


Abstract
Phosphodiesterase 10A (PDE10A) plays a central role in striatal signaling and is implicated in several neuropsychiatric disorders, such as movement disorders and schizophrenia. We performed initial brain kinetic modeling of the novel PDE10A tracer 18F-JNJ-42259152 (2-[[4-[1-(2-18F-fluoroethyl)-4-(4-pyridinyl)-1H-pyrazol-3-yl]phenoxy]methyl]-3,5-dimethyl-pyridine) and studied test–retest reproducibility in healthy volunteers. Methods: Twelve healthy volunteers (5 men, 7 women; age range, 42–77 y) were scanned dynamically up to 135 min after bolus injection of 172.5 ± 10.3 MBq of 18F-JNJ42259152. Four volunteers (2 men, 2 women) underwent retest scanning, with a mean interscan interval of 37 d. Input functions and tracer parent fractions were determined using arterial sampling and high-performance liquid chromatography analysis. Volumes of interest for the putamen, caudate nucleus, ventral striatum, substantia nigra, thalamus, frontal cortex, and cerebellum were delineated using individual volumetric T1 MR imaging scans. One-tissue (1T) and 2-tissue (2T) models were evaluated to calculate total distribution volume (VT). Simplified models were also tested to calculate binding potential (BPND), including the simplified reference tissue model (SRTM) and multilinear reference tissue model, using the frontal cortex as the optimal reference tissue. The stability of VT and BPND was assessed down to a 60-min scan time. Results: The average intact tracer half-life in blood was 90 min. The 2T model VT values for the putamen, caudate nucleus, ventral striatum, substantia nigra, thalamus, frontal cortex, and cerebellum were 1.54 ± 0.37, 0.90 ± 0.24, 0.64 ± 0.18, 0.42 ± 0.09, 0.35 ± 0.09, 0.30 ± 0.07, and 0.36 ± 0.12, respectively. The 1T model provided significantly lower VT values, which were well correlated to the 2T VT. SRTM BPND values referenced to the frontal cortex were 3.45 ± 0.43, 1.78 ± 0.35, 1.10 ± 0.31, and 0.44 ± 0.09 for the respective target regions putamen, caudate nucleus, ventral striatum, and substantia nigra, with similar values for the multilinear reference tissue model. Good correlations were found for the target regions putamen, caudate nucleus, ventral striatum, and substantia nigra between the 2T-compartment model BPND and the SRTM BPND (r = 0.57, 0.82, 0.70, and 0.64, respectively). SRTM BPND using a 90- and 60-min acquisition interval showed low bias. Test–retest variability was 5%–19% for 2T VT and 5%–12% for BPND SRTM. Conclusion: Kinetic modeling of 18F-JNJ-42259152 shows that PDE10A activity can be reliably quantified and simplified using a reference tissue model with the frontal cortex as reference and a 60-min acquisition period.
Phosphodiesterase 10A (PDE10A) is a dual-substrate enzyme with a restricted distribution, predominantly in the human brain in medium spiny neurons of the striatum, the substantia nigra, the nucleus accumbens and olfactory tuberculum (1), and—to a much lower extent—the hippocampus and cerebral cortex (1,2). PDE10A hydrolyzes the important second messengers cyclic adenosine monophosphate (cAMP) and cyclic guanosine monophosphate, with an approximately 20-times higher affinity for cAMP (3,4). Through hydrolysis of cAMP, PDE10A downregulates protein kinase A activity, which relies on cAMP for activation and regulates the phosphorylation of various intracellular targets downstream of protein kinase A such as DARPP-32 as an integrator of dopamine and glutamate signals (5). This role in striatal signaling has made PDE10A an enzyme of particular interest because it is likely involved in several neuropsychiatric disorders. This hypothesis is supported by preclinical studies with PDE10A inhibitors, showing that pharmacologic inhibition of PDE10A leads to activation of medium spiny neurons (MSNs) and alteration of behavioral aspects modulated by the striatal circuit (6). Disruption of the conditioned avoidance response, an established preclinical model for antipsychotic activity, was observed after the administration of PDE10A inhibitor TP-10 (7) and MP-10 (8), and reduced exploratory locomotor activity was seen (7). Such pharmacologic alterations were not found in PDE10A knockout mice (9). PDE10A inhibition may constitute a new approach for the treatment of disorders with altered activity of medium spiny neurons, such as schizophrenia (8), Huntington disease (10), and addiction (11).
A suitable PDE10A tracer that allows in vivo quantification of PDE10A would lead to a better understanding of the role of PDE10A activity in health and disease, can be used as a tool for early clinical evaluation of emerging PDE10A medication, and may also present new diagnostic opportunities. Several attempts to quantify PDE10A using PET have been recently published. 11C-MP-10 has been evaluated in nonhuman primates (2,12), but although it showed good affinity and high selectivity for PDE10A, a lipophilic radiolabeled metabolite hampered further human application. The preclinical evaluation of 11C-papaverine in rhesus macaques showed low retention (13). We previously evaluated 18F-JNJ41510417 in PDE10A knockout and wild-type mice and in rodents (14). Although this tracer showed high selectivity and affinity, in vivo brain kinetics were slow, hampering accurate quantification. Therefore, it was decided to investigate other candidates based on similar leads. 18F-JNJ-42259152, or 2-[[4-[1-(2-18F-fluoroethyl)-4-(4-pyridinyl)-1H-pyrazol-3-yl]phenoxy]methyl]-3,5-dimethyl-pyridine, is also an antagonist of PDE10A—with a lower half-maximal inhibitory concentration of 1.6 nM (15), a low polar surface area, and a plasma protein binding of 99.5%—and is 1,000-fold more selective than for the other phosphodiesterase enzymes (Janssen Research and Development, unpublished data, 2011). In humans, a first-in-man biodistribution and dosimetry study showed that the tracer was safe and displayed promising kinetics for further quantitative evaluation of PDE10A activity in the nigrostriatal circuit (16).
The aim of this study was to perform full brain kinetic modeling of 18F-JNJ-42259152 and study test–retest reproducibility in healthy human volunteers. Additionally, we evaluated the simplified reference region model to investigate the possibility of noninvasively quantifying PDE10A and reducing acquisition time to allow clinical application in a variety of neuropsychiatric patient populations.
MATERIALS AND METHODS
Subjects
Healthy subjects aged 40–80 y were recruited for this monocentric, academic study through advertisements in a local community newspaper and on the departmental Web site. All subjects were screened for neuropsychiatric and other medical disorders and were subjected to a physical examination, blood and urine testing (including toxicology for drugs), and structural brain MR imaging. Subjects who were current users of any illicit drugs (including recreational use) or had any history of drug or alcohol abuse were excluded. A history of tobacco use was also reason for exclusion unless the subject had completely stopped for at least 3 mo. No medication use was allowed.
In total, 12 healthy volunteers (5 men, 7 women) between the ages of 42 and 77 y (mean age ± SD, 57.8 ± 10.3 y) were included. To determine test–retest reliability over a period of weeks, 4 of these subjects (2 men, 2 women; mean age ± SD, 61.5 ± 13.9 y) underwent a second PET scan. Written informed consent was obtained from all the volunteers before participation. The study was approved by the local Ethics Committee and performed in accordance to the latest version of the World Medical Association Declaration of Helsinki.
Radiotracer Preparation
The precursor for tracer synthesis was obtained from Janssen Research and Development. Radiolabeling was performed on-site using 1-18F-fluoro-2-bromoethane, which was distilled into the precursor solution (containing Cs2CO3 in anhydrous dimethylformamide). The reaction mixture was injected onto a high-performance liquid chromatography (HPLC) system eluted with a mixture of sodium phosphate (0.01 M, pH 2.2) and ethanol (15). The radiochemical purity was more than 95%, and specific activity was always higher than 100 GBq/μmol (mean ± SD, 222 ± 156 GBq/μmol; range, 110–534 GBq/μmol). In this study, the mean administered mass dose was 0.52 ± 0.38 μg.
Pharmacologic Properties of JNJ-42259152
JNJ-42259152 is a potent and highly selective inhibitor of PDE10A. JNJ-42259152 inhibits the cAMP-hydrolyzing activity of purified recombinant PDE10A with potency pIC50 (known log of inhibition constant at 50%) of 8.82 for the human enzyme. JNJ-42259152 is more than 1,000-fold more selective for PDE10A, relative to all other human phosphodiesterases. Various radioligand binding experiments (including CEREP standard binding profiling) indicated that JNJ-42259152 at a concentration of 10 μM did not show affinity for most of the human receptors, ion channels, or transporters (Janssen Research and Development, unpublished data, 2011). In vivo blocking and displacement studies conducted in rats are the subject of a future preclinical evaluation study to be described elsewhere (17).
Imaging Procedure and Safety Assessments
All subjects were scanned on a HiRez Biograph 16 PET/CT camera (Siemens) in 3-dimensional mode. For reasons of standardization, subjects fasted for 4 h before PET. They received a fast bolus injection of 173.5 ± 6.3 MBq of 18F-JNJ-42259152. The dynamic PET brain scan was initiated simultaneously with the radiotracer bolus injection and consisted of 2 scanning segments. The first 90-min segment started on tracer injection, and the second 30-min segment started at 105 min after injection. Both segments were acquired in list-mode. A low-dose CT scan (80-kV tube potential, 11 mAs) was obtained at the beginning of both PET acquisition segments for attenuation correction.
Test–retest reproducibility was evaluated in 4 healthy volunteers. The interval between scanning was 26.3 ± 10.5 d.
For reconstruction, the first segment of each PET scan was rebinned in 26 frames with progressively increasing frame durations (4 × 15, 4 × 60, 2 × 150, and 16 × 300 s), with a total duration of 90 min. The second segment was rebinned in 6 frames of 300 s. All frames were Fourier-rebinned and reconstructed using an ordered-subset expectation maximization algorithm with 5 iterations and 8 subsets and a uniform gaussian postsmoothing filter of 5 mm. The reconstructed volume consisted of 81 slices of 2-mm thickness with a matrix size of 128 × 128 and reconstructed pixel size of 2 mm. Emission data were corrected for attenuation and scatter.
As part of the screening and for further anatomic coregistration with PET to delineate volumes of interest, volumetric magnetization prepared rapid acquisition gradient-echo T1 (repetition time, 0 ms; echo time, 4 ms; flip angle, 12°; inversion time, 300 ms; matrix, 256 × 256; 160 sagittal contiguous slices of 1 mm) and standard T2 MR images were obtained.
To further assess the safety of 18F-JNJ-42259152 administrations, before and after the PET acquisition a physical examination (including blood pressure measurement and neurologic evaluation), laboratory blood analyses (which included blood cell counts and liver and kidney function tests), and electrocardiography were performed. An electrocardiogram and blood pressure measurements were also taken at other predefined time points during the PET acquisition. All adverse experiences were monitored during and immediately after the PET scan (follow-up monitoring after the procedure comprised a telephone interview at 24 and 14 d after scanning).
Plasma Input Curve and Metabolite Determinations
In all subjects, arterial and venous blood sampling was performed manually through an arterial and venous cannula in the lower radial artery and an antecubital vein, respectively (in the opposite arm of injection), to determine the arterial input function curve and radiometabolite fraction, determined both in arterial and venous blood. Per imaging session, 2-mL arterial samples were drawn at the following times: every 10 s for the first 100 s; at 120, 140, and 160 s after injection; and at 3, 4, 5, 8, 10, 15, 20, 30, 40, 60, 90, and 120 min after injection. Additional 5-mL blood samples (both arterial and venous) were drawn at 5, 10, 20, 40, 90, and 120 min for metabolite measurement. All samples were immediately placed on ice before analysis.
Blood samples were centrifuged for 10 min at 3,000 rotations per minute. Authentic JNJ-42259152 (10 μg) was added to 0.8-mL of plasma, and the mixture was then injected onto the HPLC system as such. The HPLC eluent was collected in 3 fractions: 2–5 min, 10–12 min (metabolite fractions), and 13–16 min (parent fraction). A γ-counter (1480 Automatic γ-counter Wallac Wizard 3; PerkinElmer) was used to measure the amount of radioactivity in each fraction.
Image Analysis and Kinetic Modeling
Motion correction was performed on the dynamic dataset using PMOD (PMOD Inc.) (with sum of squared differences minimization algorithm). The retest PET scan was rigidly registered to the first PET scan and subsequently to the individual MR images (using normalized mutual information and the first 16 image frames). Three-dimensional volumes of interest were then manually delineated on individual MR images for the following brain regions: caudate nucleus, putamen, ventral striatum, substantia nigra, medial frontal cortex, thalamus, and cerebellum. The latter 3 regions were considered as potential reference regions with low PDE10A expression (1).
Tissue Distribution Volume and Metabolite Kinetics
The rate of tracer metabolism was modeled by fitting (1 − A) + A × exp(−ln(2) × t/B) to the parent fraction fparent(t), where A (0 < A < 1) represents a fast exponential metabolizing component with corresponding half-life B. This way, a parent fraction estimate was available for each time point of the arterial plasma activity samples. Using the corresponding parent fraction estimates, we subdivided arterial plasma activity samples into activity values for the intact tracer fraction and a potential metabolite fraction. This way, 2 arterial input functions were generated: a metabolite-corrected arterial input function for modeling intact tracer kinetics and a metabolite arterial input curve that can be used to model metabolite kinetics.
To quantify regional tracer kinetics, total distribution volumes VT (18) were estimated using a standard 1-tissue-compartment model (1TCM) and 2-tissue-compartment model (2TCM) together with a radiometabolite-corrected arterial input function. Time–activity curve data points were weighted according to frame duration and radioactive decay.
Because initial results showed that a 2TCM was preferable to a 1TCM for describing tracer kinetics of brain regions with presumably low PDE10A expression, we evaluated 3 possible assumptions to further investigate these findings.
A first assumption was that a 2TCM was needed for appropriate modeling of a small fraction of specific binding (2TCM[spec]). A second hypothesis was that an aspecific tracer binding with slower kinetics would require a 2TCM (2TCM[aspec]). Although both assumptions use a mathematically equivalent 2TCM, boundary conditions in terms of constraints applied to the 2TCM rate constants are different.
In the case of 2TCM[spec], the 2 compartments representing free tracer and nonspecific binding of a 3-tissue-compartment model (3TCM) were merged into 1 compartment representing the nondisplaceable distribution volume VND (Fig. 1). This simplification corresponds to the most commonly used interpretation of 2TCM. Because specific binding is modeled by a separate compartment, VND corresponding to K1/k2 was estimated for the frontal cortex and considered constant for the target and other potential reference regions. Other 2TCM parameters—K1, k3, and k4, and therefore VS (distribution volume for specific binding)—were estimated independently for each brain region.
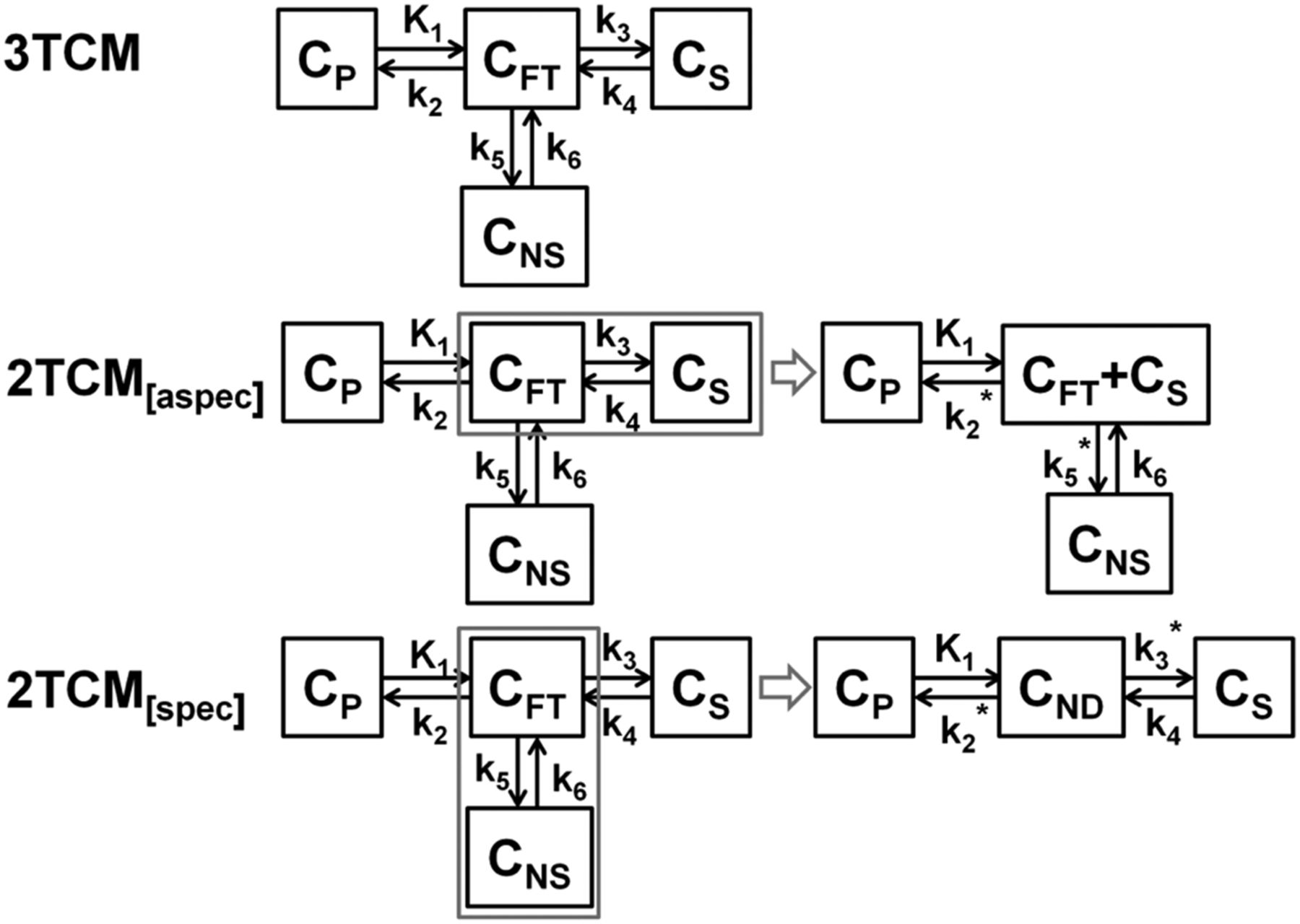
Schematic overview of 2 possible simplifications of 3TCM. For 2TCM[aspec], compartments representing free and specific binding are merged, and remaining compartment represents nonspecific binding. Latter nonspecific binding is assumed constant for target and reference regions. For 2TCM[spec], compartments representing free and nonspecific binding are merged to nondisplaceable tissue compartment, and remaining compartment represents specific binding. Former nondisplaceable compartment is assumed constant for reference and target regions. CFT = free tracer concentration; CND = nondisplaceable tracer concentration; CNS = nonspecific tracer concentration; CP = plasma tracer concentration; CS = specific tracer concentration.
For the 2TCM[aspec] hypothesis, we used an approach as proposed by Watabe (Hiroshi Watabe, written communication, 2012), where kinetic behavior in the reference tissue is described on the basis of 2 tissue compartments: one for free tracer and one for nonspecific binding. Basically, this means that 2 compartments of the 3TCM representing, respectively, free tracer and specific binding, can be merged and the remaining 2TCM compartment represents nonspecific binding (Fig. 1). Assuming that the amount of nonspecific binding in reference and target tissues was the same, the distribution volume for the remaining tissue compartment representing nonspecific binding was estimated for the frontal cortex and set to this value for both target and other potential reference regions. No further restrictions were applied to other 2TCM parameters K1, k2, and k3, and therefore VND was estimated independently for each brain region.
A third hypothesis, the possible confounding effect of a brain-penetrating radiometabolite on the quantification, was explored, where one compartment accounts for intact tracer kinetics (1TCM[intact]) and a second compartment describes the kinetics of a brain-entering radiometabolite. Intact tracer and metabolite kinetics were modeled using 2 separate parallel 1-tissue compartments with 2 separate parallel input functions. The first 1-tissue compartment with the arterial intact tracer input function described the specific and nonspecific binding of the intact tracer (1TCM[intact]), and the other 1-tissue compartment with the arterial metabolite input function was assumed to account for the brain uptake of radiometabolite. Again, assuming the radiometabolite fraction for the reference and target regions are the same, the distribution volume of the radiometabolite VT,M was estimated for the frontal cortex and set to the same value for the nigrostriatal target regions. VT,M determined with this model was compared with the distribution volume for nonspecific binding VNS obtained with 2TCM[aspec] and with the fast metabolized fraction intact tracer in the arterial blood compartment.
VT values for 1TCM, 2TCM, 2TCM[spec], and 2TCM[aspec] and the corresponding plasma to tissue uptake rates K1 were evaluated. 1TCM, 2TCM, 2TCM[spec], and 2TCM[aspec] fittings were compared using Akaike information criteria (AIC). 2TCM VT values were used as reference values for further correlation analysis to assess a possible systematic bias of the outcome measure.
Binding Potential (BPND) and Reference Tissue Models
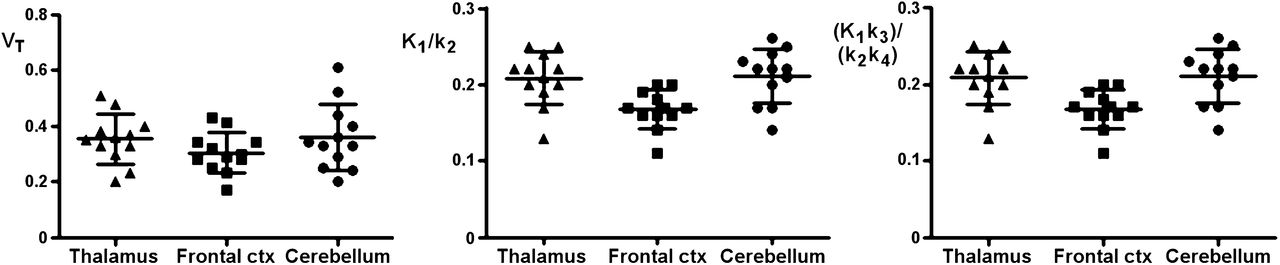
Nondisplaceable BPND using the simplified reference tissue model (SRTM) (19) and multilinear reference tissue model (MRTM) (20) were evaluated with the frontal cortex as reference tissue. The frontal cortex was considered the reference tissue of choice because it provided consistently the lowest values for 2TCM VT, K1/k2, and (K1/k2) × (k3/k4) when compared with other candidate reference tissues such as the thalamus and cerebellum (Fig. 2).

Overview of VT, K1/k2, and (K1/k2) × (k3/k4) for 2TCM for candidate reference tissue regions thalamus, frontal cortex, and cerebellum. ctx = cortex.
MRTM allows the fast generation of voxelwise parametric BPND maps and therefore voxelwise statistical data comparison. For exploratory purposes, also the thalamus and cerebellum were considered as reference regions for SRTM and were compared with SRTM BPND and AIC values using the frontal cortex.
For the target regions putamen, caudate nucleus, ventral striatum, and substantia nigra, BPND values of the reference tissue models were compared with BPND values of the full kinetic models 1TCM, 2TCM, 2TCM[spec], 2TCM[aspec], and 1TCM[intact]. In the case of 1TCM and 2TCM, BPND was estimated as the VT ratio relative to the frontal cortex minus 1, and for 2TCM[spec], BPND was calculated as the VS over VND ratio. For 2TCM[aspec], BPND values were calculated relative to the frontal cortex while discarding the distribution volume for nonspecific binding. On the other hand, BPND values for 1TCM[intact] were determined by taking into account only the intact tracer distribution volume for target and reference tissue. In this way, the potential bias due to nonspecific binding or a brain-penetrating radiometabolite was avoided. 2TCM[aspec] BPND values were considered reference values to allow correlation analysis and to assess possible over- or underestimation of the outcome measure.
Time Stability of Outcome Measures and Test–Retest Statistics
To determine the stability of the 1TCM and 2TCM VT and SRTM and MRTM BPND estimates as functions of the acquisition time, reduced acquisition times of 90 and 60 min were evaluated and compared with a 135-min acquisition interval. For each of the reduced acquisition times, 1TCM and 2TCM AIC values were compared as well.
Test–retest variability (reproducibility) was assessed as the absolute variability VAR defined as |x1 − x2|/(x1 + x2) × 200, where x refers to the parameter of interest. Test–retest reliability was evaluated in terms of the intraclass correlation coefficient (ICC), defined as (BSMS − WSMS)/(BSMS + WSMS), where BSMS is the between-subject mean square and WSMS the within-subject mean square. The ICC ranges from 0 to 1, with values closer to 1 indicating better reliability, which means that most variance is due to between-subject rather than within-subject variation.
Statistical Analysis
Statistical analysis was performed using Prism software (version 5.0; GraphPad Software). Parameters of interest were compared using 1-way repeated-measure ANOVA including Bonferroni multiple-comparison tests. When appropriate, a 2-tailed paired Student t test was used. Correlation analysis was performed using a Spearman ρ correlation coefficient. A P value of 0.05 was considered a threshold criterion of statistical significance. Values are represented as mean ± SD unless otherwise stated. The ratio between parameters was determined as the slope of linear regression analysis with the intercept set to zero.
RESULTS
Brain Uptake and Plasma Metabolites
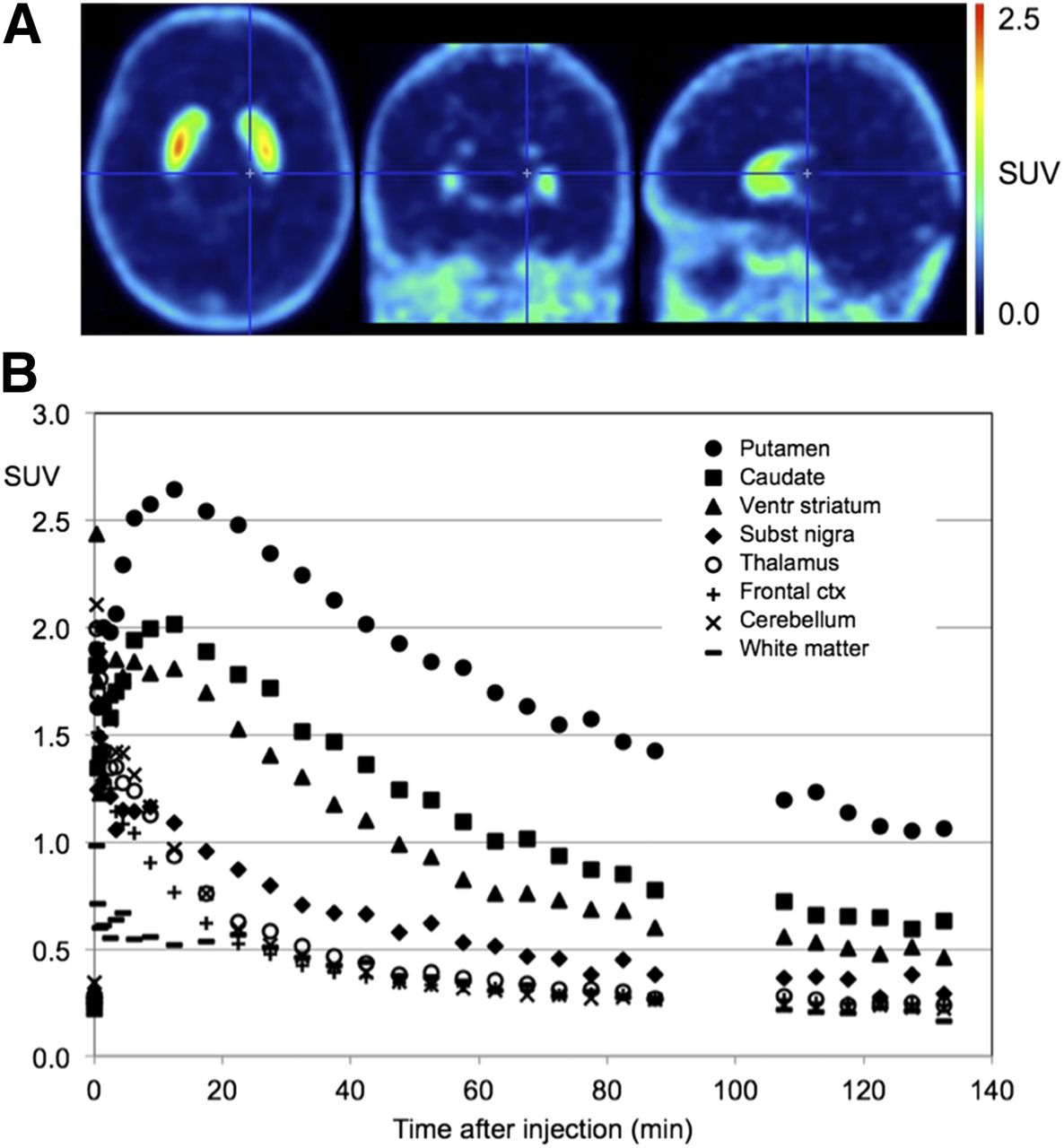
Figure 3A shows orthogonal late standardized uptake value images (averaged over 60–90 min) of 18F-JNJ-42259152 in a representative subject. Figure 3B shows the corresponding time–activity curves for the striatal regions, substantia nigra, thalamus, cerebellum, and medial frontal cortex. 18F-JNJ-42259152 exhibits relatively fast brain kinetics for the striatal regions, with a time to peak after bolus injection of 10–15 min, followed by a moderately fast washout. No subjective or objective adverse events during or after tracer administration were reported in any of the subjects.
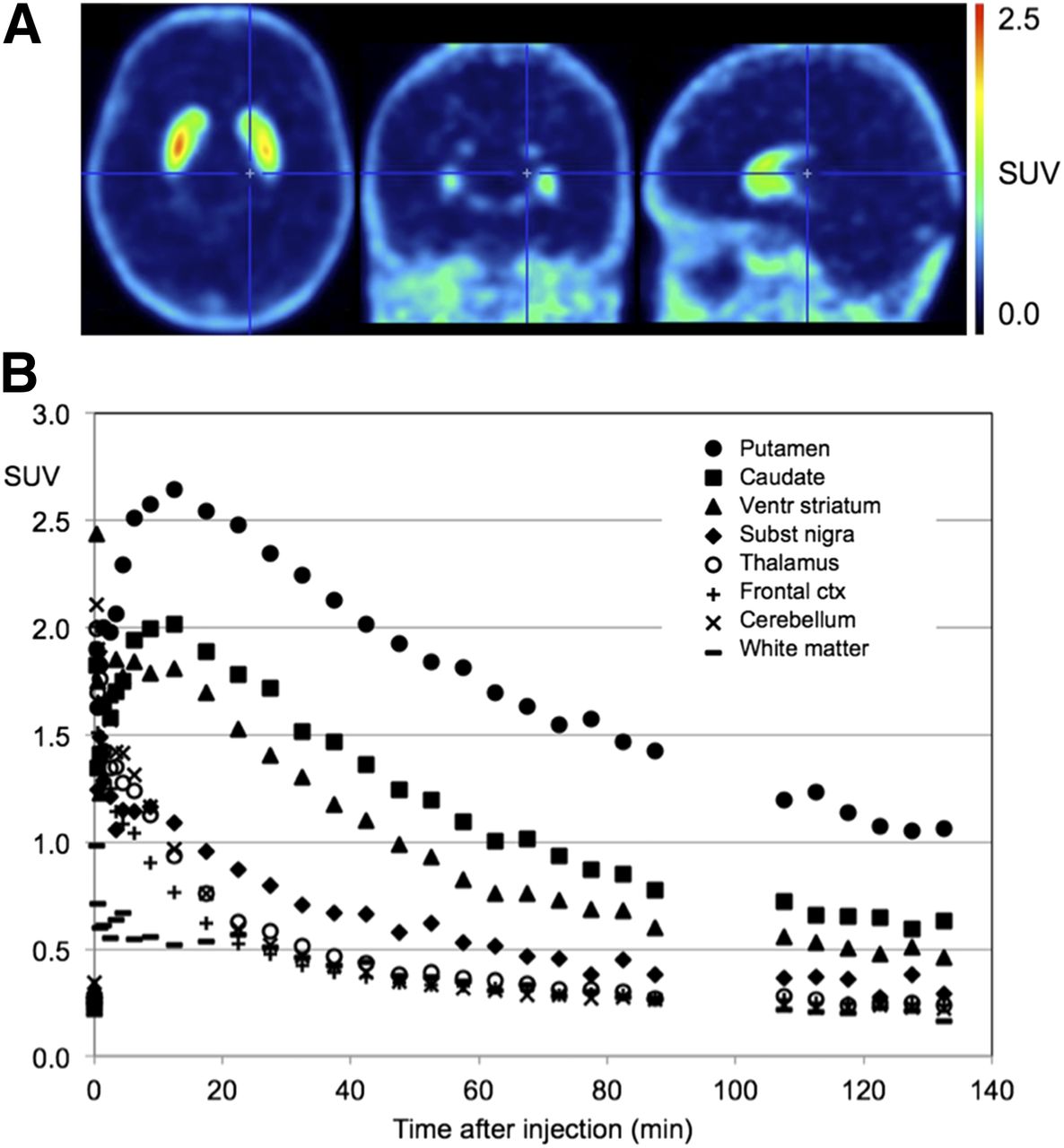
(A) Average standardized uptake value image (SUV) (averaged over 60- to 90-min interval) for 18F-JNJ-42259152 in human brain. (B) Time–activity curves for putamen, caudate nucleus, ventral striatum, substantia nigra, thalamus, frontal cortex, cerebellum, and white matter. ctx = cortex; ventr = ventral.
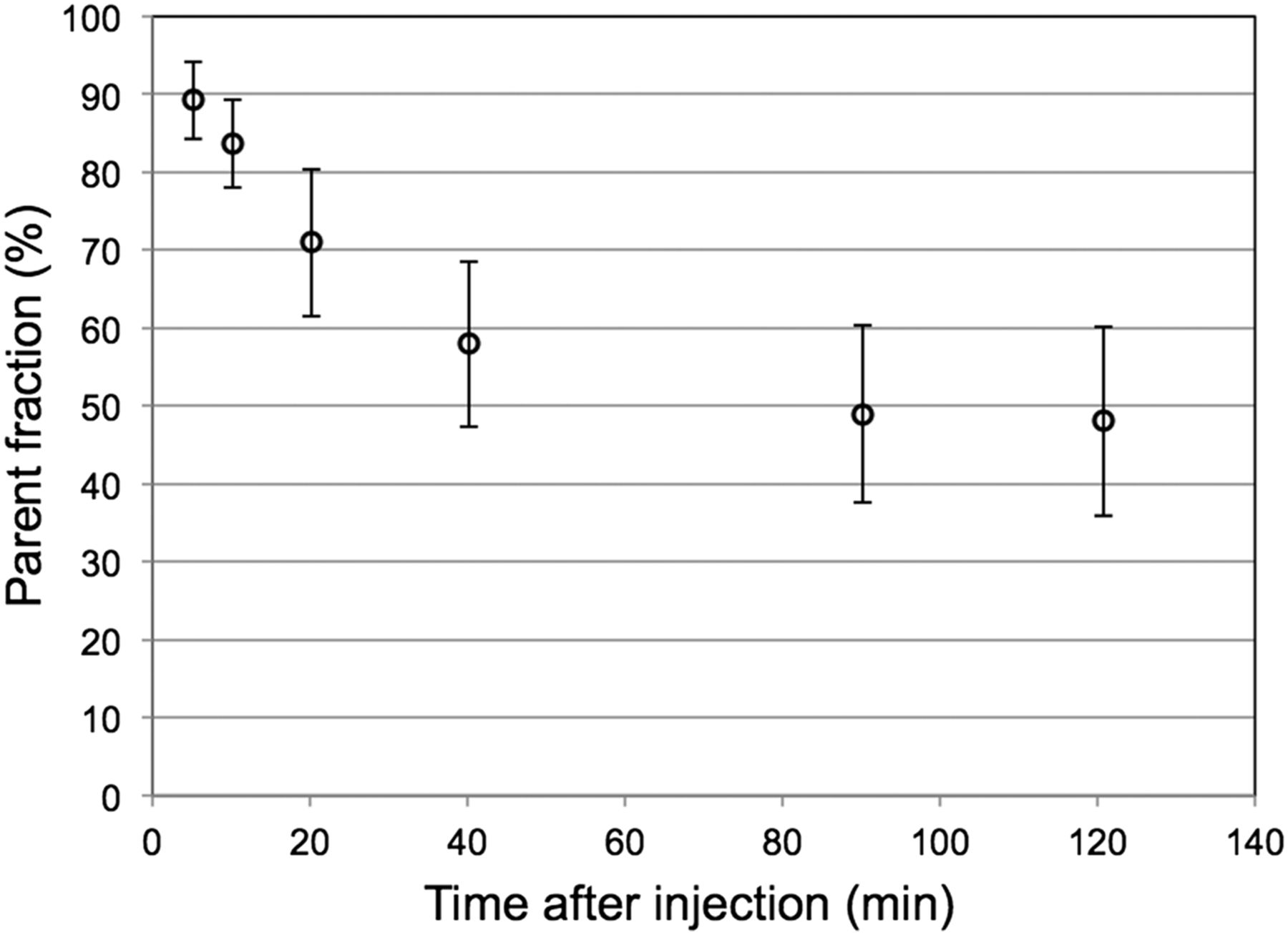
Metabolite analysis identified 2 metabolites in plasma after 40 min, 1 polar and 1 nonpolar metabolite, through cleavage of the 18F-containing pyridine ring with fenol formation, both unlikely to pass the blood–brain-barrier. The fraction of radiometabolites of 18F-JNJ-42259152 in arterial plasma, as percentage of the intact fraction, is given in Figure 4. A fraction of 51.9% ± 9.8% was metabolized, with a half-life of 18.7 ± 7.6 min constituting a fast metabolism component, whereas the remaining fraction was metabolized slowly, resulting in an approximately 50% fraction of intact tracer after 2 h.

Fraction of intact 18F-JNJ-42259152 tracer in arterial plasma as function of time after injection.
Kinetic Modeling
Tissue Distribution Volume and Metabolite Kinetics
An overview of average VT for the considered models and brain regions is given in Figure 5. The fraction nondisplaceable binding VND and specific binding VS is given for 2TCM and 2TCM[spec] whereas for 2TCM[aspec] VT is subdivided in the fraction free and specifically bound tracer VFT+S and nonspecific binding VNS. A significant difference between VT values was found for all pairs of models, except between 2TCM and 2TCM[aspec].

Obtained cerebral VT for 18F-JNJ-42259152 with 1TCM, 2TCM, 2TCM with constant nondisplaceable distribution volume VND and variable distribution volume for specific binding VS (2TCM[spec]), 2TCM with constant nonspecific distribution volume VNS and variable distribution volume for free tracer and specific binding Vft+s (2TCM[aspec]), and 1TCM taking into account brain-penetrating radiometabolite (1TCM[intact]) using 135-min acquisition time (n = 12).
When the distribution volume of the intact tracer determined using 1TCM[intact] and the distribution volume of the free and specifically bound tracer VFT+S determined using 2TCM[aspec] were compared, no significant difference was found. For 1TCM[intact], the VT,M was 0.16 ± 0.05, and VNS for 2TCM[aspec] was 0.12 ± 0.06. There was a significant correlation between the 2TCM[aspec] VNS and 1TCM[intact] VT,M values (ρ = 0.79) and between 2TCM[aspec] VNS and the fraction intact tracer A that was metabolized in the arterial blood compartment (ρ = 0.74).
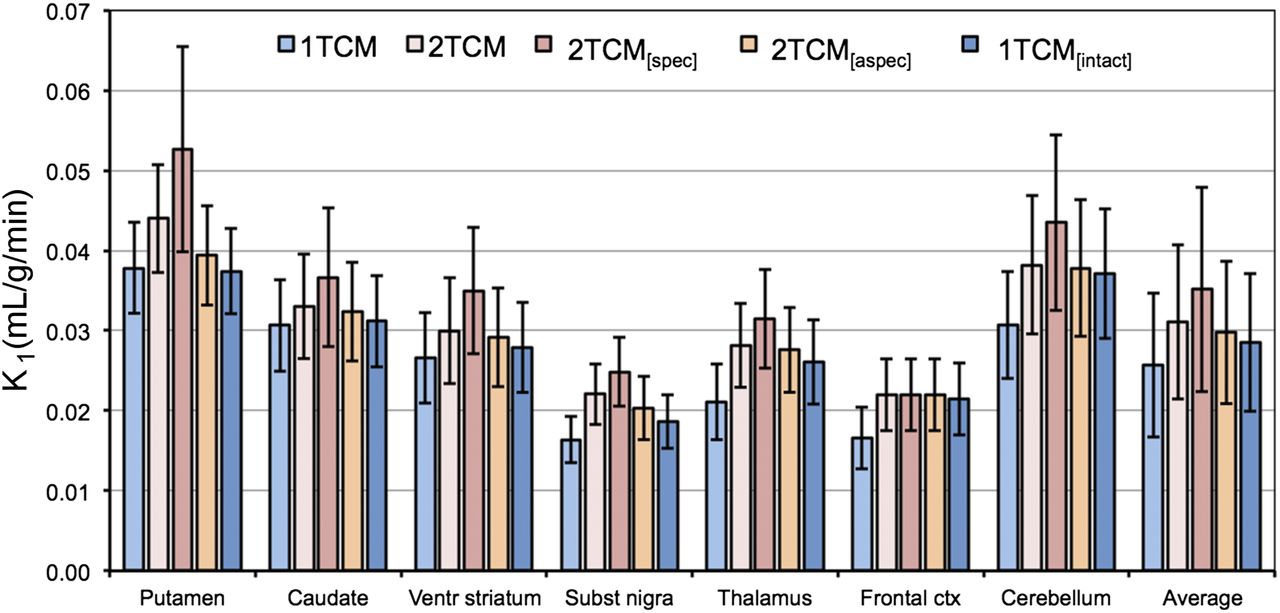
K1 values for the different models are summarized in Figure 6, and K1, k2, k3, and k4 values are presented in Table 1 for 1TCM and 2TCM. K1 values differed significantly between all model pairs, except for the pairs 2TCM and 2TCM[aspec] and 2TCM[aspec] and 1TCM[intact].
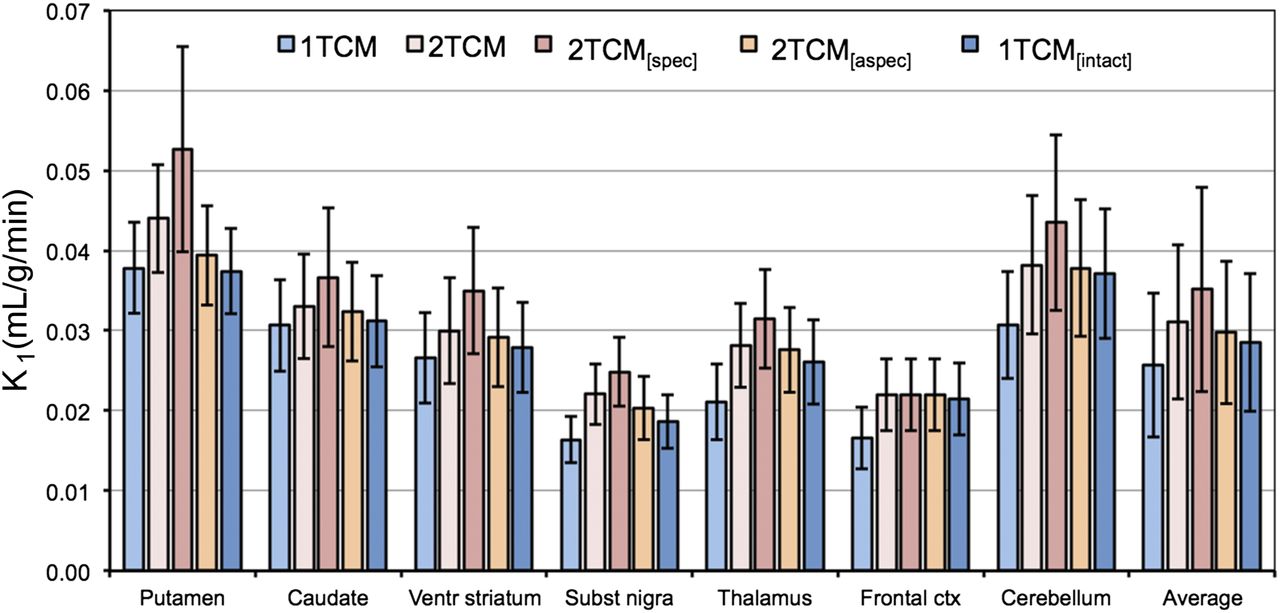
Plasma–to–tissue tracer uptake rate (K1) for 18F-JNJ-42259152 for 1TCM, 2TCM, 2TCM with constant nondisplaceable distribution volume VND and variable distribution volume for specific binding VS (2TCM[spec]), 2TCM with constant nonspecific distribution volume VNS and variable distribution volume for free tracer and specific binding VFT+S (2TCM[aspec]), and 1TCM taking into account brain-penetrating radiometabolite (1TCM[intact]) using 135-min acquisition time (n = 12).
Overview of Rate Constants of 1TCM and 2TCM for 18F-JNJ-42259152 and Different Brain Regions
Average AIC values for 1TCM, 2TCM, 2TCM[spec], and 2TCM[aspec] were, respectively, 261.1 ± 15.0, 243.7 ± 21.0, 249.9 ± 21.4, and 245.7 ± 23.9. The AIC for the 1TCM was significantly higher than for the 2TCM. Furthermore, significant differences were found between the 2TCMs except between 2TCM and 2TCM[aspec].
Table 2 gives an overview of the correlations between 2TCM VT and other models per brain region. The global VT ratios are given in Table 3. For all target regions, the correlation between models was excellent.
Correlation Coefficients for Different Models for 18F-JNJ-42259152 Versus 2TCM
Global Ratio of VT and BPND for 18F-JNJ-42259152
BPND and Reference Tissue Models
An overview of average BPND for the considered models and brain target regions is given in Figure 7. No significant BPND differences were found among 2TCM, SRTM, and MRTM or between 2TCM[aspec] and 1TCM[intact]. All other BPND differences were significant. Table 2 summarizes the Spearman correlation coefficients separately for each target region, and Table 3 gives overall BPND ratios, both using 2TCM[aspec] BPND as a reference value.
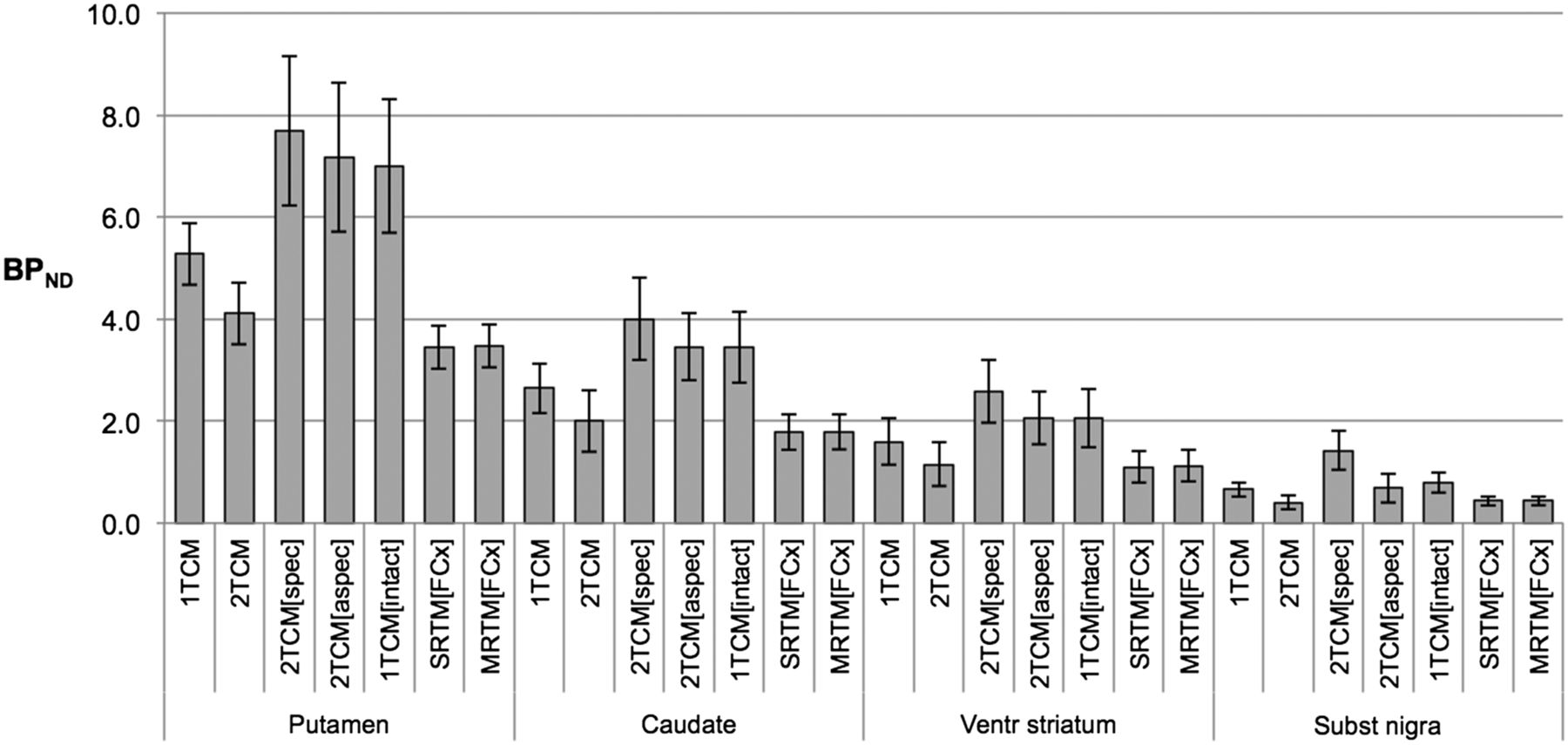
BPND for 18F-JNJ-42259152 for target regions putamen, caudate nucleus, ventral striatum, and substantia nigra, using frontal cortex as reference region. For 1TCM and 2TCM, BPND was estimated as ratio minus 1. For 2TCM with constant nondisplaceable distribution volume VND and variable distribution volume for specific binding VS (2TCM[spec]), BPND was calculated ratio of VS over VND. For 2TCM with constant nonspecific distribution volume VND and variable distribution volume for free tracer and specific binding VFT+S (2TCM[aspec]), BPND values were calculated discarding nonspecific binding. BPND values for 1TCM taking into account brain-penetrating radiometabolite (1TCM[intact]) consider only intact tracer distribution volume. BPND for SRTM and MRTM are reported as well. All values are estimated for 135-min acquisition time (n = 12).
When the thalamus and cerebellum were considered as reference tissues for SRTM, average BPND values for the putamen, caudate nucleus, ventral striatum, and substantia nigra were, respectively, 2.78 ± 0.47, 1.35 ± 0.31, 0.77 ± 0.23, and 0.22 ± 0.10 for the thalamus and, respectively, 2.91 ± 0.44, 1.43 ± 0.36, 0.84 ± 0.34, and 0.26 ± 0.12 for the cerebellum. Compared with the corresponding values 3.45 ± 0.43, 1.78 ± 0.35, 1.10 ± 0.31, and 0.44 ± 0.09 for the frontal cortex, SRTM BPND values referenced to the thalamus and cerebellum both differed significantly from those referenced to the frontal cortex whereas no significant difference was found between the thalamus and cerebellum. SRTM BPND values using the thalamus and cerebellum relate to SRTM BPND values using the frontal cortex by a factor 0.79 and 0.83, respectively. In terms of correlation with SRTM BPND using the frontal cortex, spearman ρ correlation coefficients of SRTM BPND using thalamus and cerebellum were, respectively, 0.83 and 0.76 for the putamen, 0.92 and 0.80 for the caudate nucleus, 0.81 and 0.87 for the ventral striatum, and 0.64 and 0.52 for the substantia nigra. Overall correlation coefficient for both the thalamus and the cerebellum was 0.98. All correlations were significant except the SRTM BPND for the substantia nigra using the cerebellum. To compare reference regions in terms of model fitting, SRTM AIC values using the thalamus, frontal cortex, and cerebellum were, respectively, 239.1 ± 17.4, 240.5 ± 16.7, and 240.8 ± 16.3 and not significantly different.
Time Stability and Test–Retest of VT and BPND
The time stability of VT for 1TCM and 2TCM and of BPND for SRTM and MRTM are given in Figure 8. 1TCM and 2TCM VT values for the 135- and 90-min acquisition times were not significantly different contrary to 60-min VT values, which were significantly lower than those for the 135-min acquisition time. 2TCM was the preferred model for the 135-min acquisition time, with a significantly lower AIC and significantly higher VT than 1TCM. Although 2TCM AIC (204.0 ± 19.3) is still significantly lower than 1TCM AIC (212.7 ± 12.1) for the 90-min acquisition time, the situation is reversed for the 60-min acquisition time where—based on the lower AIC—1TCM (64.0 ± 15.4) is the preferred model over 2TCM (69.9 ± 15.5).
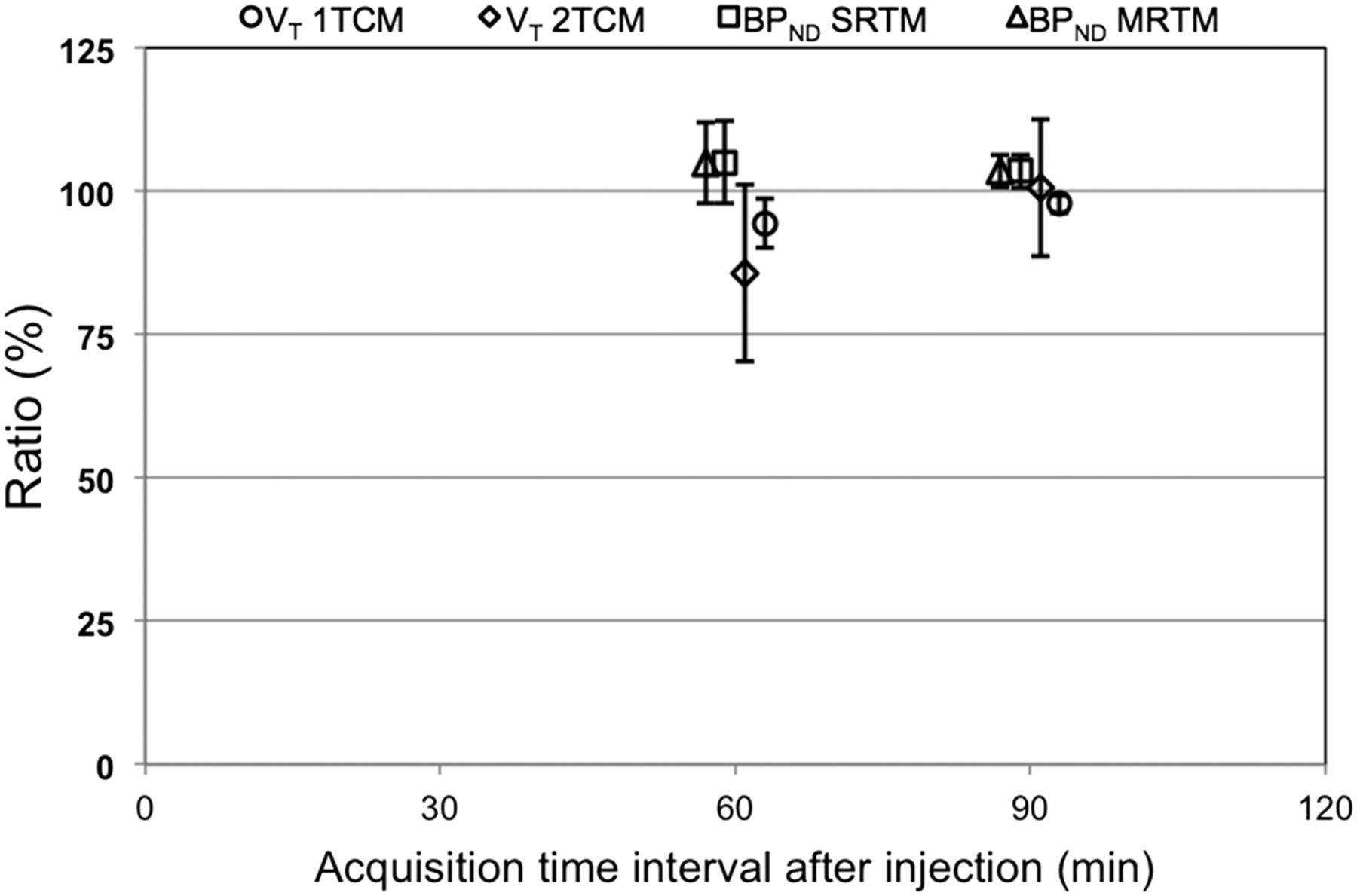
Induced bias and variance of reduced acquisition times of 60 and 90 min for 18F-JNJ-42259152, relative to acquisition time of 135 min. This is shown both for VT of 1TCM and 2TCM and for BPND values of SRTM and MRTM using frontal cortex as reference tissue.
For MRTM and SRTM, BPND values were slightly but significantly higher for the 90- and 60-min acquisition times than for the 135-min acquisition time. However, global correlations between BPND values for the shorter times and the 135-min acquisition time were significant and excellent for both reference tissue models (Spearman ρ ≥ 0.94).
Region-specific test–retest statistics for VT and BPND are summarized in Table 4. No significant differences between the VT values of different compartmental models were found in terms of variability or ICC. In addition, no significantly different variability or ICCs were found for BPND values determined with SRTM and MRTM.
Test–Retest Statistics for 18F-JNJ-42259152
DISCUSSION
The regional in vivo distribution of 18F-JNJ-42259152 in the brain was concordant with the known distribution of PDE10A from preclinical and histopathologic findings, with the highest activity in the putamen and in second order the caudate nucleus, ventral striatum, and substantia nigra. Cortical and cerebellar activity was more than 10-fold lower than activity in the striatum. Previous studies using in situ hybridization and quantitative polymerase chain reaction have shown that levels in the cortex, cerebellum, and thalamus are 10- to 40-fold lower than in the striatum (1,10). This finding indicates that, although expression levels outside the nigrostriatal region are low, they are not negligible and a careful evaluation of their use as a reference region is therefore warranted.
Tracer kinetics were first assessed using a standard 1TCM and 2TCM. Our findings indicated that the 2TCM is the preferred method to describe tracer kinetics, both for target and for potential reference regions. To determine whether 2TCM was more appropriate for potential reference regions because of low PDE10A expression in these regions, we constrained the distribution volume for nondisplaceable binding VND for all brain regions to a constant value determined using the frontal cortex. In this way, VT differences between regions could be induced only by differences in specific binding. However, VT values of this constrained 2TCM[spec] approach were significantly different from unconstrained 2TCM fittings. Second, we assumed the presence of a nonspecific binding tissue compartment with slower tracer kinetics next to the free and specific binding tissue compartment with distribution volume VFT+S. Distribution volume for nonspecific binding VNS was estimated using a frontal cortex time–activity curve and constrained to this value for the other brain regions. The VT values of this constrained 2TCM[aspec] approach were not significantly different from unconstrained 2TCM fittings. Moreover, AIC values of the 2TCM[aspec] fittings were not significantly different from those of the 2TCM fittings. To determine whether this extra compartment was needed to model the nonspecific binding of a potential brain-penetrating radiometabolite instead of the intact tracer, we also modeled intact tracer kinetics with metabolite kinetics for the target and reference regions using 2 separate parallel 1-tissue compartments with 2 separate arterial input functions (i.e., intact tracer and radiometabolite input function [1TCM[intact]]). For this approach, the tissue VT,M was estimated using the frontal cortex and was constrained to this value for the other brain regions, to be able to estimate the distribution volume of the intact tracer for each brain region. No significant difference was observed between intact tracer VT and VF+S. Furthermore, significant correlations were found between VT,M and VNS and between VNS and the fraction of metabolites in the arterial plasma compartment. Because there was evidence from preclinical experiments of a brain-penetrating radiometabolite in rodents (17), the current findings can be compatible also with a radiometabolite that is entering the brain, which could be a confounding factor for tracer quantification. Nevertheless, these findings also proved that both approaches were essentially the same and that metabolite kinetics could be modeled as nonspecific binding using only an intact tracer arterial input function.
Although possibly biased by a brain-penetrating radiometabolite, significant VT correlations were found with the standard 1TCM and 2TCM approach for all regions. For the target regions, correlations were still significant between VT using a 2TCM approach and VT of the intact tracer, demonstrating that in any case there is a reproducible bias of the tentative radiometabolite on tracer quantification.
Plasma–to–tissue uptake rates K1 (Table 2) are relatively low, with significantly lower values due to poorer fittings for the 1TCM than 2TCM approach. These K1 values are comparable to tracers such as 18F-fluorodopa (population average K1, 0.03 mL/g/min) (21), meaning that tracer uptake is perfusion independent. Although in general this can have the possible disadvantage of low signal because of the limited tracer uptake, the current spatial restriction to the striatum, with excellent signal-to-background ratio, indicates that an accurate quantification is feasible.
Because 2TCM[aspec] accounts for nonspecific binding of either intact tracer or radiometabolite and provided similar VT and AIC values as unconstrained 2TCM fittings, BPND calculated as the ratio VFT+S minus 1 relative to the frontal cortex was used as a reference value. As expected, no significant difference was found between these BPND 2TCM[aspec] reference values and BPND calculated as the 1TCM[intact] tracer VT ratio minus 1 relative to the frontal cortex. Although SRTM and MRTM BPND values were significantly lower than the reference values because the radiometabolite uptake or nonspecific binding is included when determining the reference tissue BPND values, SRTM and MRTM BPND showed good correlations with the 2TCM[aspec] reference values for the different target regions. Only the SRTM BPND correlation for the putamen was borderline not significant, with a P value of 0.055.
Concerning the use of the thalamus and cerebellum as reference tissues for SRTM, both regions proved to be acceptable alternatives, with BPND values that correlated significantly with those for the frontal cortex (except for the substantia nigra), although BPND values were significantly different.
For 1TCM, VT was stable down to 60 min, with a significant negative bias of 5.5%. For 2TCM, a more pronounced significant negative bias of 14% was noticed. 2TCM fittings became less stable for reduced acquisition times whereas for the 60-min acquisition time, differences between 1TCM and 2TCM VT became insignificant and 1TCM was the preferred model with the lower AIC. These findings are in agreement with the assumption that 2TCM accounts for a compartment with slow kinetics representing either nonspecific tracer binding or radiometabolite brain uptake.
When the small positive bias for SRTM and MRTM (both 4% for 90 min and 5% for 60 min) is taken into account, we can conclude that 60- to 90-min dynamic acquisition should be sufficient to allow accurate quantification with SRTM or MRTM using the frontal cortex as a reference tissue.
In terms of long-term (weeks) test–retest, all models presented similar statistics, with repeatability values of 5%–20% for VT and 5%–12% for BPND. Although ICC values were within acceptable range, overall variability was relatively high and should be considered when comparing intersubject VT and BPND values. ICC values were lower for the reference tissue models than for the compartmental models because of the lower between-subject variability. Variability was higher for compartmental models than for reference tissue models, probably because of added variability induced by arterial sampling and radiometabolite fraction corrections.
Concerning the limitations of the study and possible future perspectives, it needs to be emphasized that in the current design there was no independent reference available to provide an objective assessment of quantitative accuracy of the all proposed kinetic models. However, 3 different scenarios were considered: a brain-penetrating metabolite, nonspecific binding with slower kinetics, and a fraction of specific binding (because low nonzero PDE10A expression has been documented in histologic postmortem studies) in candidate reference regions such as the frontal cortex. Independent of the real underlying physiologic phenomenon, the kinetic models that take into account these different scenarios provide highly correlated quantitative endpoints that are also highly correlated with the standard and more straightforward reference tissue models. Therefore, future PET dose–occupancy studies (enzyme-blocking studies) are needed to allow a better estimate of the nondisplaceable binding of the tracer and to confirm or exclude one of the before-mentioned scenarios. PET dose–occupancy studies will also provide a better insight into the regional and interindividual differences of nondisplaceable binding and justify use of reference tissue models as quantitative models. Furthermore, the small extraction fraction of the tracer with high affinity and accumulation in the striatum might cause some concern that delivery could be rate-limiting. This issue needs to be addressed in future studies and by comparison to other radioligands that are currently under development.
CONCLUSION
We successfully characterized kinetic modeling of the PDE10A tracer 18F-JNJ-42259152 in the human brain. 18F-JNJ-42259152 currently is the only published PDE10A PET tracer with good characteristics for human studies. A potential metabolite contribution in brain uptake cannot be excluded, although nonspecific binding of the intact tracer can be an alternative explanation for the observed kinetic behavior. In any case, this contribution is small and reproducible. Therefore, a 90-min dynamic acquisition and a reference tissue approach with the frontal cortex as reference tissue provides a reproducible and accurate estimation of BPND, allowing more feasible quantitative studies in neuropsychiatric study populations.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. Koen Van Laere is Senior Clinical Fellow for the Flemish Science Foundation (FWO), Belgium. Kristof Dubois and Mark E. Schmidt are employees of Janssen Pharmaceutica R&D, Belgium. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Kwinten Porters and Mieke Steukers and the clinical PET radiopharmacy team for their skilled help. Janssen Pharmaceutica is gratefully acknowledged for the supply of the precursor and reference molecule.
Footnotes
Published online Jul. 10, 2013.
- © 2013 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication December 17, 2012.
- Accepted for publication March 26, 2013.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Discovery and Development of 11C-Lu AE92686 as a Radioligand for PET Imaging of Phosphodiesterase10A in the Brain
- In Vivo Assessment and Dosimetry of 2 Novel PDE10A PET Radiotracers in Humans: 18F-MNI-659 and 18F-MNI-654
- Phosphodiesterase 10A PET Radioligand Development Program: From Pig to Human
- PET imaging shows loss of striatal PDE10A in patients with Huntington disease