


Visual Abstract

Abstract
Multidrug resistance–associated protein 1 (adenosine triphosphate–binding cassette subfamily C member 1 [ABCC1]) is abundantly expressed at the lung epithelial barrier, where it may influence the pulmonary disposition of inhaled drugs and contribute to variability in therapeutic response. The aim of this study was to assess the impact of ABCC1 on the pulmonary disposition of 6-bromo-7-11C-methylpurine (11C-BMP), a prodrug radiotracer that is intracellularly conjugated with glutathione to form the ABCC1 substrate S-(6-(7-11C-methylpurinyl))glutathione (11C-MPG). Methods: Groups of Abcc1(−/−) rats, wild-type rats pretreated with the ABCC1 inhibitor MK571, and wild-type control rats underwent dynamic PET scans after administration of 11C-BMP intravenously or by intratracheal aerosolization. In vitro transport experiments were performed with unlabeled BMP on the human distal lung epithelial cell line NCI-H441. Results: The pulmonary kinetics of radioactivity significantly differed between wild-type and Abcc1(−/−) rats, but differences were more pronounced after intratracheal than after intravenous administration. After intravenous administration, lung exposure (area under the lung time–activity curve from 0 to 80 min after radiotracer administration [AUClung]) was 77% higher and the elimination slope of radioactivity washout from the lungs (kE,lung) was 70% lower in Abcc1(−/−) rats, whereas after intratracheal administration, AUClung was 352% higher and kE,lung was 86% lower in Abcc1(−/−) rats. Pretreatment with MK571 decreased kE,lung by 20% after intratracheal radiotracer administration. Intracellular accumulation of MPG in NCI-H441 cells was significantly higher and extracellular efflux was lower in the presence than in the absence of MK571. Conclusion: PET with pulmonary administered 11C-BMP can measure ABCC1 activity at the lung epithelial barrier and may be applicable in humans to assess the effects of disease, genetic polymorphisms, or concomitant drug intake on pulmonary ABCC1 activity.
- 6-bromo-7-11C-methylpurine
- lung epithelial barrier
- multidrug resistance–associated protein 1
- PET
- pulmonary drug disposition
Chronic respiratory diseases, such as chronic obstructive pulmonary disease and asthma, affect millions of people all over the world. These diseases are frequently treated with orally inhaled drugs, such as bronchodilators and corticosteroids. Patient response to these drugs is characterized by a great degree of heterogeneity (1). Although efforts have been made to understand this variability at the level of the molecular targets of these drugs (2), another possible source of variability may be found in the absorption pathways across the pulmonary mucosa.
The airways are lined by epithelial cells, which form a physical barrier protecting the lungs from the external environment and express transport proteins belonging to the adenosine triphosphate–binding cassette (ABC) and solute carrier families (3). These transporters recognize several drugs used for the treatment of respiratory diseases as their substrates and may display interindividual variability in their abundance and activity, such as due to disease-associated factors, genetic polymorphisms, or drug–drug interactions. Although transporters have received a great deal of attention as important sources of pharmacokinetic variability of orally and intravenously administered drugs (4), comparatively little information is available on their role at the lung epithelial barrier, where they can regulate the pulmonary disposition of inhaled drugs. This knowledge gap can be potentially filled by nuclear imaging methods, such as PET or SPECT, in combination with radiotracers, which are administered as aerosols into the airways. Although this approach has been used before to assess the pulmonary disposition of some radiolabeled drugs (5–7), very few studies have focused on assessing the activity of pulmonary ABC or solute carrier transporters as factors potentially contributing to the pharmacokinetic variability of inhaled drugs (8,9).
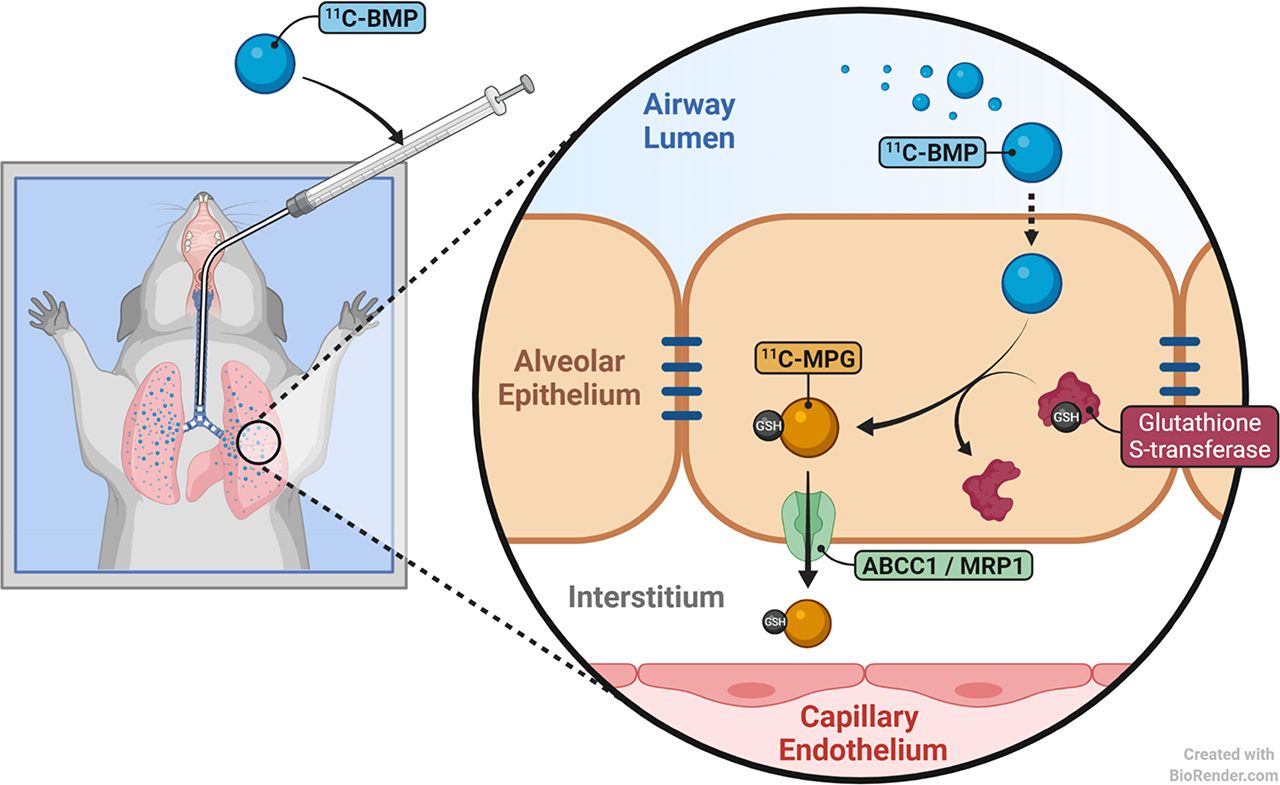
The most abundant ABC transporter at the lung epithelial barrier is multidrug resistance–associated protein 1 (ABC subfamily C member 1 [ABCC1]) (10). 6-bromo-7-11C-methylpurine (11C-BMP) is a PET tracer that has previously been used to assess ABCC1 activity in the mouse brain (11). 11C-BMP is a prodrug radiotracer that crosses cellular membranes by passive diffusion and is then intracellularly converted via glutathione-S-transferases into the corresponding glutathione conjugate S-(6-(7-11C-methylpurinyl))glutathione (11C-MPG), which is effluxed from cells by ABCC1 (Fig. 1) (11). One study found pronounced differences between wild-type and Abcc1(−/−) mice in the lung kinetics of radioactivity after intravenous administration of 11C-BMP, suggesting that this radiotracer may be suitable to assess pulmonary ABCC1 activity (12).

Schematic representation of 2 different radiotracer administration modes used in the present work. (A) Prodrug 11C-BMP is administered intravenously and crosses pulmonary capillary wall and basolateral membrane of pulmonary epithelial cells by passive diffusion. Intracellularly, 11C-BMP is converted by glutathione-S-transferases (GST) into corresponding glutathione conjugate 11C-MPG, which is effluxed across basolateral membrane of pulmonary epithelial cells by ABCC1, followed by transfer across capillary endothelial cells by unknown carriers. (B) Prodrug 11C-BMP is administered intratracheally and crosses luminal membrane of pulmonary epithelial cells by passive diffusion. Intracellularly, 11C-BMP is converted by GST into corresponding glutathione conjugate 11C-MPG, which is effluxed across basolateral membrane of pulmonary epithelial cells by ABCC1, followed by transfer across capillary endothelial cells by unknown carriers.
In the present study, we established a protocol to administer 11C-BMP into the airways of rats by aerosol to better understand the influence of ABCC1 on the pulmonary disposition of inhaled ABCC1 substrate drugs and to investigate whether pulmonary ABCC1 activity can be assessed with such an imaging protocol. To this end, we directly compared the lung kinetics of 11C-BMP–derived radioactivity between wild-type and Abcc1(−/−) rats after both intratracheal and intravenous administration of 11C-BMP (Fig. 1). To assess the potential translatability of our imaging protocol to humans, we additionally performed in vitro transport experiments on the human distal lung epithelial cell line NCI-H441 (13) with unlabeled BMP.
MATERIALS AND METHODS
Chemicals
Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich or Merck. The ABCC transporter inhibitor MK571 (5-(3-(2-(7-chloroquinolin-2-yl)ethenyl)phenyl)-8-dimethylcarbamyl-4,6-dithiaoctanoic acid sodium salt hydrate) was obtained from ApexBio Technology or from Santa Cruz. MK571 was dissolved in phosphate-buffered saline solution at a concentration of 150 mg/mL and injected intraperitoneally into rats at a volume of 2 mL/kg of body weight. Unlabeled BMP and MPG were synthesized as described in the literature (14).
Cell Culture
NCI-H441 human distal lung epithelial cells (ATCC HTB-174) were purchased from LGC Promochem and cultured according to a previously published protocol (13). Briefly, cells were cultured at 7 × 104 cells/cm2 in 24-well plates (Greiner Bio-One) using Gibco RPMI 1640 medium (Biosciences), supplemented with 5% fetal bovine serum and 1 mM sodium pyruvate. The cells were grown in a humidified atmosphere at 37°C and 5% CO2 for 5–8 d, until confluent monolayers were formed.
Radiotracer Synthesis
11C-BMP was synthesized as described before (15) in a decay-corrected radiochemical yield of 4.7% ± 1.1% (based on 11C-methane) with a radiochemical purity of more than 98% and a molar activity at the end of synthesis of 129 ± 74 GBq/μmol (n = 24). For intravenous or intratracheal administration to rats, 11C-BMP was formulated in 0.9% (w/v) aqueous saline solution.
Animals
Female wild-type Sprague–Dawley rats were obtained from Janvier-Labs. Female Abcc1(−/−) rats with a Sprague–Dawley genetic background were obtained from Horizon Discovery (formerly SAGE Labs Inc.). At the time of the experiment, animals weighed 239 ± 60 g. The animals were housed under controlled environmental conditions (21°C ± 2°C, 40%–70% humidity, 12-h light/dark cycle) and had free access to a standard laboratory-animal diet (LASQdiet Rod16; LASvendi GmbH) and water. An acclimatization period of at least 1 wk was allowed before the animals were used in the experiments. The study was approved by the national authorities (Amt der Niederösterreichischen Landesregierung), and the study procedures were in accordance with the European Communities Council Directive of September 22, 2010 (2010/63/EU). The animal experimental data reported in this study are in compliance with the ARRIVE (Animal Research: Reporting of in Vivo Experiments) guidelines.
Experimental Design
An overview of the examined animal groups is given in Table 1. Groups of wild-type and Abcc1(−/−) rats received a PET scan after intravenous or intratracheal administration of 11C-BMP. A further group of wild-type rats received a PET scan after intratracheal administration of 11C-BMP at 30 min after intraperitoneal injection of the ABCC transporter inhibitor MK571 (300 mg/kg). The dose of MK571 was based on previous work (12). In addition, 2 further groups of wild-type rats were used to determine the percentage of unchanged 11C-BMP in lung tissue both after intravenous and after intratracheal radiotracer administration.
Overview of Examined Animal Groups and Numbers
Intratracheal Aerosol Administration
Intratracheal aerosol was administered using a Microsprayer (Penn-Century, Inc.) connected to a FMJ-250 high-pressure syringe (Penn-Century, Inc.) containing 200 μL of formulated 11C-BMP solution. Under isoflurane anesthesia, the tip of the Microsprayer was introduced into the animals’ trachea using a laryngoscope. Aerosol was administered in a laminar flow cabinet to avoid contamination of the operator. Immediately after aerosol administration, PET imaging was performed. The mean time delay between radiotracer administration and the start of the PET data acquisition was 80 ± 18 s (range, 64–122 s).
PET Imaging
Imaging experiments were performed under isoflurane–air anesthesia. Animals were warmed throughout the experiment, and body temperature and respiratory rate were constantly monitored. A microPET Focus220 scanner (Siemens Medical Solutions) was used for PET imaging. 11C-BMP was administered either in a volume of 250 μL as an intravenous bolus via a tail vein or in a volume of 200 μL as an intratracheal aerosol (Table 1 shows the administered radioactivity amount and mass of unlabeled compound). At the start of intravenous administration or shortly after intratracheal administration, a 90-min dynamic PET scan (energy window, 350–750 keV; timing window, 6 ns) was initiated. At the end of the PET scan, the animals were euthanized. The lungs were collected for Western blot analysis of the abundance of ABCC1.
Western Blot Analysis
Rat lung tissue was lysed by suspending a small piece of the organ in 0.5 mL of ice-cold Invitrogen cell extraction buffer (Thermo Fisher Scientific) containing 1:25 ethylenediaminetetraacetic acid–free cOmplete (Roche) protease inhibitor cocktail followed by a 30-s homogenization step, using an Ultra-Turrax T25 (IKA). The lysate was then sonicated for 10 s and centrifuged at 17,000g for 10 min. The supernatant was transferred to a fresh tube and the pellet discarded. To harvest protein from NCI-H441 cells, a similar protocol without the homogenization step was followed. Tissue and cell samples were kept on ice for all protein extraction steps. The total protein amount in all samples was determined by a Pierce bicinchoninic acid assay (Thermo Fisher Scientific) according to the manufacturer’s instructions. Equal amounts of protein (40 µg) were mixed with loading buffer and heated to 37°C for 30 min, 72°C for 10 min, or 95°C for 5 min. Samples and 10 μL of PageRuler Plus (Thermo Fisher Scientific) were loaded onto an 8% sodium dodecyl sulphate–polyacrylamide gel, and electrophoresis was performed at 100 V. Afterward, a semidry transfer onto immunoblot polyvinylidene fluoride membranes (Bio-Rad) was performed at 22 V for 40 min using a Trans-Blot SD transfer cell (Bio-Rad). Membranes were blocked with 2% bovine serum albumin (w/v) in washing buffer (phosphate-buffered saline supplemented with 0.05% polysorbate 20) for 60 min at room temperature. Membranes were then washed twice and incubated at 4°C overnight with a monoclonal rat anti–multidrug resistance-associated protein 1 primary antibody (clone MRPr1, recombinant human multidrug resistance-associated protein 1, 1:50, GTX50894; GeneTex). A monoclonal mouse anti–β-actin primary antibody (clone AC-15, 1:5,000, A1978; Sigma-Aldrich) was used as the control. The following day, membranes were washed 3 times before incubation for 60 min at room temperature with a goat–antirat secondary antibody (1:5,000, 50320-200; Alpha Diagnostic) or a goat–antimouse secondary antibody (1:5,000, A4416; Sigma-Aldrich). After 3 further washing steps, peroxidase activity was determined using Immobilon western chemiluminescent horseradish peroxidase substrate (Millipore) in a ChemiDoc system (Bio-Rad).
Analysis of Glutathione Conjugation
In 2 groups of wild-type rats, the percentage of glutathione conjugation of 11C-BMP in the lungs was analyzed by thin-layer chromatography or high-performance liquid chromatography (HPLC) at 5 min after intravenous (n = 4) or intratracheal (n = 5) administration of 11C-BMP to anesthetized (isoflurane–air) animals. The animals were killed by intracardiac injection of Euthasol (pentobarbital sodium and phenytoin sodium; Vana GmbH) under deep anesthesia. The lungs were collected and homogenized using an Ultra Turrax T10 (IKA Laboratory Equipment), and proteins were precipitated with acetonitrile (1 mL/g of lung). The mixture was vortexed and then centrifuged (12,000g, 1 min, 21°C). The supernatant (5 µL) and diluted radiotracer solution as the reference were spotted on silica gel 60 F254 thin-layer chromatography plates (10 × 20 cm; Merck), and the plates were developed in ethyl acetate/ethanol (9/1, v/v). Detection was performed by placing the thin-layer chromatography plates on multisensitive phosphor screens (Perkin-Elmer Life Sciences). The screens were scanned at a 300-dpi resolution using a PerkinElmer Cyclone Plus Phosphor Imager (Perkin-Elmer Life Sciences). The retardation factor of 11C-BMP was 0.35, whereas the glutathione conjugate 11C-MPG had an retardation factor of 0.0. In addition, for the group in which 11C-BMP was administered intratracheally, the supernatant of the lung extract was analyzed with HPLC using an Agilent 1260 system. Analysis was carried out at 40°C using a Macherey-Nagel C18 high-density column (particle size: 5 μm, length: 250 mm, diameter: 10 mm), which was isocratically eluted with acetonitrile/water/aqueous acetic acid (40/60/0.1 v/v/v) at a flow rate of 1 mL/min. The HPLC eluate was monitored in series for radioactivity and ultraviolet absorption at a wavelength of 254 nm. 11C-BMP and 11C-MPG eluted with retention times of 13.8 min and 10.5 min, respectively.
Transport Studies
Uptake studies were performed using NCI-H441 cells cultured in 24-well plates according to a modification of a previously published protocol (16). All experiments were performed in Krebs–Ringer buffer (KRB; 116.4 mM NaCl, 5.4 mM KCl, 0.78 mM NaH2PO4, 25 mM NaHCO3, 5.55 mM glucose, 15 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 1.8 mM CaCl2, and 0.81 mM MgSO4; pH 7.4) at 37°C. Cell monolayers were incubated with KRB with or without MK571 (20 μM) for 60 min before experiments were initiated by exchanging the KRB for a solution of unlabeled BMP (100 μM in KRB). Monolayers were incubated for up to 150 min. After 15, 30, 60, 90, 120, and 150 min, respective cells were washed twice with ice-cold KRB and lysed with 1% (w/v) Triton X-100 (Dow Chemical Co.). The concentration of MPG in all samples was determined by HPLC. For standardization, the total protein amount of cell layers was determined by a Pierce bicinchoninic acid assay according to the manufacturer’s instructions.
Efflux studies of MPG were performed under conditions similar to those for the uptake experiments. Cell monolayers were loaded with unlabeled BMP solution (100 μM) for 60 min in the presence or absence of 20 μM of MK571. Subsequently, cell monolayers were washed twice with KRB, and either fresh KRB or KRB containing MK571 (20 μM) was added. At 15, 30, 45, 60, 75, 90, 105, and 120 min, 200-μL samples were taken from the cell supernatant and analyzed by HPLC. Each sample was replaced with an equal volume of the respective transport buffer. Uptake and efflux experiments were performed in triplicate using cells from 3 different passages.
HPLC Analysis
Samples were analyzed by HPLC according to a modification of a previously published method (11). A Hypersil BDS C18 column (250–4.6 mm; Thermo Fisher Scientific) was isocratically eluted (0.8 mL/min) at 40°C with water/acetonitrile (90/10) containing 0.1% trifluoroacetic acid, and ultraviolet detection was performed at 288 nm.
PET Data Analysis
The dynamic PET data were sorted into 24 frames, which incrementally increased in time length. PET images were reconstructed using Fourier rebinning of the 3-dimensional sinograms followed by 2-dimensional filtered backprojection with a ramp filter, giving a voxel size of 0.4 × 0.4 × 0.796 mm. Using AMIDE software (17), the left and right lungs were manually outlined on the PET images to derive average (left and right lung) time–activity curves expressed in units of SUV (SUV = [radioactivity per gram of administered radioactivity] × body weight). The area under the lung time–activity curve from 0 to 80 min after radiotracer administration (AUClung) was calculated using Prism 8 software (GraphPad Software). From the log-transformed lung time–activity curves, the elimination slope of radioactivity washout from the lungs (kE,lung, h−1) was determined by linear regression analysis (12). For the intratracheal group, the linear phase occurred from 0 to 6.25 min after radiotracer administration for wild-type rats and from 0 to 12.5 min for Abcc1(−/−) rats; for the intravenous group, the linear phase occurred from 17.5 to 80 min for both wild-type and Abcc1(−/−) rats.
Statistical Analysis
With Prism 8 software, differences between 2 groups were analyzed by a 2-sided t test, and differences between multiple groups were analyzed by 1-way ANOVA followed by a Dunnett multiple-comparison test. The level of statistical significance was set to a P value of less than 0.05. All values are given as mean ± SD.
RESULTS
We used PET to assess the influence of ABCC1 on the pulmonary kinetics of 11C-BMP–derived radioactivity after intravenous or intratracheal administration of 11C-BMP (Fig. 1). Two different approaches were followed: the use of Abcc1(−/−) rats and the use of pharmacologic inhibition of ABCC1 with MK571. To validate the Abcc1 knockout rat model, we first performed Western blot analysis of ABCC1 in lung tissue obtained from wild-type and Abcc1(−/−) rats, which confirmed absence of ABCC1 in the lungs of Abcc1(−/−) rats (Supplemental Fig. 1; supplemental materials are available at http://jnm.snmjournals.org). In Table 1, an overview of all examined animal groups and corresponding animal numbers is given. In total, 32 animals were used in the experiments. The study procedures were well tolerated by all animals, with no animals dying.
Thin-layer chromatography and HPLC analysis of radioactivity in the lungs at 5 min after intravenous or intratracheal administration of 11C-BMP revealed that more than 98% of total radioactivity was in the form of the corresponding glutathione conjugate 11C-MPG (Supplemental Fig. 2).
In Figure 2, serial PET images of wild-type and Abcc1(−/−) rats after intravenous administration of 11C-BMP are shown. These images revealed at later time points prolonged retention of radioactivity in the lungs of Abcc1(−/−) versus wild-type rats. The corresponding time–activity curves are shown in Figure 3A, revealing divergent profiles between the 2 rat strains after an initial distribution phase, that is, at time points more than 10 min after injection. As quantitative outcome parameters, we determined AUClung and kE,lung for the linear elimination phase occurring from 17.5 to 80 min after radiotracer injection (Supplemental Table 1). AUClung was significantly higher (+77%, Fig. 3B) and kE,lung significantly lower (−70%, Fig. 3C) in Abcc1(−/−) rats than in wild-type rats.

Representative coronal PET images at different time points after intravenous administration of 11C-BMP into 1 wild-type and 1 Abcc1(−/−) rat. L = lung; S = stomach; K = kidney.

Mean (±SD) time–activity curves (A), AUClung values (B), and kE,lung values (C) in wild-type rats (n = 4) and Abcc1(−/−) rats (n = 4) after intravenous administration of 11C-BMP. **P < 0.01, 2-sided t test.
Serial PET images after intratracheal administration of 11C-BMP are shown in Figure 4. In early time frames after the start of the PET acquisition (0–1.5 min), radioactivity was homogeneously distributed in the lungs, supporting the conclusion that intratracheal administration was successful. In 2 of 15 animals, intratracheal administration of 11C-BMP failed (the esophagus and stomach were visible on the PET images; Supplemental Fig. 3), and these 2 animals (both Abcc1(−/−)) were excluded from analysis. The effect of Abcc1 knockout on the pulmonary kinetics of 11C-BMP–derived radioactivity was more pronounced after intratracheal than after intravenous administration and predominantly affected the early part of the pulmonary time–activity curves (from ∼0–20 min after the start of the PET acquisition; Fig. 5A). Accordingly, kE,lung was determined from 0 to 6.25 min for wild-type rats and from 0 to 12.5 min for Abcc1(−/−) rats. kE,lung values were significantly lower in Abcc1(−/−) rats than in wild-type rats (±86%; Fig. 5C), and AUClung values were significantly higher (+352%; Fig. 5B; Supplemental Table 1). We also assessed the effect of the prototypical ABCC transporter inhibitor MK571 on the pulmonary kinetics of radioactivity after intratracheal administration of 11C-BMP (Figs. 4 and 5). Similar to Abcc1 knockout, pretreatment with MK571 resulted in a significant decrease in kE,lung as compared with untreated wild-type rats (−20%; Fig. 5C; Supplemental Table 1), although the effect was less pronounced than in Abcc1(−/−) rats. AUClung did not differ between untreated and MK571-pretreated wild-type rats (Fig. 5B).

Representative coronal PET images at different time points after start of PET acquisition after intratracheal administration of 11C-BMP into 1 wild-type rat, 1 Abcc1(−/−) rat, and 1 wild-type rat pretreated with MK571 at 30 min before radiotracer administration (300 mg/kg, intraperitoneally). There was a short delay between intratracheal administration and start of PET acquisition (80 ± 18 s). L = lung; S = stomach; K = kidney.

Mean (±SD) time–activity curves (A), AUClung values (B), and kE,lung values (C) in wild-type rats (n = 6), Abcc1(−/−) rats (n = 4), and MK571-pretreated wild-type rats (n = 3) after intratracheal administration of 11C-BMP. *P < 0.05. ***P < 0.001. ****P < 0.0001. P values are for 1-way ANOVA with Dunnett multiple-comparison test.
We performed in vitro transport experiments with unlabeled BMP using monolayers of NCI-H441 cells (Fig. 6). Intracellular accumulation of the glutathione conjugate MPG over time was significantly higher when MK571 was added to the incubation buffer than in the absence of MK571 (Fig. 6A). Moreover, the extracellular efflux of MPG from BMP-loaded NCI-H441 cells was significantly lower in the presence than in the absence of MK571 (Fig. 6B).

(A) Time-dependent accumulation of MPG in NCl-H441 cell monolayers. Cells were incubated with unlabeled BMP (100 μM) in KRB without or with MK571 (20 μM), and intracellular glutathione conjugate MPG was measured at different time points with HPLC. (B) Time-dependent efflux of MPG from NCl-H441 cell monolayers. Cells were loaded with unlabeled BMP (100 μM) in presence or absence of MK571 (20 μM). Then, extracellular efflux of MPG in presence or absence of MK571 (20 μM) was measured at different time points with HPLC. Data are shown as mean (±SD) from 3 independent experiments performed each in triplicate. **P < 0.01, 2-sided t test. ***P < 0.001, 2-sided t test.
DISCUSSION
The aim of this study was to investigate whether ABCC1 activity at the lung epithelial barrier can be measured with intratracheally administered 11C-BMP, a prodrug radiotracer that is rapidly transformed in vivo into a radiolabeled ABCC1 substrate (12). We found pronounced increases in the pulmonary exposure to 11C-BMP–derived radioactivity in Abcc1(−/−) rats as compared with wild-type rats, highlighting the important role ABCC1 may potentially play in controlling the lung disposition of inhaled ABCC1 substrate drugs. We also demonstrated proof of the concept that there is an ABCC1-mediated drug–drug interaction at the level of the pulmonary epithelium with the prototypical ABCC transporter inhibitor MK571.
ABCC1 is an ABC transporter with a wide substrate spectrum that includes several different drugs and has a broadly accepted role in tumor multidrug resistance (18). ABCC1 is the most abundant ABC transporter in the human lung, where it is located mainly in the basolateral membrane of bronchial and alveolar epithelial cells (10,19). Quantitative targeted absolute proteomics has revealed a remarkable 18-fold variability in the abundance of ABCC1 in the bronchial region among 5 different donors (10). Several inhaled drugs for the treatment of respiratory diseases were found to interact in vitro in lung epithelial cell lines with ABCC1, including formoterol, budesonide, ipratropium bromide, and N-acetyl cysteine (20). Moreover, there is evidence for disease-induced alterations in the abundance of pulmonary ABCC1 (e.g., in chronic obstructive pulmonary disease) (21) as well as for an effect of cigarette smoke extract on pulmonary ABCC1 activity (22). Given its high abundance in the pulmonary epithelium and broad substrate spectrum, ABCC1 may potentially contribute to interindividual variability in the lung disposition of inhaled drugs and thereby affect therapeutic response.
The applicability of drugs used for the treatment of respiratory diseases as transporter imaging probes for PET or SPECT may be limited by their lack of selectivity among individual pulmonary ABC transporters. A potentially more powerful approach would therefore be the use of dedicated transporter imaging probes. The SPECT tracer 99mTc-sestamibi is a substrate of P-glycoprotein (ABC subfamily B member 1, ABCB1), and a substrate of ABCC1 has been used to image ABCB1 activity in tumors (23). In a study in which 99mTc-sestamibi was administered as an aerosol into the airways of lung cancer and pneumothorax patients, a positive correlation between the pulmonary clearance half-life of 99mTc-sestamibi and the expression of ABCC1 in surgically excised lung tissue was found (9). However, the utility of 99mTc-sestamibi as a radiotracer to image pulmonary ABCC1 activity is compromised by the fact that 99mTc-sestamibi is also a substrate of ABCB1, which works in a transport direction opposite to that of ABCC1 in the pulmonary epithelium (3).
11C-BMP has been developed to visualize ABCC1 activity in the brain (11). Because ABCC1 preferably transports organic anions, 11C-BMP has been designed as a prodrug with good passive permeability, which effectively penetrates the blood–brain barrier. Once inside the brain, it is quantitatively converted by intracellular glutathione-S-transferases into the hydrophilic glutathione conjugate 11C-MPG, which has poor passive permeability and whose elimination from the brain into the blood is mediated in mice by ABCC1, organic anion transporter 3 (solute carrier family 22 member A8), and multidrug resistance–associated protein 4 (ABC subfamily C member 4, ABCC4) (11,24). One study found a 17-fold decrease in the elimination rate of radioactivity from the lungs in Abcc1(−/−) mice relative to wild-type mice after intravenous administration of 11C-BMP (12). No differences in pulmonary elimination of radioactivity were found in mice lacking ABCB1 and breast cancer resistance protein (ABC subfamily G member 2, ABCG2), suggesting selectivity of this imaging probe for ABCC1 over ABCB1 and ABCG2, both of which are also located in the lung epithelium (12).
On the basis of these promising earlier results, we hypothesized that 11C-BMP may be suitable to measure the influence of ABCC1 at the pulmonary epithelium on the lung disposition of inhaled drugs. To this end, we set up a simple and effective protocol for intratracheal administration of aerosolized 11C-BMP using the Microsprayer device. We used rats rather than mice, as the larger size of rats facilitated intratracheal radiotracer administration. We could show that 11C-BMP is, in rat lung tissue, very efficiently converted into its glutathione conjugate within 5 min after administration (Supplemental Fig. 2), as was consistent with previous mouse data (12).
We used a novel, commercially available Abcc1 knockout rat model, in which we demonstrated by Western blot analysis absence of ABCC1 in lung tissue (Supplemental Fig. 1). A direct comparison of intratracheal with intravenous administration of 11C-BMP (Fig. 1) revealed that ABCC1 exerts a greater influence on the pulmonary disposition of radioactivity after intratracheal administration. Knockout of Abcc1 led to a marked increase in AUClung and a decrease in kE,lung (Fig. 5), as was consistent with a role for ABCC1 in mediating the efflux of its substrates across the basolateral membrane of epithelial cells into the interstitium (Fig. 1) (3). It can be expected that Abcc1 knockout led to decreased systemic absorption of 11C-BMP–derived radioactivity, as was in good agreement with our observation that the kidneys as the main excretory organ were clearly visible on the PET images of wild-type rats but not of Abcc1(−/−) rats after intratracheal 11C-BMP administration (Fig. 4). The pronounced effect of Abcc1 knockout on elimination of radioactivity from the lungs (Fig. 5) suggested lack of functional redundancy between ABCC1 and other transporters (e.g., ABCC4) in mediating efflux of the radiolabeled glutathione conjugate from pulmonary epithelial cells. However, as 11C-MPG presumably cannot cross cell membranes by passive diffusion, some active carriers must also be involved in its transfer across endothelial cells of pulmonary capillaries (Fig. 1). Because ABCC1 does not appear to be expressed in the pulmonary endothelium (19), these carriers could be—as at the mouse blood–brain barrier (24)—solute carrier family 22 member A8 and ABCC4, both of which have been detected in human lung tissue (10). The influence of ABCC1 on the pulmonary disposition of clinically used ABCC1 substrate drugs will most certainly depend on the individual characteristics of each drug (passive permeability, interaction profile with other pulmonary transporters) and may therefore not always be of the same magnitude as for 11C-BMP.
We could demonstrate the sensitivity of our imaging protocol to measure a drug-induced modulation of ABCC1 activity by systemic administration of the non–subtype-selective ABCC transporter inhibitor MK571. We observed a 20% reduction of kE,lung in MK571-pretreated rats as compared with untreated rats (Fig. 5), suggesting partial inhibition of ABCC1 in pulmonary epithelial cells and possibly other anion transporters involved in the pulmonary elimination of radioactivity by MK571. Our results indicate that our imaging protocol has the potential to assess ABCC1-mediated drug–drug interactions in the lung.
Similar to the effects seen in vivo in rats, in vitro transport experiments demonstrated MK571-sensitive flux of unlabeled MPG across monolayers of the human distal lung epithelial cell line NCI-H441 (Fig. 6), which highly expresses ABCC1 (Supplemental Fig. 1). These results support the possibility that MPG is a substrate of human ABCC1 (14) and that imaging of pulmonary ABCC1 activity may also be feasible in humans with either intratracheally or intravenously administered 11C-BMP. Our imaging protocol may allow a better understanding of the influence of lung disease on the pulmonary disposition of clinically used, inhaled ABCC1 substrate drugs and potential tailoring of the treatment schedules to the individual pulmonary-barrier properties of each patient.
CONCLUSION
PET with intratracheally administered 11C-BMP can measure ABCC1 activity at the lung epithelial barrier as a possible determinant of the pulmonary disposition of inhaled ABCC1 substrate drugs. Our imaging protocol may be potentially applicable in patients to assess the effects of disease, genetic polymorphisms, or drug–drug interactions on pulmonary ABCC1 activity. The imaging-based assessment of the barrier properties of the pulmonary epithelium may open an entirely new field of research, which may contribute to a better understanding of the sources of variability in treatment response to inhaled therapeutics and ultimately provide patients with more effective drugs.
DISCLOSURE
Financial support was provided by the Lower Austria Corporation for Research and Education (NÖ Forschungs-und Bildungsges.mbH [NFB]) (LS17-009, to Oliver Langer). No other potential conflict of interest relevant to this article was reported.
KEY POINTS
QUESTION: Can ABCC1 at the lung epithelial barrier affect the pulmonary disposition of inhaled ABCC1 substrates and thereby potentially contribute to variability in the therapeutic response to drugs used for the treatment of chronic respiratory diseases?
PERTINENT FINDINGS: We found that knockout of Abcc1 led to severalfold increases in lung radioactivity exposure after intratracheal administration of 11C-BMP. The effect of ABCC1 on the pulmonary disposition of radioactivity was more pronounced after intratracheal than after intravenous radiotracer administration.
IMPLICATIONS FOR PATIENT CARE: PET with intratracheally administered 11C-BMP may be applied in patients to assess interindividual differences in lung epithelial barrier properties and thereby potentially tailor treatment schedules to the individual characteristics of each patient.
Acknowledgments
We thank Mathilde Löbsch for help in conducting the PET experiments and Dr. Ben Forbes (King’s College London, U.K.) for providing the Microsprayer.
Footnotes
Published online Apr. 13, 2020.
- © 2020 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication February 24, 2020.
- Accepted for publication March 19, 2020.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.