Enhanced homing of OVA-Th1 cells in OVA-immunized and OVA-challenged mice to pulmonary LNs; Dunnet’s test. *P < 0.05. **P < 0.01. Untreated, n = 6; OVA-immunized + OVA-challenged, n = 7–19; OVA-challenged, n = 7–18; and PBS-challenged mice, n = 4–12. Mean ± SEM. (B) Representative PET/CT images 24 and 48 h after intraperitoneal administration of 64Cu-OVA-Th1 cells focusing on pulmonary LNs (upper) and perithymic LNs (lower; arrows).
To exclude an unspecific signal caused by the 64Cu efflux from 64Cu-OVA-Th1 cells, we intraperitoneally injected 0.7 MBq of 64Cu-PTSM in OVA-immunized and OVA-challenged mice or in healthy PBS-challenged control mice. We could not identify unspecific 64Cu-PTSM accumulation in both experimental groups in the lung tissue, pulmonary, and perithymic LNs 24 and 48 h after injection (Fig. 5). The PET/CT measurements agreed with the autoradiography and biodistribution in the lung tissue and the thymus (Fig. 6; supplemental data).

Distribution of 64Cu-PTSM in lung and perithymic LNs after intraperitoneal injection into OVA AHR–diseased and control mice. Representative PET/CT images 24 and 48 h after intraperitoneal administration of 0.7 MBq of 64Cu-PTSM indicate no accumulation of 64Cu-PTSM in lung tissue, pulmonary (left), and perityhmic LNs (right). High-resolution template CT scan provided detailed anatomic information for accurate localization of 64Cu-PTSM. Scans were normalized to injected activity of 107 64Cu-OVA-Th1 cells in OVA AHR–diseased animals (Fig. 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
64Cu-PTSM–labeled OVA-Th1 cells home specifically to lung and perithymic LNs of OVA AHR–diseased mice. (A) Autoradiography revealed 24 h after intraperitoneal 64Cu-OVA-Th1 cell administration increased lung-to-muscle and thymus-to-muscle ratios in OVA-immunized and OVA-challenged animals (n = 3), compared with ratios in OVA-challenged (n = 5) or PBS-challenged animals (n = 6). Dunnet’s test. *P < 0.05; mean ± SEM. (B) Biodistribution, performed 48 h after intraperitoneal 64Cu-OVA-Th1 cell injection, showed higher percentage injected dose per gram (%ID/g) in lung and thymus including perithymic LNs in OVA-immunized and OVA-challenged animals (n = 7), compared with OVA- (n = 6) or PBS-challenged animals (n = 5); mean ± SEM.
DISCUSSION
T-cell–based immunotherapy of cancer is an emerging novel therapeutic approach (18,19). Some data suggest that the adoptive transfer of autologous tumor-infiltrating lymphocytes may induce cancer regression in up to 50% of metastatic melanomas (20). Thus, tumor-associated antigen (TAA)–specific IFN-γ–producing CD8+ (Tc1) and CD4+ T cells (Th1) display strong antitumor effects in humans and mice (19,21–23).
To gain deeper insights into the early homing patterns, to quantify T-cell accumulation at the homing sites, and to identify the best mode of TAA T-cell administration, we applied 64Cu-PTSM labeling and highly sensitive noninvasive in vivo PET imaging. 64Cu-PTSM labeling of T cells may be superior to other labeling strategies, and the transfer of this strategy from rodents to humans may be safe because no viral-based manipulation with reporter genes is required. Alternatively, Matsui et al. investigated a labeling strategy based on T-cell–specific radiolabeled antibodies, which were used for the in vivo labeling of the TAA T cells (6). Their data raise several concerns, because nonbound or nonspecifically bound radiolabeled antibodies produce a severe background signal (6). Furthermore, the antibody can cause immune reactions, and most importantly, specific antibody binding to T cells may depend on the delivery efficacy of the antibody in vivo to the sites of T-cell accumulation (6,24). Thus, the spatial and temporal T-cell detection and quantification would depend on the bioavailability and the pharmacokinetics of the antibody (24,25). Similar limitations apply for transduced cells that use a reporter probe (9).
Several research groups routinely use 111In-oxine, which is a SPECT tracer, for immune cell labeling, to reveal non-identified sites of inflammation in patients or to study cell trafficking in rodents (26,27). However, we detected with 0.7 MBq of 111In-oxine–labeled murine OVA-Th1 cells an induction of double-strand breaks (unpublished results), which even was more severe than with 64Cu. Furthermore, the detection sensitivity of SPECT is 100- to 1,000-fold lower than that of PET (13,28).
As an alternative to nuclear methods, MR imaging has been established preclinically and clinically. Several groups are focusing on T-cell–trafficking studies using SPIOs or 19F-labeled cells and 19F MR imaging (5,29–33). T cells labeled with SPIOs provide a negative contrast signal in MR images, from which it is challenging to extract information about the whole-body biodistribution of the labeled immune cells (5,30,31). The detection of individual homing sites without a priori knowledge and absolute quantification are impossible. Therefore, Thu et al. injected SPIO-labeled bone marrow stromal cells intracranially at a defined detection site for MR imaging (5). This is in sharp contrast to our approach, in which we injected the radiolabeled cells systemically and followed up and quantified their whole-body biodistribution in vivo. The 19F MR imaging approach is interesting but suffers from the low detection sensitivity of MR imaging, resulting in a measurement time of several hours even for larger cell clusters of tens of thousands of cells and a limited field of view (29,32,33).
Because direct radioactive labeling approaches for cells of interest are limited by the radionuclide half-life, the induction of double-strand breaks, and the dilution of the radiolabel due to cell proliferation, indirect labeling approaches using reporter genes are an alternative labeling strategy for long-term cell-tracking studies (34). We transduced OVA-Th1 cells with the triple-fusion cassette containing the herpes simplex thymidine kinase as PET reporter gene (7) but achieved only an insufficient transduction efficiency (unpublished results). In addition, the application of virally modified cells in humans raises ethical concerns (4).
Our study provides the first, to our knowledge, comprehensive evaluation of 64Cu-PTSM labeling of murine antigen-specific Th1 cells and the intracellular 64Cu accumulation and impact on viability, function, and double-strand breaks.
Thus far, only a few research groups have focused on 64Cu-PTSM labeling of cells for cell-tracking studies, and only 2 groups investigated the cellular 64Cu efflux of 64Cu-PTSM–labeled cells (8,14). In our studies, we characterized and quantified the efflux of 64Cu from murine OVA-Th1 cells in vitro for the first time in detail and detected in in vitro experiments an efflux of 53% within 5 h and 86% within 24 h after labeling with 0.7 MBq of 64Cu-PTSM. These in vitro experiments reflect, compared with in vivo conditions, a worst case because the cells are densely packed and thus exposed to extended radiation from neighboring cells and activity in the medium. Adonai et al. performed efflux studies with C6 rat glioma cells and detected an efflux of 62% within 5 h and 78% within 24 h after the labeling procedure (8). Prince et al. claimed an efflux of 20% from human dendritic cells within the first hour after 64Cu-PTSM labeling but did not provide detailed efflux studies nor report the efflux 24 h after labeling (14). Thus, no direct comparison to our 64Cu efflux studies is possible because of the differences in cell type.
Th1 cells labeled with 0.7 MBq of 64Cu displayed only a slight reduction in viability, IFN-γ secretion, and cell proliferation and an induction of phosphorylated γ-H2AX histones. In contrast to Adonai et al., who proposed the hypothesis that 64Cu is mainly retained in the cytoplasm, we also found 64Cu in the cell nucleus by EDX analysis. The double-strand breaks might be caused by the localized radiation within the nucleus because 64Cu decays in 18% by positron emission (β+) and in 40% by electron capture, emitting Auger electrons that are radiotoxic to DNA (35,36).
Most importantly, despite the impact of the radiolabel on OVA-Th1 cells, we detected almost equivalent numbers of intraperitoneally injected 64Cu-OVA-Th1 cells and nonlabeled OVA-Th1 cells in the perithymic LNs and spleens of littermates by FACS. This finding supports our hypothesis that the radiolabeling of Th1 cells causes only minor cell damage in vivo. In our in vitro conditions, the effect on viability and function was most likely overestimated and represents the worst-case scenario regarding radiolabeling/cell concentrations. Using 64Cu-PTSM labeling of OVA-Th1 cells, we were able to identify clear differences between the intraperitoneal and intravenous administration of Th1 cells and were able to detect OVA-Th1 cells within a single LN for up to 40 h. Moreover, we could follow specific OVA-Th1 cell-trafficking to the site of inflammation in an experimental model of OVA-induced lung inflammation for up to 48 h. No 64Cu accumulation in the pulmonary and perithymic LNs could be observed in mice with OVA AHR after intraperitoneal injection of free 64Cu-PTSM (Fig. 5). Thus, 64Cu-PTSM labeling and PET imaging of autologous leukocytes is an appropriate approach to track immune cells in vivo and permits the study of the mechanistic properties of cell homing and function. The temporal monitoring also revealed the important finding that all trafficking processes occur quickly, most within several minutes or hours, suggesting that a fast imaging modality, such as PET, is beneficial. Interestingly, we also found that Th1 cells accumulated in the perithymic LNs, which was described so far in only one other report and leaves the underlying mechanism open to question (37). In contrast to Klinkert, we observed accumulation in perithymic LNs after intraperitoneal but not after intravenous injection, and PET imaging revealed that most of the Th1 cells had already reached the perithymic LNs 2 h after injection. Because the role of perithymic LNs remains unclear, further studies using different cell populations and stimulations are necessary.
Although PET exhibits in general a good quantification accuracy, specifically compared with other imaging modalities, it clearly reveals limitations in absolute quantification in small organs such as LN or the spleen; however, it still allows comparative and temporal quantification. There are large differences between ex vivo γ counting and in vivo PET quantification of the spleen (Fig. 2A) and lung. The density of the lung is low, overestimating the weight of the lung in the PET analysis (1 cm3 ≠ 1 g). The discrepancy of the quantification of the spleen is caused by a large volume of interest drawn on normalized PET images not corresponding to the long but flat shape of the spleen. This significant quantification difference is specifically reflected in spleens of intravenously injected mice that have, compared with spleens of intraperitoneally injected animals, a much higher signal leading to larger volumes of interest. In addition, the tiny spleen of a mouse yields a large partial-volume effect and low recovery (38). Thus, the in vivo quantification accuracy in such studies would significantly improve using combined PET/MR, yielding high resolution morphology and soft-tissue contrast, for an accurate placement of volumes of interest (1).
CONCLUSION
For a better understanding of the basic mode of T-cell homing and for the improvement of established T-cell–based treatment protocols, a knowledge of the homing dynamics of adoptively transferred T cells is essential and can be gained using PET imaging of 64Cu-PTSM–labeled cells. 64Cu-PTSM labeling of T cells can thus provide insights into the homing dynamics of TAA-specific T cells and into the effector phase of autoimmune diseases. The high-detection sensitivity of PET permits the labeling of only a small fraction of the cells in therapeutic interventions to monitor the trafficking routes in vivo without impairing the long-term functionality of most of the administered therapeutic cells. The established methods are therefore valuable for personalized medicine strategies.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This work was supported by the SFB685 and NIH R03 grants. The Department of Preclinical Imaging and Radiopharmacy serves as Reference and Training site for Siemens. Only institutional but no personal funds are related to this. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Nadine Bauer, Maren Koenig, Renate Nordin, and Denis Lamparter for excellent technical support.
Footnotes
Published online Jan. 16, 2014.
- © 2014 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- Received for publication May 14, 2013.
- Accepted for publication October 9, 2013.