


Abstract
Aromatase (an enzyme that converts androgens to estrogens) in the brain is involved in neuroprotection, synaptic plasticity, and regulation of sexual and emotional behaviors. To investigate the physiologic and pathologic importance of aromatase in the brain, including in humans, we here report the development of a novel PET probe for aromatase, 11C-cetrozole, which allows noninvasive quantification of aromatase expression. Methods: 11C-cetrozole was synthesized by the C-11C-methylation method developed by our group. In vitro autoradiography of frozen sections and a binding study with rat brain homogenates were conducted to demonstrate the specific binding and the dissociation constant. PET studies with anesthetized rhesus monkeys were performed to analyze the dynamics in the brain. Results: In vitro and in vivo studies using 11C-cetrozole showed its superiority in brain aromatase imaging in terms of specificity and selectivity, compared with previously developed 11C-vorozole. PET studies showed that 11C-cetrozole had a higher signal-to-noise ratio, providing a sharper image than 11C-vorozole, because the radioactive metabolite of 11C-vorozole was taken up into the brain. High specific binding of 11C-cetrozole was observed in the amygdala and hypothalamus, and we also noted binding in the nucleus accumbens of rhesus monkeys for the first time. Conclusion: These results suggest that PET imaging with newly developed 11C-cetrozole is suitable for quantifying the expression of brain aromatase in vivo, possibly providing critical information regarding the functional roles of aromatase in human neurologic and emotional disorders.
Aromatase, an estrogen synthetic enzyme that converts testosterone and androstenedione to estradiol and estrone (1), is considered to play an important role in various brain functions. Studies using aromatase knockout mice demonstrated that aromatase deficiency leads to susceptibility of neurons to neuronal apoptosis in the frontal cortex of aged females (2) and in the hypothalamus of aged males (3). These mice are also susceptible to neuronal toxins such as domoic acid (4) and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, which is used to produce animal models of Parkinson's disease (5). In behavior studies, aromatase knockout mice exhibit depressive-like behavior (6), aggressive behaviors leading to infanticide (7,8), repressed aggression to intruders (9,10), impaired spatial reference memory (11), and disrupted sexual behavior (7,10). These impaired behaviors can be considerably recovered by specifically introducing human aromatase into the brain of aromatase knockout mice, strongly suggesting a relationship between aromatase and brain functions (9). A relationship between aromatase and Alzheimer's disease or autism was also suggested (12,13). In normal conditions, aromatase is expressed in neurons, but after brain injury, neogenetic aromatase is expressed in astrocytes, which are considered to protect neurons (14). Although these results suggest the importance of local estrogen biosynthesis in the brain, the precise roles of aromatase in neurologic and emotional disorders are not fully understood.
Studying the relationships between aromatase and brain functions would be aided by the ability to obtain quantitative measurements of the expression level and the ability to localize aromatase directly in living animals and humans. For this purpose, the Uppsala University group developed the first PET probe, 11C-vorozole, based on the aromatase inhibitor developed for breast cancer therapy (15). In collaboration with them, we first succeeded in obtaining sufficient PET images in the rhesus monkey brain in which specific binding of 11C-vorozole was detected in the amygdala and hypothalamic regions (16). A method for isolating 11C-vorozole labeled in the right position was established later on (17). Up to now, using 11C-vorozole, the distribution of aromatase in the brain of rats, rhesus monkeys (16), baboons (17), and humans (18) and the effect of anabolic-androgenic steroids on aromatase (19,20) have been demonstrated. Although 11C-labeled sulfonanilide analogs (21), 11C-labeled sulfamate derivatives (dual aromatase–steroid sulfatase inhibitors) (22), and 11C-letrozole (23) have also been developed as aromatase imaging PET probes, 11C-vorozole is still the most popular probe for imaging brain aromatase. However, 11C-vorozole has some drawbacks. First, the greatest disadvantage is that a certain amount of nonspecific signal is observed in PET studies. This may be caused by metabolic cleavage of the N-11C-methyl bond followed by unintended reuptake of the radioactive metabolite into the brain. Second, the precursor of 11C-vorozole, norvorozole, is not easily available. Although we have recently established a practical preparative method (24), synthesis of 11C-vorozole still requires a multistep procedure and optical resolution of the racemic mixture. And third, it was reported that all 3 possible N-11C-methylation at the triazole moiety of norvorozole proceeded in the reaction with 11C-CH3I, decreasing the yield and complicating isolation of the desired probe (17).
To address these issues and explore a novel PET probe for aromatase imaging, we focused on YM511, an aminotriazole derivative developed as an aromatase inhibitor for breast cancer (25). Because this compound showed high inhibitory competence with a half maximal inhibitory concentration (IC50) of 0.12 nM for human placental aromatase and had a simple structure that was easy to synthesize, we planned to modify and optimize this compound to obtain a more efficient PET probe. Here, we describe the development of a new probe, named 11C-cetrozole, in which the bromophenyl moiety of YM511 was changed to an 11C-methylphenyl group. The new compound works better than 11C-vorozole, with improved characteristics suitable for brain aromatase imaging.
MATERIALS AND METHODS
Synthesis of Pinacol Boranyl Precursor 1 and Tributylstannyl Precursor 3 for 11C-Cetrozole (11C-2) and Nonradiolabeled Cetrozole (2)
Detailed procedures are provided in the supplemental data (available at http://jnm.snmjournals.org).
Synthesis of 11C-Cetrozole (11C-2) (Fig. 1)
11C-carbon dioxide was produced by a 14N(p,α)11C reaction using a cyclotron (CYPRIS HM-18; Sumitomo Heavy Industries Ltd.) and then converted to 11C-methyl iodide by treatment with lithium aluminum hydride, followed by hydriodic acid. The obtained 11C-methyl iodide was used for palladium(0)-mediated rapid 11C-methylation as follows (26).
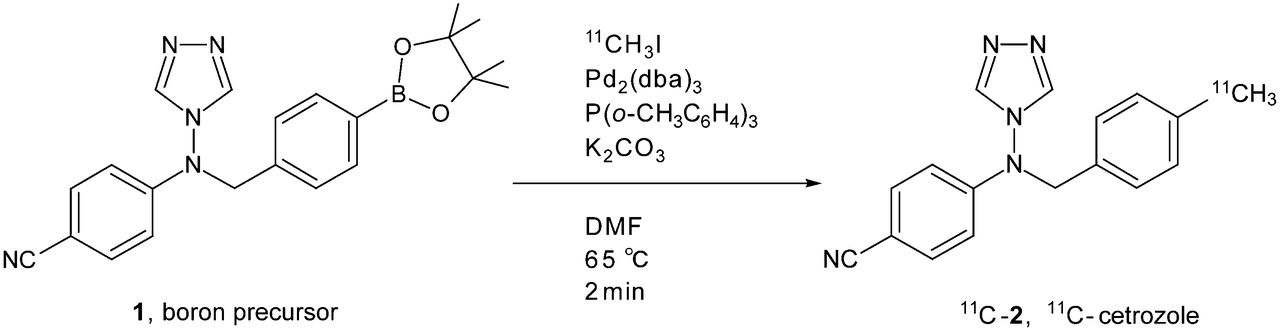
Radiosynthesis of 11C-cetrozole (11C-2) by rapid 11C-methylation using pinacol boranyl precursor 1. DMF = dimethylformamide.
11C-methyl iodide was transferred by a stream of helium gas (30 mL/min) into a solution of boron precursor 1 (4.0 mg, 10 μmol), Pd2(dba)3 (2.9 mg, 2.5 μmol), P(o-tolyl)3 (4.0 mg, 13 μmol), and K2CO3 (4.0 mg, 30 μmol) in dimethylformamide (0.5 mL) at room temperature. The resulting mixture was heated at 65°C for 2 min. Salts and palladium residue in the reaction mixture were removed by a solid-phase extraction, washing with 1 mL of a solution of CH3CN:30 mM CH3COONH4 (35:65). The combined elutes were injected into preparative high-performance liquid chromatography (HPLC) with a γ detector (mobile phase: CH3CN:30 mM CH3COONH4 [35:65]; column: COSMOSIL AR-II C18 [Nacalai], 10 × 250 mm; guard column: SUMIPAX Filter PG-ODS [Sumika]; flow rate: 6 mL/min; ultraviolet detection: 254 nm; retention time of 11C-2: 13.5 min). The desired fraction was collected into a flask, and then the organic solvent was removed under reduced pressure. The desired 11C-labeled compound was dissolved in a mixture of polysorbate 80 (0.05 mL), propylene glycol (0.3 mL), and saline (4 mL). The total synthesis time including HPLC purification and radiopharmaceutical formulation for intravenous administration was 30 min. The radioactivity of 11C-2 prepared for administration was 2.6–6.2 GBq, and the specific radioactivity was 90–127 GBq/μmol. The identification of 11C-2 was confirmed by coinjection with the unlabeled authentic sample on analytic HPLC with a γ detector (mobile phase: CH3CN:30 mM CH3COONH4 [40:60]; column: COSMOSIL AR-II C18 [Nacalai], 4.6 × 100 mm; flow rate: 1 mL/min; ultraviolet detection: 254 nm; retention time of 11C-2: 4.5 min). The chemical purity and the radiochemical purity were greater than 95%. The decay-corrected yield based on 11CH3I in a reaction vial was 54%–60%.
11C-cetrozole could also be prepared efficiently from tributylstannyl precursor 3 (Supplemental Fig. 1).
Synthesis of 11C-Vorozole
11C-vorozole was synthesized according to a previously published procedure (17) using norvorozole as the precursor (24). The specific radioactivities were in the range of 22–90 GBq/μmol, and the radiochemical purity was greater than 99.5%.
Animals
Male and female Sprague–Dawley rats (8–12 w) were housed under constant temperature and humidity and were maintained under a 12-h light–dark cycle with free access to water and food. Male adult rhesus monkeys (Macaca mulatta; weight, 4.6–7.1 kg) were housed individually, food was given twice a day, and water was available ad libitum. Animals were maintained and handled in accordance with the recommendations of the U.S. National Institutes of Health, and the study was approved by the Animal Care and Use Committee of Kobe Institute at RIKEN (MAH18-05, MAH21-13, MAH21-10).
In Vitro Assays Using 11C-Cetrozole
In vitro autoradiography using rat brain sections was performed as described previously (16). The method is described in the legend to Supplemental Figure 2.
Ligand saturation experiments with brain homogenate were performed as described previously (16) with suitable modifications. The method is described in the legend to Supplemental Figure 3.
PET Studies
PET scans were obtained with monkeys using 11C-cetrozole (n = 4) or 11C-vorozole (n = 5). All monkeys were used for the tracer dose studies, and 2 monkeys were used for one additional blocking study with unlabeled vorozole at a dose of 150 μg/kg. The monkeys were sedated with ketamine hydrochloride (15 mg/kg, intramuscularly), and venous cannulae were placed in the saphenous veins for further continuous anesthesia with propofol (10 mg/kg/h) and PET tracer injection. Before the emission scan, a transmission scan was obtained for 30 min for attenuation correction. Each tracer (11C-cetrozole, 106–535 MBq; 11C-vorozole, 117–231 MBq) was administered intravenously as a bolus. The monkeys were scanned for 90 min (4 × 30, 3 × 60, 2 × 150, 2 × 300, and 7 × 600 s) using a microPET Focus 220 (Siemens). For quantitative data analysis with an input function, 2 monkeys underwent additional PET with arterial blood sampling at the following time points: 8, 16, 24, 32, 40, 48, 56, 64, 72, 90, and 150 s and 4, 6, 10, 20, 30, 45, 60, and 90 min. The samples at 64 s and 10, 20, 30, and 60 min were also used for metabolite analysis.
Analysis of Radiolabeled Metabolites in Brain and Plasma
Rats received radioactivity (∼110 MBq/body) and were sacrificed at 5-, 10-, and 20-min time points (n = 3 at each time point). Plasma and whole brains, which were removed immediately and homogenized in water, were used for analyses of 11C-cetrozole and 11C-vorozole metabolites. The plasma of 2 monkeys was taken at time points of 64 s and 4, 10, 20, 30, and 60 min during PET scans and used for analyses of 11C-cetrozole and 11C-vorozole metabolites. Measurements of radiolabeled metabolites in the brain and plasma were performed as described previously (27).
Analysis of PET Data
For quantitative analysis of PET images (i.e., kinetic modeling), PMOD software (PMOD Technologies Ltd.) was used. Volumes of interest were delineated in the cerebellum, amygdala, hypothalamus, nucleus accumbens, thalamus, white matter, and temporal cortex. Decay-corrected time–activity curves were generated for each brain region and arterial blood plasma and parent compound. The time–activity curves for plasma and metabolites were fitted to a 3 exponential model and a Hill function, respectively. The data with arterial blood sampling were analyzed with a Logan plot (28), and the total distribution volume (Vt) in each brain region was calculated. The data without arterial blood sampling were analyzed with a Logan reference tissue model based on average k2’ (29), using the cerebellum as a reference. Nondisplaceable binding potential (BPnd) and distribution volume ratio (DVR), which are linear functions of enzyme availability, were calculated.
RESULTS
In Vitro Assays Using 11C-Cetrozole and Biodistribution
In an autoradiographic study with 11C-cetrozole of the rat brain, a high level of binding was observed in the medial amygdala and bed nucleus of the stria terminalis, and a moderate level of binding was seen in the hypothalamus similar to the study with 11C-vorozole (Supplemental Fig. 2) (16). The binding was clearly displaced by application of excess amounts of unlabeled vorozole. Little signal was detected in other brain regions including the cerebellum. Male rats showed higher binding of 11C-cetrozole in the medial amygdala and bed nucleus of stria terminalis than female rats (Supplemental Fig. 3).
The Scatchard plot analysis of 11C-cetrozole in the rat medial amygdala showed a single binding site of aromatase with the binding affinity (Kd) value of 0.11 ± 0.02 nM (n = 3) (Supplemental Fig. 4).
The biodistribution of 11C-cetrozole in rats showed relatively high accumulation in the small intestine at 30 min after the administration (Supplemental Fig. 5).
Analysis of 11C-Cetrozole and 11C-Vorozole Metabolites in Brain and Plasma
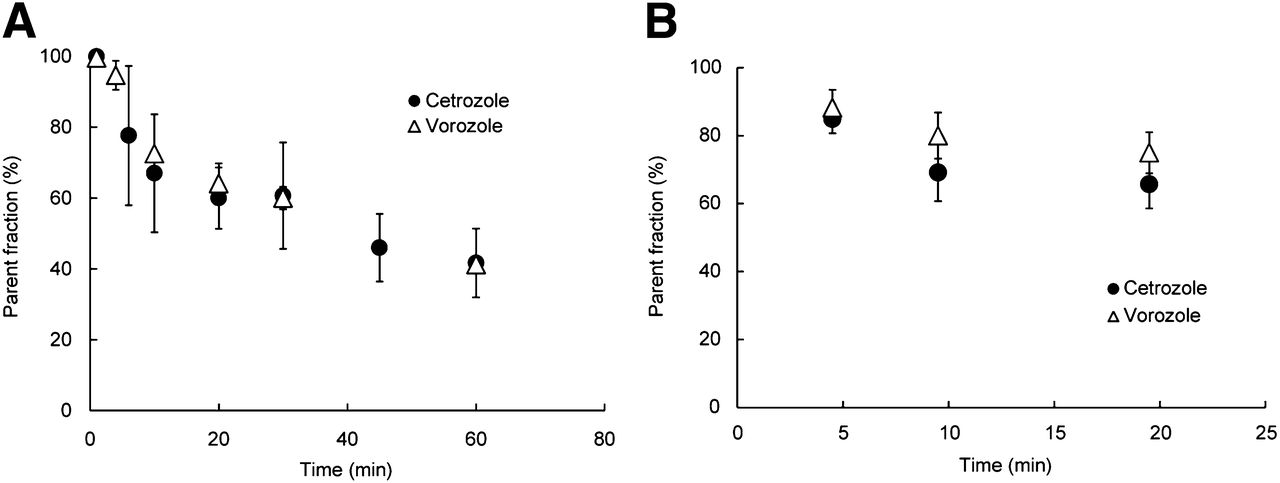
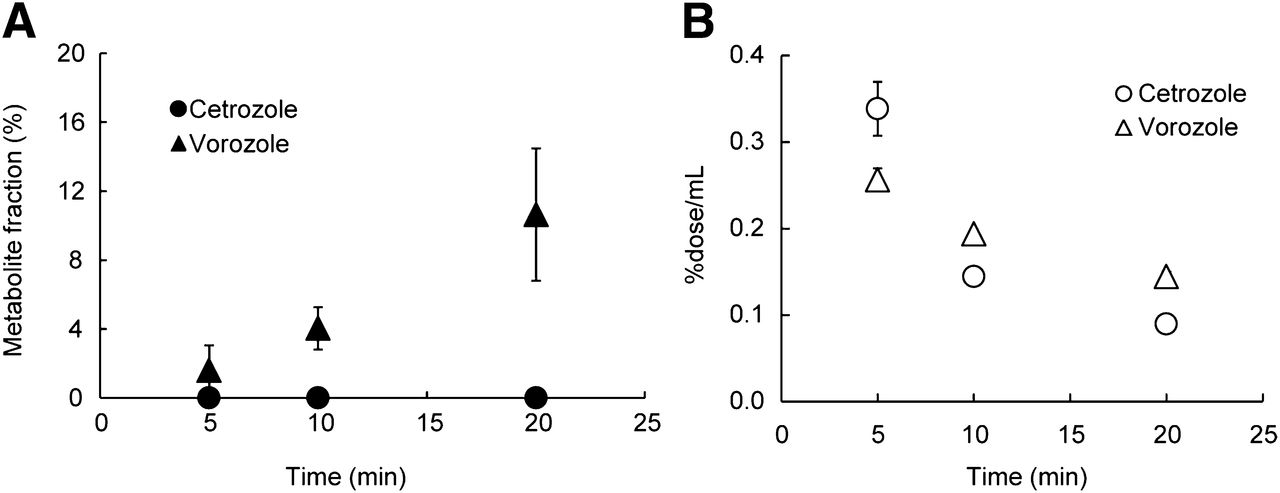
The proportions of the parent compound in plasma decreased over time, and we observed no difference between 11C-cetrozole and 11C-vorozole in both monkeys and rats (Fig. 2). At 30 min after administration of the tracers, about 60% of 11C-cetrozole and 11C-vorozole remained intact; the level was about 40% at 60 min in rhesus monkey plasma (Fig. 2A). Similarly, we found no difference between the proportions of the parent compound of each tracer in the plasma of rats (Fig. 2B). To also investigate metabolites in the brain, the radiolabeled parent compound and metabolites were measured in the rat brain. Although radiolabeled metabolites of 11C-vorozole accumulated in the brain over time, radiolabeled metabolites of 11C-cetrozole were not detected even 20 min after administration of the tracer (Fig. 3). Representative radiochromatograms are shown in Supplemental Figure 6. 11C-cetrozole and 11C-vorozole with retention times of 3.0 and 2.6 min gave one major metabolite at 3.7 and 0.4–0.5 min, respectively. The amount of 11C-vorozole metabolite increased over time both in plasma and in brain. In contrast, 11C-cetrozole metabolites accumulated only in plasma but not in the brain.
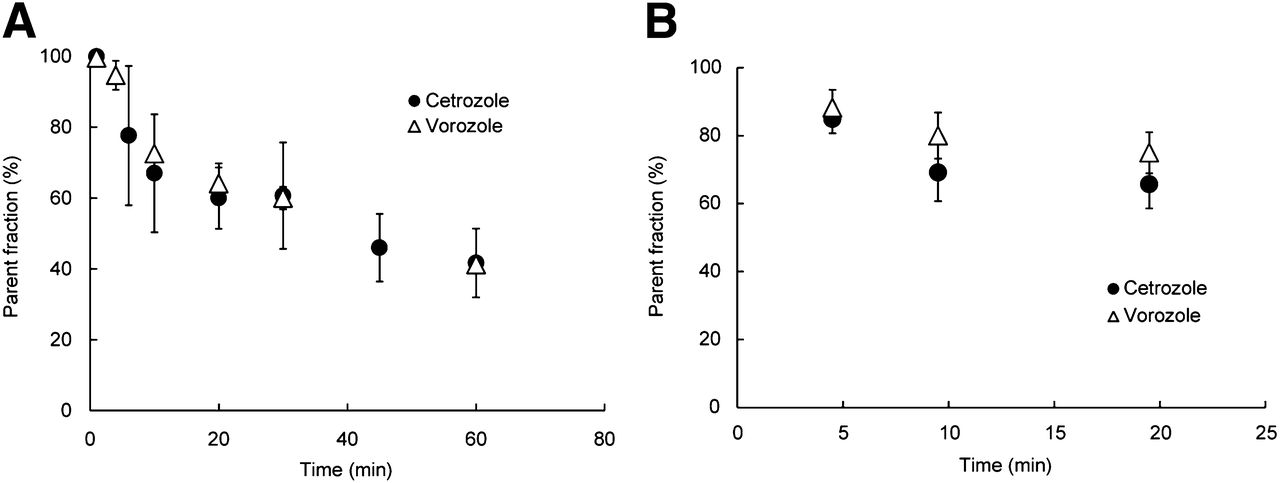
Elimination curve of intact 11C-cetrozole in plasma of rhesus monkeys (A, n = 4, mean ± SEM) and rat (B, n = 3, mean ± SEM). 11C-cetrozole was metabolized over time as much as 11C-vorozole. Curve was fitted to percentage of intact 11C-cetrozole.

Radiolabeled metabolite fraction (A, n = 3, mean ± SEM) and time–activity of 11C-cetrozole and 11C-vorozole (B, n = 3, mean ± SEM) in rat brain. (A) Radiolabeled metabolite of 11C-cetrozole was not detected even 20 min after injection of radioactivity, in contrast to 11C-vorozole for which radiolabeled metabolite accumulated in brain over time. (B) Both 11C-cetrozole and 11C-vorozole were eliminated from brain over time.
PET Studies
DVR images of 11C-cetrozole and 11C-vorozole showed higher binding activity in the amygdala with both PET tracers (Fig. 4). However, the accumulation of the radiotracers in the amygdala and hypothalamus was delineated more clearly with 11C-cetrozole than with 11C-vorozole. Additionally, the nucleus accumbens was clearly delineated only with 11C-cetrozole. Time–activity curves showed that the clearance rate of 11C-vorozole was slower than that of 11C-cetrozole, especially in the late phase (Fig. 5). The standardized uptake value (SUV) of 11C-vorozole plateaued after 30 min of scan time, and that in the amygdala and nucleus accumbens seemed to slightly increase until 90 min (Fig. 5B). The SUV of 11C-cetrozole continued to decrease (Fig. 5A).

DVR images of 11C-cetrozole (A and C) and 11C-vorozole (B and D) in amygdala (A and B) and nucleus accumbens (C and D) of rhesus monkey (coronal slices). DVRs in amygdala and nucleus accumbens regions are 0.4–1.5 and 0.8–1.5, respectively. DVR of 11C-cetrozole was higher in these regions than that of 11C-vorozole.
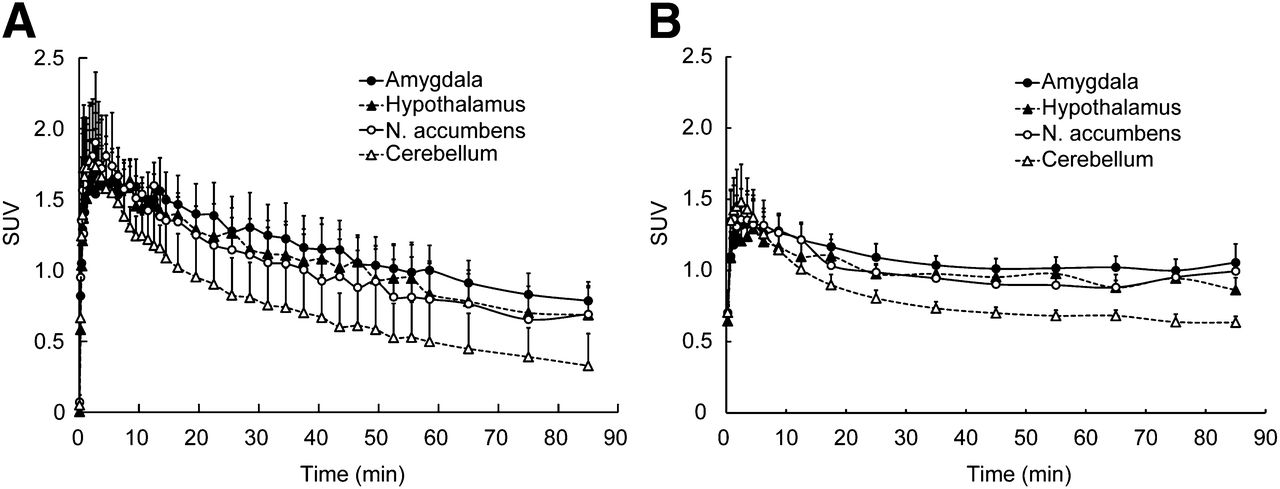
Time–activity curves of 11C-cetrozole (A, n = 4, mean ± SEM) and 11C-vorozole (B, n = 5, mean ± SEM) in rhesus monkey brain. SUV of high-binding regions (amygdala, hypothalamus, and nucleus accumbens) and reference region for Logan reference tissue model analysis (cerebellum) are shown. N. = nucleus.
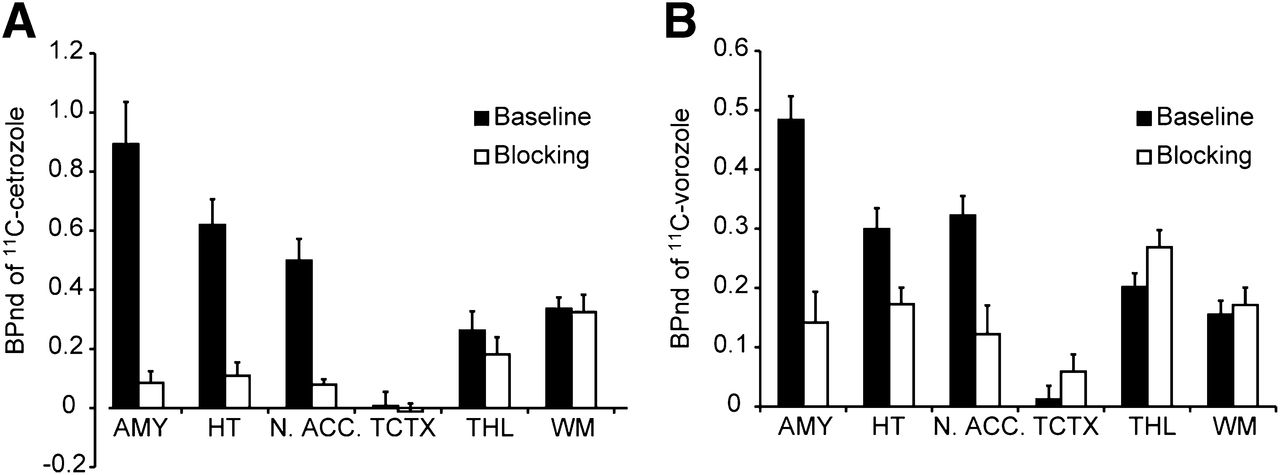
The BPnd of 11C-cetrozole was high in the amygdala, hypothalamus, and nucleus accumbens (Fig. 6A). In these 3 regions, the BPnd of 11C-vorozole was also high, although relatively low BPnd was observed in the hypothalamus (Fig. 6B). The BPnd in the cortices was low with both tracers. Most of the signals in the white matter could be regarded as nonspecific binding. The BPnd of 11C-cetrozole was higher than that of 11C-vorozole by 88%, 170%, and 55% in the amygdala, hypothalamus, and nucleus accumbens, respectively. 11C-cetrozole also showed better signal-to-noise ratios (ratio of baseline to blocking) in the amygdala, hypothalamus, and nucleus accumbens: 11C-cetrozole, 10.5, 5.7, and 6.3, respectively; 11C-vorozole, 3.4, 1.7, and 2.6, respectively (Fig. 6).
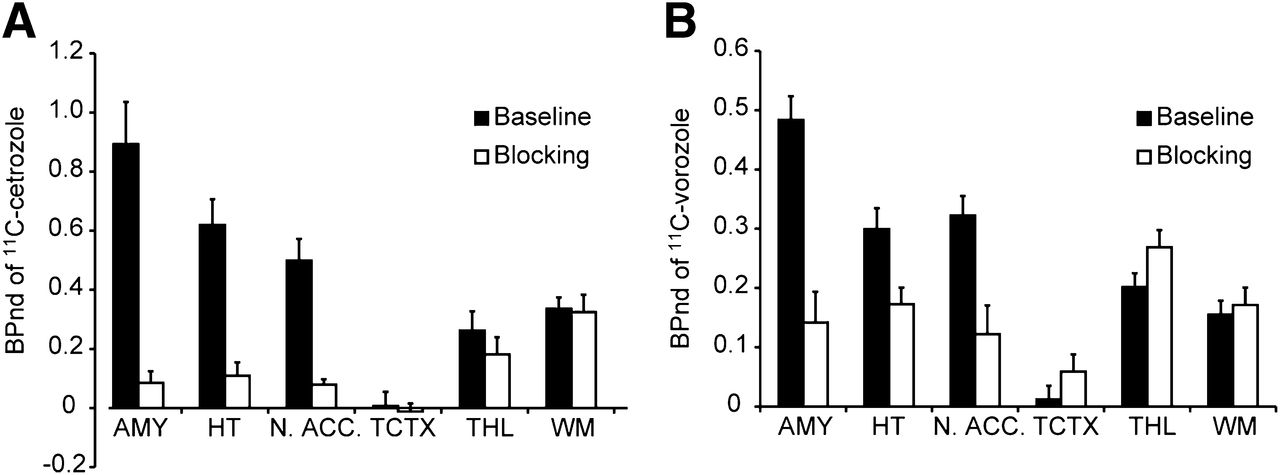
BPnd of 11C-cetrozole (A, n = 4, mean ± SEM) and 11C-vorozole (B, n = 5, mean ± SEM) with (blocking) or without (baseline) unlabeled vorozole in amygdala (AMY), hypothalamus (HT), nucleus accumbens (N. ACC.), temporal cortex (TCTX), thalamus (THL), and white matter (WM) in rhesus monkey. 11C-cetrozole showed better signal-to-noise ratio than 11C-vorozole in amygdala, hypothalamus, and nucleus accumbens.
DISCUSSION
To evaluate the in vivo function of aromatase playing important roles in the brain, molecular PET imaging is one of the most straightforward approaches. 11C-labeled vorozole, the first aromatase inhibitor–type PET probe developed at the Uppsala University PET Centre, allowed imaging of the dynamic status of aromatase in the living body including the brain (15). PET studies using this probe clarified the distribution of aromatase in living rhesus monkeys (16,24,27), baboons (17), and also humans (18), strongly suggesting a role for aromatase in brain functions. For example, an increase in aromatase expression was detected in the hypothalamic nuclei of rats and rhesus monkeys by the repeated administration of anabolic androgenic steroids for 3 wk daily, inducing psychologic adverse effects (27). However, during PET studies using 11C-vorozole in rhesus monkeys, we showed increased brain SUV in the late time phase of PET scans, suggesting reuptake of the radiolabeled metabolites of 11C-vorozole into the brain. Because the N-11C-methyl bond in the 11C-vorozole structure was presumed to be susceptible to metabolic cleavage producing undesired 11C-labeled metabolites, we designed a new PET probe with a more metabolically stable C-11C-methyl structure. On the basis of a potent aromatase inhibitor YM511 with an IC50 of 0.12 nM on human placental aromatase (25), we developed a new compound, 11C-cetrozole, by changing the bromophenyl moiety of YM511 to 11C-methylphenyl group. Cetrozole does not have an asymmetric center, making its synthesis easy, and showed potent aromatase inhibitory activity comparable with vorozole. The introduction of the 11C-methyl group to the phenyl moiety was efficiently achieved using the Suzuki- or Stille-type rapid 11C-methylation reaction of corresponding boryl or stannyl precursors developed by our group (Fig. 1; Supplemental Fig. 1) (26).
Metabolite analyses of plasma and brain of rats and monkeys administered 11C-cetrozole or 11C-vorozole revealed that both tracers were metabolized at a similar rate in plasma in both rats and monkeys. However, in the rat brain, the 11C-vorozole metabolites were detected, but 11C-cetrozole metabolites were not, indicating that 11C-vorozole metabolites entered the brain in radiolabeled form but those of 11C-cetrozole did not. This means that the PET signal from 11C-vorozole originated from both the parent compound and the metabolites but that of 11C-cetrozole was mostly from the parent compound. This idea was also supported by the observation of increased brain SUV for 11C-vorozole in the late time phase in the rhesus monkey study; increased brain SUV was not observed in the 11C-cetrozole study. These results indicated that 11C-cetrozole may be a more preferable tracer than 11C-vorozole for the quantitative measurement of aromatase in the brain. Even in a human PET study (18), the time–activity curve of 11C-vorozole in the brain was almost the same as that of the monkey: specifically, that increased radioactivity was also observed in the late phase.
The Scatchard plot for the binding assay using the rat amygdala homogenate showed linearity of the slope, indicating that 11C-cetrozole bound to a single binding site of aromatase, similar to 11C-vorozole (16). The Kd value was 0.11 nM, indicating that 11C-cetrozole had a higher affinity to aromatase than 11C-vorozole, which has a Kd of 0.60 nM (16). The IC50 values obtained with the aromatase inhibition assay using marmoset placenta were 1.27 and 0.66 nM for cetrozole and vorozole, respectively (Supplemental Fig. 7). Our PET data showed binding potentials that were higher for 11C-cetrozole than 11C-vorozole and may represent the Kd of each compound rather than inhibitory activity. The binding of 11C-cetrozole was inhibited by the administration of 1 μM unlabeled vorozole, suggesting that cetrozole binds to an androgen-specific cleft of aromatase, similar to vorozole (30).
To analyze PET data, the method of using a reference region may be less quantitative than the method of using an arterial blood sample as an input function. To explore the possibility of using reference region–based analysis for 11C-cetrozole, we compared the result from the Logan reference tissue model based on the average k2’ (BPnd) and that from arterial blood sampling as the input function (Vt). The cerebellum was selected as a reference region because it contains negligible amounts of aromatase messenger RNA and enzymatic activity (31). The radioactivity in all examined brain regions was reduced to the level of the cerebellum with pretreatment with an excess amount of unlabeled vorozole. Additionally, to confirm that 11C-cetrozole does not specifically bind in the cerebellum, both 1-tissue- and 2-tissue-compartment models were applied to the kinetics of radioactivity in the cerebellum. The kinetics fit better with the 1-tissue-compartment model with smaller values of Akaike information criteria, suggesting the suitability of the cerebellum as the reference region. To compare the calculated with the input function of arterial blood and the BPnd determined with the Logan noninvasive model using the cerebellum as the reference region, Vts of all examined brain regions were normalized to the Vt of the cerebellum. The normalized values of Vt and BPnd were well correlated (r > 0.99), and the difference between these 2 values was less than 5%. Thus, the Logan noninvasive model using the cerebellum could be used for data analysis of brain PET imaging with 11C-cetrozole.
As a result, high specific binding of 11C-cetrozole was observed in the amygdala, hypothalamus, and nucleus accumbens. The signal-to-noise ratios of BPnd in these 3 regions were 10.4, 5.6, and 6.2, respectively, which were better than those for 11C-vorozole: 3.4, 1.7, and 2.6, respectively. Early studies measuring the messenger RNA level showed that aromatase is expressed at high levels in the amygdala and hypothalamus of rhesus monkeys (31). A previous PET study with 11C-vorozole showed high specific binding in the amygdala and relatively weak binding in the hypothalamus (16), results that were reproduced in this study. The present study with 11C-cetrozole clearly demonstrated that the amygdala and hypothalamus contained high amounts of aromatase in the rhesus monkey brain. In addition, the nucleus accumbens was clearly delineated only with 11C-cetrozole. DVR images of 11C-vorozole had too-high nonspecific binding to delineate the nucleus accumbens clearly. The localization of aromatase in the nucleus accumbens was also supported by Western blotting (Takahashi et al., unpublished data, 2012). The nucleus accumbens is thought to be involved with reward, motivation, addiction, and aggression (32,33) through the dopaminergic system, which is partly modulated by estrogen (34). Therefore, aromatase in the amygdala, hypothalamus, and also the nucleus accumbens may be involved in emotional behavior. Further research is needed to clarify this idea.
CONCLUSION
We developed 11C-cetrozole, which showed a higher signal-to-noise ratio suitable for the quantitative analysis of aromatase expression in the amygdala, hypothalamus, and nucleus accumbens in the brain of nonhuman primates. 11C-cetrozole will be a powerful tool for exploring the functional roles of aromatase in human neurologic and emotional disorders.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This work was supported in part by a consignment expense for the Molecular Imaging Research Programs entitled “Research Base for Exploring New Drugs” from the Japanese Ministry of Education, Culture, Sports, Science and Technology and JSPS KAKENHI grants 22791155 and 25830024. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Dr. Yasuhisa Tamura, Masahiro Kurahashi, Akihiro Kawasaki, Chiho Takeda, Yumiko Katayama, Tomoko Mori, and Emi Hayashinaka (RIKEN) for their assistance with the PET study and Masayo Ishikawa (Tokyo Institute of Technology) and Ayako Hosoya (Tokyo Medical and Dental University) for HRMS analysis.
Footnotes
Published online Mar. 27, 2014.
- © 2014 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication August 22, 2013.
- Accepted for publication January 21, 2014.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}