


Abstract
Peptide-receptor radionuclide therapy (PRRT) with radiolabeled somatostatin analogs such as octreotide is an effective therapy against neuroendocrine tumors. Other radiolabeled peptides and antibody fragments are under investigation. Most of these compounds are cleared through the kidneys and reabsorbed and partially retained in the proximal tubules, causing dose-limiting nephrotoxicity. An overview of renal handling of radiolabeled peptides and resulting nephrotoxicity is presented, and strategies to reduce nephrotoxicity are discussed. Modification of size, charge, or structure of radiolabeled peptides can alter glomerular filtration and tubular reabsorption. Coinfusion of competitive inhibitors of reabsorption also interferes with the interaction of peptides with renal endocytic receptors; coinfusion of basic amino acids is currently used for kidney protection in clinical PRRT. Furthermore, nephrotoxicity may be reduced by dose fractionation, use of radioprotectors, or use of mitigating agents. Decreasing the risk of nephrotoxicity allows for administration of higher radiation doses, increasing the effectiveness of PRRT.
The potential to irradiate tumors using radiolabeled antibodies that target tumor-associated epitopes was first reported in the 1950s. A drawback of conventional radioimmunotherapy is the slow extravasation and clearance of intact antibodies from the blood due to their size (∼150 kDa). The optimal tumor-to-background ratio is reached only after several days, and the radiosensitive bone marrow receives a considerable radiation dose. Radiolabeled antibody fragments and analogs, such as Fab fragments and Affibody molecules, were developed to improve pharmacokinetics and increase tumor–to–normal tissue dose ratios.
An alternative approach to tumor targeting is provided by the overexpression of specific cell-surface receptors by certain tumors. For example, somatostatin receptors can be targeted by somatostatin analogs such as octreotide. The use of radiolabeled somatostatin analogs for imaging and therapy of neuroendocrine tumors was first studied in the 1990s (1). Nowadays, peptide-receptor radionuclide therapy (PRRT) with somatostatin analogs has an established role in the therapy of metastasized neuroendocrine tumors (2). Many other radiolabeled peptides have been studied, such as gastrin and cholecystokinin analogs, which target the cholecystokinin-2 or gastrin receptor, and exendin, which targets the glucagonlike peptide receptor (3,4).
Hydrophilic radiolabeled peptides, including small antibody fragments, are excreted mainly via the kidneys and are partly reabsorbed in the proximal tubular cells. Renal retention of therapeutic radionuclides causes a relatively high radiation dose to the kidneys, which can lead to kidney failure (1,5). This toxicity precludes the use of higher doses, thus limiting the efficacy of therapy.
Insight into the mechanisms of renal retention and toxicity of radiolabeled peptides is necessary for the study, design, and use of strategies to reduce nephrotoxicity in PRRT and radioimmunotherapy. Several such strategies have already been studied, including structural modification of radiolabeled peptides, competitive inhibition of their renal reabsorption, and use of radioprotectors. Reduction of nephrotoxicity may enable the administration of higher-activity doses, thus improving the efficacy of therapy.
This article presents an overview of the mechanisms of renal handling, retention, and toxicity of radiolabeled peptides and antibody fragments and discusses strategies to reduce nephrotoxicity.
PHYSIOLOGY OF RENAL HANDLING AND RETENTION OF RADIOLABELED PEPTIDES
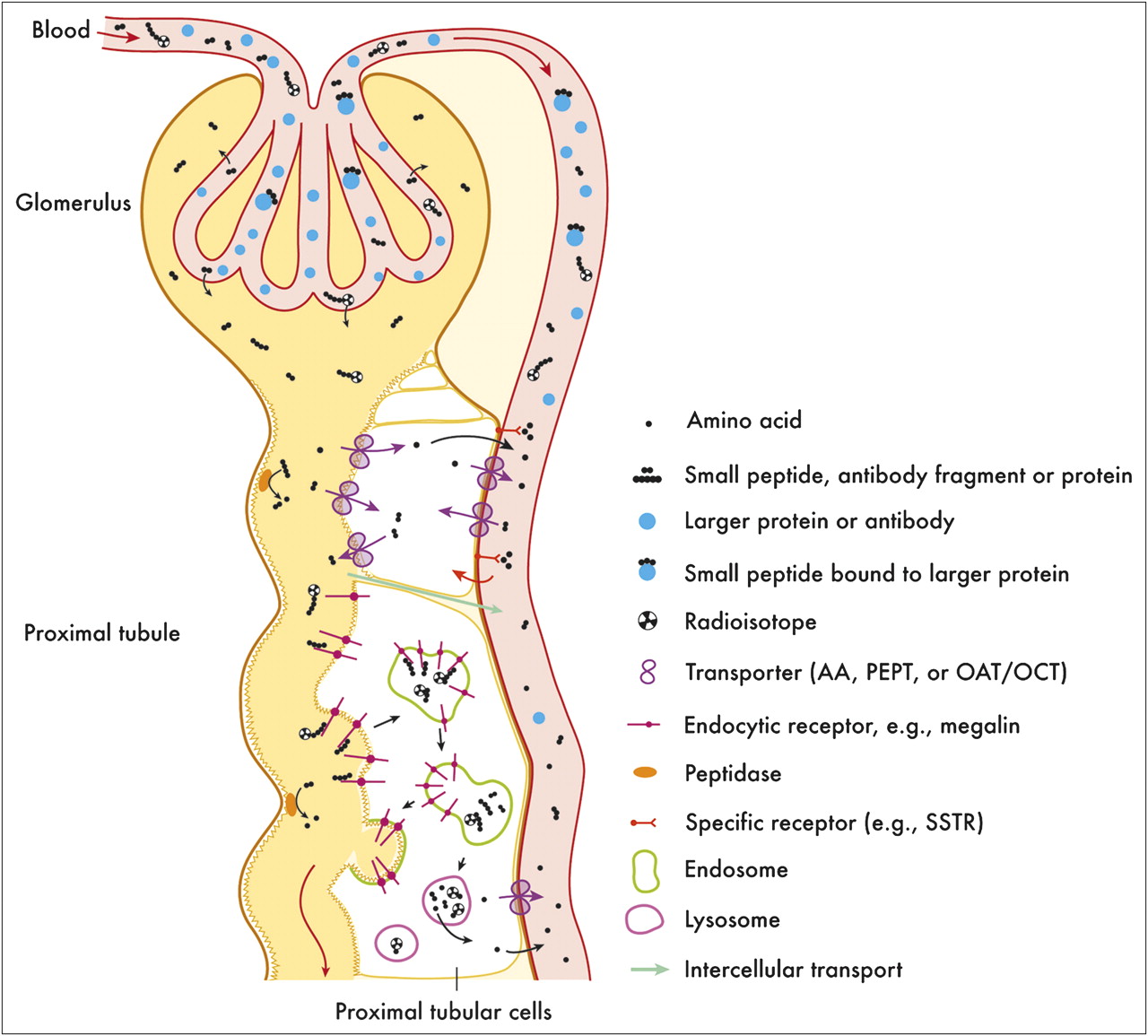
In the human kidneys, approximately 180 L of plasma are filtered through the glomeruli each day, forming the ultrafiltrate that enters the lumen of the proximal tubules. About 99% of the filtered water and a large part of the solutes is actively reabsorbed from the ultrafiltrate and mostly transported back into the bloodstream, to prevent the excessive loss of water and useful nutrients and to preserve homeostasis. The major part of the filtered load of proteins and peptides is reabsorbed in the proximal tubules. Other solutes are actively cleared from the blood into the ultrafiltrate by tubular secretion (6). Figure 1 presents an overview of glomerular and proximal tubular handling of peptides and proteins.

Overview of glomerular and proximal tubular handling of proteins and peptides. AA = amino acid transporter; OCT = organic cation transporter; PEPT = oligopeptide transporter.
Glomerular Ultrafiltration
The degree of filtration of solutes is largely dependent on their size, charge, and protein binding. Molecules with an effective radius of less than 1.8 nm (or weighing <∼12 kDa) are freely filtered through the pores in the glomerular membrane, whereas molecules larger than 4.2 nm (weighing >70 kDa) are not filtered or are filtered to only a very limited extent. Small molecules that are bound to larger transport proteins such as albumin will not pass the glomerular filter. Evidence indicates that cationic and neutral molecules are filtered more efficiently than anionic molecules (6), although this concept of charge selectivity has been challenged in recent years (7). Most radiolabeled peptides are not bound to proteins and weigh less than 12 kDa; consequently, they are cleared efficiently via glomerular filtration.
Proximal Tubular Reabsorption and Processing
In the proximal tubule, approximately 50%−60% of filtered water and solutes is reabsorbed by active and passive transport processes. The main driving force for these processes is the Na+-K+-adenosine triphosphatase in the basolateral membrane, which pumps Na+ out of the cell and K+ into the cell. The ensuing concentration gradient stimulates reentry of Na+ through various symporters and antiporters, providing energy for the cotransport of other molecules among which most are amino acids (6).
Multiple transport mechanisms are involved in the proximal tubular reabsorption of proteins and peptides. Some oligopeptides are hydrolyzed at the brush border of proximal tubular cells, after which the resulting amino acids and di- or tripeptides are transported into the cells by amino acid transporters and the oligopeptide transporters, respectively (6). Larger peptides and proteins are internalized via endocytosis. Most peptides are almost quantitatively reabsorbed in the proximal tubules.
Receptor-Mediated Endocytosis
Endocytosis is triggered by the binding of ligands to endocytic receptors, providing a positive feedback mechanism that accelerates endocytosis in response to an increase in the amount of filtered proteins. After binding of ligands to endocytic receptors, the plasma membrane invaginates, forming clathrin-coated pits and early endosomes. Early endosomes fuse with late endosomes, in which the acidic pH causes dissociation of ligands from receptors. The receptors concentrate on 1 side of the endosomal membrane, which buds off and recycles to the cell membrane. The late endosomes fuse with lysosomes, in which the ligands are catabolized by proteolytic enzymes. The cytoskeleton plays an important role in the intracellular trafficking of these vesicles (6).
Megalin and cubilin, large endocytic receptors, play an important role in the proximal tubular reabsorption of various structurally different proteins, peptides, drugs and other molecules, including albumin, apolipoproteins, hemoglobin, gentamycin, and many others (8). Megalin and cubilin are expressed exclusively on the apical membrane of proximal tubular cells (8). The cubilin receptor does not possess an intracellular signaling domain, and evidence suggests that expression and endocytosis of the cubilin receptor are dependent on interactions with megalin and amnionless, another transmembrane protein (9,10). Several ligands have been reported to bind both to megalin and to cubilin (8). In vitro studies have indicated that radiolabeled somatostatin analogs are actively reabsorbed in proximal tubular cells by receptor-mediated endocytosis, probably involving megalin or cubilin (11). Autoradiography studies of rodent kidneys showed that various radiolabeled peptides accumulate mainly in the renal cortex (12). De Jong et al. reported that the renal uptake of 111In-octreotide was significantly decreased in kidney-specific megalin-deficient mice (13), and our group recently found that renal uptake of 111In-labeled octreotate, minigastrin, exendin, and neurotensin was also decreased by 40%−70% in these megalin-deficient mice (14), implying that megalin or megalin-dependent receptors such as cubilin are indeed involved in the proximal tubular reabsorption of these radiolabeled peptides.
Cationic peptides are generally reabsorbed more efficiently by the proximal tubules than are neutral and anionic peptides. This difference is thought to be caused by preferential binding of cationic groups to anionic binding sites on the brush border membrane. However, the highly anionic peptide minigastrin also expresses high renal uptake (3), and megalin ligands can be cationic or anionic. This indicates that other ligand properties, such as structure and charge distribution, also determine reabsorption.
Role of Ligand-Specific Receptors
Particular peptides may also be taken up by ligand-specific receptors, such as somatostatin receptors (15). Abundant expression of all somatostatin receptor subtypes has been described in the human and murine kidney (16,17), and subtypes 3 and 4 were found in the rat kidney (18). Rolleman et al. retrospectively studied 111In-octreotide scans of patients who were scanned before and during treatment with unlabeled octreotide. A mean 18% reduction in kidney uptake during octreotide treatment was found, suggesting some involvement of somatostatin receptors in the renal uptake of 111In-octreotide (19). However, an excess of unlabeled octreotide may also interfere with binding of 111In-octreotide to other receptors such as megalin. Expression of cholecystokinin 2 and glucagonlike peptide 1 receptors on proximal tubular cells was described (20,21), and many other ligand-specific receptors may be expressed in the kidneys. The importance of these receptors for the renal uptake of radiolabeled peptides remains to be determined.
Other Pathways for Reabsorption
Fluid-phase endocytosis (pinocytosis) is likely to play a role in the proximal tubular reabsorption of proteins and peptides. However, the capacity of this pathway is reported to be much lower than the capacity of receptor-mediated endocytosis (22). For hydrophobic molecules, passive diffusion across the luminal membrane may play an additional role.
Part of the transport of water and electrolytes occurs via the intercellular pathway, through pores in the tight junctions between cells and through the interstitium. It is unknown whether small peptides may be reabsorbed through this route. Recently, a transcellular pathway for reabsorption of albumin has been proposed, consisting of large endocytic vesicles that are transported rapidly from the apical to the basolateral membrane of proximal tubular cells (7). This route may also play a role in the reabsorption of radiolabeled peptides, but its existence remains controversial (23). In any case, peptides and proteins transported via these routes are thought to be returned to the circulation intact and are not retained in the kidneys. Therefore, these pathways are unlikely to contribute to nephrotoxicity significantly.
Peritubular Absorption and Tubular Secretion
Many endogenous and exogenous substances, such as hormones and drugs, are actively secreted from the blood into the lumen of the proximal tubules by the processes of peritubular absorption and tubular secretion. Several transporters involved in these energy-requiring processes have been identified, such as the organic anion transporters (OATs) and organic cation transporter families, oligopeptide transporters, and the multidrug resistance proteins (6). Different transporters are expressed on the basolateral and on the apical membranes of the proximal tubular cells, and some are also involved in the reabsorption of substances from the tubular lumen. The substrates transported by these pathways are relatively small. The tripeptide 99mTc-mercaptoacetyltriglycine is excreted mainly by tubular secretion, and Trejtnar et al. reported that three 99mTc-labeled tetrapeptides were secreted in the proximal tubules (24). A study suggests that OATs may be involved in the tubular uptake of 111In-octreotide, either from the lumen or from the plasma (25).
Intracellular Catabolism and Retention
Peptides and proteins that undergo endocytosis in the proximal tubules are subsequently catabolized into amino acids in lysosomes. These amino acids are transported or diffuse out of the lysosomes into the cytoplasm and return to the circulation. Monoiodotyrosine, the primary radiolabeled catabolite from iodinated antibodies, is indeed released quickly from cells (26). Other radiolabeled catabolites, such as lysine–chelate conjugates, N-terminal amino acid chelate conjugates (27), and specifically engineered iodinated products (28), cannot cross the lysosomal membrane. These products, which are trapped in the lysosomes, are referred to as residualizing radiolabels. Both for scintigraphy and for PRRT, the entrapment of radiolabels is a desirable effect in target cells, enhancing the tumor-to-background ratio. However, this entrapment also contributes to the relatively high retention of radioactivity in the kidneys.
Oligopeptides that are taken up by transporters such as OATs or organic cation transporters can leave the cell via similar transporters or by diffusion or can be processed further in the cytosol or in lysosomes (6). The contribution of these transporters to the renal retention of radiolabeled peptides remains to be investigated.
KIDNEY TOXICITY
The bone marrow is the dose-limiting organ in most types of radionuclide therapy, since its rapidly dividing cells are sensitive to radiation. Acute myelodepression can occur in the first days to weeks after treatment and can be managed by supportive measures or by bone marrow transplantation in the case of myeloablative therapy. Up to years after therapy, late myelotoxicity such as myelodysplastic syndrome and leukemia can develop, which may also be treated with bone marrow transplantation (29–31). However, the kidneys are sensitive to radiation as well, and given the relatively high renal retention of many radiolabeled peptides, the kidneys are often the dose-limiting organs in PRRT (5,32,33).
Nephropathy After External-Beam Radiation Therapy (EBRT)
Radiation nephropathy caused by EBRT has been described extensively (34,35). The only signs of radiation nephropathy may be solitary hypertension or asymptomatic proteinuria. Acute radiation nephropathy is defined as signs of kidney failure (proteinuria, hypertension, heart failure, azotemia, and anemia) developing within 6–12 mo after irradiation. Pathologic kidney changes include atrophy, tubulointerstitial scarring, mesangiolysis, and, in severe cases, thrombotic microangiopathy. Acute radiation nephropathy can be reversible or progress to chronic nephropathy. Chronic radiation nephropathy can also develop up to many years after irradiation and is characterized by loss of kidney mass and function (36). In animal models, early kidney damage developing in the first days to weeks after irradiation has been described, including damage to glomeruli, tubuli, and vasculature (35). The initial radiation-induced damage is DNA damage, which is caused mostly indirectly through the formation of reactive oxygen species and can lead to cell death. Subsequent tissue reactions to this damage, mediated in part by the renin-angiotensin-aldosterone system, are involved in the development of chronic damage (36).
Nephropathy After Radionuclide Therapy
Data about nephrotoxicity resulting from radionuclide therapy are limited. In mice, acute radiation nephritis with uremia and pronounced tubular damage developing in the first weeks after administration of 90Y-labeled anti–carcinoembryonic antigen Fab fragments (9–15 MBq) and 225Ac-labeled antibodies (13 kBq) has been described (37,38). Renal damage in rats was described 3–9 mo after injection of 278–555 MBq of [177Lu-DOTA0,Tyr3]octreotate (177Lu-DOTATATE) (39,40). In patients, acute radiation nephropathy developing 0.5–6 mo after treatment with more than 7.4 GBq of [90Y-DOTA0-Tyr3]octreotide (90Y-DOTATOC) per square meter has been described (1,41), as well as chronic radiation nephropathy after PRRT with 90Y-labeled somatostatin or gastrin analogs (32,42).
In EBRT, kidney dose limits of 15–17 Gy in 2-Gy fractions are considered safe (43). Doses of 23 Gy cause chronic kidney failure in approximately 5% of patients within 5 y (44). However, the reaction of tissue to radiation does not depend only on the total absorbed dose but also on dose rate, fractionation, microdistribution of dose within the organ, and radiation type (α-, β-, γ-, or x-radiation) and energy (29,33,43). Typically, in PRRT, dose rates are much lower than in EBRT, distribution within the organ is more inhomogeneous, and the radiation has a shorter range of penetration. Therefore, maximum tolerable dose limits for PRRT may differ significantly from those for EBRT. Use of the biological effective dose, which takes the effects of dose fractionation and sensitivity of the organ to radiation into account, may be more appropriate for setting dose limits than is the absorbed dose. Barone et al. described a group of 18 patients treated with 90Y-DOTATOC and found that a kidney biological effective dose of more than 45 Gy increased the risk of rapid deterioration of renal function after PRRT (45). Bodei et al. studied dosimetry in 28 patients receiving PRRT with 90Y-DOTATOC or 177Lu-DOTATATE and found that a biological effective dose of less than 40 Gy was safe in patients without preexisting risk factors for renal disease, such as hypertension, diabetes, or old age (n = 16), and that a lower biological effective dose threshold of less than 28 Gy was safe for patients who did have these risk factors (n = 12) (42).
METHODS TO REDUCE NEPHROTOXICITY
Several different approaches to reduce the risk of nephrotoxicity in PRRT have been studied. Most of these methods aim at decreasing the radiation dose to the kidneys, such as by reducing the proximal tubular reabsorption of peptides. Other methods focus on reducing the harmful effects of the radiation. An overview of these methods is presented in Table 1.
Processes and Factors in Renal Handling and Retention of Radiolabeled Peptides and Strategies to Interfere
Reduction of Renal Uptake
Structural Modification of Radiolabeled Peptides
Various attempts have been made to change the elimination characteristics of radiolabeled peptides in order to reduce their renal retention. Structural change of a peptide may result in reduced glomerular filtration, such as by increasing size or protein binding, or in reduced tubular reabsorption, such as by reducing the affinity of the peptide for renal transporters. Obviously, affinity of the radiolabeled peptides for their specific receptors, which is important for accumulation in target tissues, should remain intact.
Several studies have shown that the introduction or substitution of positively or negatively charged groups can influence the renal uptake of peptides and antibody fragments. Ekblad et al. studied renal uptake of 99mTc-labeled HER2 Affibody molecules (ZHER2:342) with different chelators. They reported that the substitution of a negatively charged glutamic acid residue by a neutral serine residue in the chelator sequence mercaptoacetyl-Glu-Glu-Glu significantly reduced renal uptake without reducing tumor uptake (46). Miao et al. found that the renal uptake of 90Y- and 177Lu-labeled α-melanocyte–stimulating hormone analogs was decreased by adding a glutamic acid residue to the peptide. Although the uptake in melanoma was significantly decreased as well, an improved tumor-to-kidney uptake ratio was achieved (47). Removal of the anionic N-terminal pentaglutamic acid sequence of minigastrin analogs, or replacement by a histidine-histidine-glutamic acid sequence, resulted in a marked decrease in kidney uptake. Again this was accompanied by a decrease in tumor uptake but an increased tumor-to-kidney ratio (48). Replacing minigastrin's pentaglutamic acid sequence by a neutral pentaglutamine sequence reduced uptake in tumors and organs positive for the cholecystokinin 2 receptor more than it reduced renal uptake (3). 99mTc-labeled bombesin analogs were shown to express higher renal uptake when 2 glutamic acid residues were introduced, compared with 1 (49).
Akizawa et al. studied the renal uptake of 111In-diethylenetriaminepentaacetic acid (DTPA)-l-Phe1-octreotide analogs in which the neutral phenylalanine residue was substituted by a neutral methionine, a positively charged lysine, or a negatively charged aspartic acid residue. They observed that the renal uptake of 111In-DTPA-l-Lys1-octreotide was significantly higher than that of 111In-DTPA-l-Phe1-octreotide, whereas the renal uptake of 111In-DTPA-l-Asp1-octreotide was lower. However, the effect of these substitutions on the uptake in somatostatin receptor–positive organs was not reported (50). Dimeric cyclic RGD peptides also showed increased renal accumulation when a lysine residue was introduced in the spacer linking both RGD moieties (51).
Apart from the substitution, addition, or deletion of amino acids, other molecules can also be coupled to peptides to alter their pharmacokinetics. Examples include poly(ethylene glycol) (PEG) groups or albumin binding sequences. Addition of PEG groups (which can range in molecular weight from ∼2 to >200 kDa) is referred to as PEGylation (52,53). PEGylation can alter pharmacokinetics by increasing the size of the molecule, increasing hydrophilicity, masking charges, and decreasing antigenicity. Insertion of 2- to 4-kDa PEG groups was reported to reduce the renal uptake and increase the tumor-to-kidney ratio of 125I- and 18F-labeled RGD peptides (52,54). Addition of larger PEG groups or albumin binding sequences can reduce glomerular filtration and increase the circulatory half-life of peptides (53,55). Coupling of albumin binding sequences to radiolabeled Fab fragments and Affibody molecules resulted in an increased circulatory half-life, a reduced kidney uptake, and good tumor targeting, compared with the native constructs (55,56). However, an increased circulatory half-life also increases the bone marrow radiation dose.
Use of Different Radiolabels, Linkers, or Chelators
As described above, after endocytosis and catabolism in the proximal tubules, iodinated tyrosine can diffuse freely out of the lysosomes and the tubular cells, leading to relatively low renal retention of conventionally radioiodinated products. However, many peptides and antibodies are also internalized in the target organs or tumors, from which iodine is also rapidly cleared. This leads to low target-to-background ratios and renders conventionally 131I-labeled products less useful for labeling of internalizing antibodies or peptides (57).
Radiometal-chelated products are retained longer in targeted tissue but also in the kidneys. A strategy to reduce the kidney retention of radiolabeled peptides involves the insertion of metabolizable linkers between the radiometal–chelator complex and the peptide. These linkers must be stable in plasma. Specific linkers were designed to be cleaved in the kidneys by hydrolases at the proximal tubular brush border membrane, releasing the radiometal–chelator complex from the peptide before it undergoes endocytosis. The radiometal will then be excreted in the urine. Uehara et al. used a glycyl-lysine bond which can be cleaved by renal brush border enzymes. A radiolabeled construct of Fab fragments with these linkers, 188Re-tricarbonyl-(cyclopentadienyl)-glycyl-lysine-Fab, expressed significantly lower kidney uptake than the native construct. Tumor uptake was not affected (58).
Other linkers that have been studied are cleaved by lysosomal enzymes, releasing the radiometal–chelator complex from the cells after endocytosis. Tibben et al. found that a cleavable ester bond between a chelator and a Fab′ fragment reduced the kidney uptake of Fab′ fragments without affecting tumor uptake (59). Smith-Jones et al. studied the biodistribution of 111In-DTPA–labeled somatostatin analogs with several ester and diester linkers. None of the metabolizable linkers led to higher tumor-to-kidney ratios, because tumor uptake decreased more than kidney uptake (60).
Competitive Inhibition of Proximal Tubular Reabsorption
The interaction of peptides with receptors or transporters on the apical membrane of the proximal tubular cells may be disturbed by other ligands of these transporters. An excess of ligands in the ultrafiltrate during the elimination phase of the radiolabeled peptides causes competitive inhibition of the renal transport mechanisms, leading to a reduction in the uptake of the radiolabeled peptides. Again, the used substances should not interfere with the specific binding of the peptides to their native receptors, because this would also decrease uptake in target lesions.
The inhibition of tubular reabsorption of proteins by positively charged (basic) amino was first described in the 1970s. Several years later, coadministration of the basic amino acids lysine and arginine was found to reduce the renal uptake of radiolabeled antibody fragments and somatostatin analogs (61,62). Behr et al. reported that poly-l-lysine was more potent than l-lysine or d-lysine in reducing the renal retention of radiolabeled Fab′ fragments in rats, but poly-l-lysine was also more toxic. No difference between l-lysine and d-lysine was found (63). Rolleman et al. reported a significant decrease in parameters of kidney damage after coinfusion of lysine in rats receiving 177Lu-DOTATATE therapy (40). The same group studied the effects and side effects of coinfusion of an amino acid mixture, l-lysine, or a combination of l-lysine and l-arginine during PRRT with [111In-DTPA0]octreotide in patients. The combination of 25 g of lysine and 25 g of arginine was more effective than 50 g of lysine and caused relatively few side effects. The highest dose—75 g of lysine—was most effective in reducing renal uptake but frequently caused side effects, such as vomiting (64). Therefore, further increasing the lysine dose to improve kidney protection is not recommended. For somatostatin analogs, a maximum reduction of the kidney radiation dose of approximately 50% has been achieved (64,65). Coinfusion of basic amino acids is currently the standard renoprotective regimen in clinical PRRT.
The renal uptake of minigastrin, a peptide that lacks positively charged amino acid residues, is not reduced by coinfusion of lysine (4,66). Béhé et al. found that negatively charged polyglutamic acid chains of at least 5 glutamic acid residues did reduce renal uptake of minigastrin significantly (66).
Our group found that the plasma expander succinylated gelatin (Gelofusine; Braun) efficiently reduced the renal reabsorption of 111In-octreotide in humans (67) and of 111In-labeled octreotide, minigastrin, and exendin in rats (68,69). Succinylated gelatin consists of peptides with an average molecular weight of 30 kDa and a net negative charge. The group of Baum et al. used succinylated gelatin combined with the above-mentioned lysine/arginine mixture for kidney protection in 902 courses of PRRT with 90Y-DOTATOC (mean activity, 14 ± 4.8 GBq in 4 cycles) or 177Lu-DOTATATE (mean activity, 28 ± 6 GBq in 4 cycles). Since this regimen has been in use, no serious kidney toxicity has occurred. Mild to moderate allergic side effects (common toxicity criteria grade 1 or 2) of the renoprotective regimen were recorded in 1.4% of courses, and grade 3 reactions occurred in 0.6% (Richard P. Baum, written communication, November 2009). Extensive experience with the administration of succinylated gelatin to patients in acute care or during surgical procedures exists, and serious side effects are rare. Severe anaphylactoid reactions are described in approximately 0.04% of patients (70).
We recently described that coadministration of fractionated albumin or an albumin-derived peptide efficiently reduced renal uptake of radiolabeled peptides in rats. Five milligrams of the albumin-derived peptide was equally effective as 20 mg succinylated gelatin in reducing renal uptake of 111In-octreotide, 111In-minigastrin, and 111In-exendin (71). This peptide consists of the 36 C-terminal amino acid residues of albumin and contains 3 positively and 6 negatively charged residues.
The wide variety of substances able to reduce renal reabsorption of radiolabeled peptides may well reflect the nonselective nature of renal transport systems such as the megalin receptor. The selectivity of certain competitive inhibitors, such as lysine for octreotide and polyglutamic acid for minigastrin, may reflect their binding to different sites on megalin, or the involvement of other receptors or transporters.
Alternative Ways to Interfere with Proximal Tubular Reabsorption
Apart from competitive inhibition, the process of endocytosis may be disturbed at various other stages. Albumin endocytosis was reported to be decreased by extracellular hypertonicity, which interferes with the formation of clathrin lattice, thus inhibiting the formation of clathrin-coated pits (72). Gekle et al. also reported that alkalinization of endosomes by the addition of NH4Cl or bafilomycin A1 to opossum kidney cells in vitro reduced the receptor-mediated endocytosis of albumin but did not affect fluid-phase endocytosis of dextran (73). The same group described reduced endocytosis of albumin by cytochalasin D, which disrupts the actin cytoskeleton, and by nocodazole, which interferes with microtubule formation (72). However, toxicity of these agents may preclude their clinical use. Disruption of microtubule formation by pretreatment of rats with colchicine also significantly reduced renal uptake of 111In-octreotide (74).
Because the process of endocytosis requires energy, disrupting the energy supply also reduces endocytosis. De Jong et al. showed that maleate, an inhibitor of the citric acid cycle, significantly reduced renal uptake of 111In-octreotide in rats. However, because of the reported renal toxicity of maleate, its use in PRRT was not studied further. Acidification of urine by administration of NH4Cl significantly reduced renal uptake of 111In-octreotide as well, possibly through neutralization of negatively charged membrane binding sites (65).
Stahl et al. reported that probenecid, an inhibitor of OATs and multidrug resistance proteins, reduced renal uptake of 111In-octreotide in rats by 30%. Octreotide is an organic cation, but OATs have been shown to transport neutral and weak cations as well as anions (25).
The influence of increasing urinary flow on the excretion of radiolabeled peptides has been studied in rats as well, either by increasing fluid intake or by administering furosemide. However, increasing fluid intake had no effect on renal uptake of 111In-octreotide (65), and furosemide even caused a 44% increase in uptake (25).
Reduction of Nephrotoxicity at a Given Radiation Dose
Other approaches to preventing nephrotoxicity aim at reducing the damage induced by a given absorbed radiation dose.
Use of Radioprotectors
Radioprotectors reduce oxidative stress by scavenging free radicals. Because the formation of free radicals depends on the presence of oxygen, healthy, well-oxygenated tissues are protected to a larger extent than tumors, which are usually hypoxic. Furthermore, amifostine, the prototype radioprotector, is converted to its active metabolite by alkaline phosphatase. In healthy tissue, including the renal brush border membrane, this enzyme is expressed at much higher levels than in tumor capillaries, leading to 10- to 100-fold higher concentrations of the active metabolite in healthy tissue than in tumor cells (75). Amifostine is approved for use in EBRT and has been studied in PRRT. Rolleman et al. coadministered this radioprotector to rats during administration of 177Lu-DOTATATE and for 7 d after PRRT. They found that amifostine was equally as effective as lysine in preventing nephrotoxicity (76).
Use of Mitigating Agents
Mitigating agents aim at reducing the risk of nephrotoxicity by inhibiting counterproductive tissue reactions to radiation-induced damage (77). From research in EBRT it is known that inhibition of the renin-angiotensin-aldosterone system after irradiation of the kidneys can prevent nephropathy (78). Activation of the renin-angiotensin-aldosterone system can be inhibited by angiotensin-converting-enzyme inhibitors, angiotensin II receptor blockers, or aldosterone receptor blockers. Jaggi et al. studied the effects of renin-angiotensin-aldosterone system inhibition in mice that were injected with α-emitting 225Ac-labeled untargeted antibodies. Starting 10 d after injection, the mice were treated for 39 wk with spironolactone (an aldosterone receptor antagonist), captopril (an angiotensin-converting-enzyme inhibitor), L-158,809 (an angiotensin II receptor 1 blocker), or placebo. Animals receiving placebo developed significant kidney damage (tubulolysis, thickening of basement membranes, and elevated blood urea nitrogen), which was efficiently reduced by administration of spironolactone and to a lesser extent by L-158,809, but not by captopril (77).
Dose Fractionation
Dose fractionation is used in EBRT to use the potential of normal tissue to repair sublethal damage between irradiation cycles. In radionuclide therapy, dose fractionation is difficult because of the relatively low dose rates, long residence times, and long half-lives of the radionuclides. In PRRT, comparisons between different dose fractionation schemes are scarce. Barone et al. analyzed 18 patients who were treated with 90Y-DOTATOC and found that patients receiving large radiation doses with little fractionation had the highest risk of developing nephrotoxicity (45). Rolleman et al. studied fractionation of 177Lu-DOTATATE therapy in rats. Dividing the total dose over 2 fractions significantly reduced kidney damage, and 3 fractions caused significantly less kidney damage when given at weekly or monthly intervals than when given at daily intervals. No significant differences in tumor response were found between these regimens (40).
Influence of Radionuclide Characteristics
Renal toxicity appears to occur less frequently after therapy with 177Lu-DOTATATE than after therapy with 90Y-DOTATOC (42). Of 504 patients receiving up to 30 GBq of 177Lu-DOTATATE (predicted kidney dose, <23 Gy), only 2 (0.4%) developed nephrotoxicity, which was probably unrelated to PRRT (31). In another series of 60 patients receiving up to 15 GBq of 90Y-DOTATOC (again, predicted kidney dose was <23 Gy), 7 (12%) developed kidney toxicity (5). The difference may be due to the lower β-energy and the consequently lower range of the β-emission of 177Lu, which results in a relatively lower radiation dose to the radiosensitive glomeruli.
Combination Strategies
Several groups have studied combinations of renoprotective strategies for PRRT in rats. Combining lysine and succinylated gelatin showed additive reduction of the kidney uptake of 111In-DTPA-octreotide and 111In-DOTA-octreotate, whereas tumor uptake was not affected (4,79). Kidney uptake of 111In-exendin was reduced most potently by a combination of polyglutamic acid and succinylated gelatin (4). Combining lysine with colchicine also additively reduced the renal uptake of 111In-DTPA-octreotide (74). All above-mentioned strategies used combinations of agents that interfere with proximal tubular endocytosis. Combinations of these with other strategies, such as radiation protection or mitigation, are likely to further reduce the risk of nephropathy.
CONCLUSION AND FUTURE DIRECTIONS
High kidney uptake of radiolabeled peptides can cause dose-limiting nephrotoxicity after PRRT. Reduction of this uptake can enable the administration of higher activities, which improves the effectiveness of therapy. Although administration of higher activities also increases the risk of bone marrow toxicity, the high radiation dose to the kidneys remains a problem in current somatostatin analog PRRT with renoprotective regimens, especially when 90Y-labeled analogs are used. For other peptides such as minigastrin and exendin, which express significantly higher kidney uptake than somatostatin analogs, efficient reduction of kidney uptake will be even more important to enable safe PRRT. Evidence indicates that the most important pathway for the renal uptake of most radiolabeled peptides (including antibody fragments) consists of glomerular filtration followed by receptor-mediated endocytosis and retention in the proximal tubular cells. Many strategies have been designed to improve the target-to-kidney ratio of radiolabeled peptides, several of which show promising results. Subtle changes in the amino acid sequence of a peptide, thereby changing its structure or charge distribution, can affect renal uptake considerably. Several peptide analogs have been designed that exhibit lower renal retention with unaltered target uptake. Another approach focuses on enhancing the clearance of radiolabeled products from the kidneys. Linkers that are selectively cleaved in the kidneys may prevent the accumulation of radiolabels in the kidneys without loss of retention in targeted tissues. In clinical PRRT, an established method to reduce the absorbed kidney dose is the coinfusion of competitive inhibitors of proximal tubular reabsorption. These have been shown to significantly reduce the renal uptake of various peptides. However, a baseline renal uptake of radiolabeled somatostatin analogs seems to persist, possibly reflecting nonsaturatable fluid phase endocytosis. Different methods to interfere with endocytosis, such as disruption of microtubule formation, may also reduce fluid-phase endocytosis. To prevent nephrotoxicity at a given radiation dose, dose fractionation and treatment with radioprotectors or mitigating agents have shown promising results.
Because most of these strategies interfere with different mechanisms of kidney uptake and damage, the combination of 2 or more strategies is likely to reduce kidney damage progressively. Combining 2 agents, both of which reduce endocytosis, has already been shown to further reduce the kidney uptake of various radiolabeled peptides.
Most strategies for the reduction of kidney damage in PRRT and RIT have, as yet, not been tested on patients, and more research is needed to study their potential for clinical application. If we can further reduce the risk of renal damage caused by radiolabeled peptides, higher-activity doses can be administered safely, and highly effective targeted tumor therapy comes within reach.
Acknowledgments
We thank Professor R.P. Baum (Bad Berka, Germany) and his group for sharing their preliminary data on the combined renoprotective regimen of succinylated gelatin and lysine/arginine in clinical PRRT.
Footnotes
-
COPYRIGHT © 2010 by the Society of Nuclear Medicine, Inc.
-
Learning Objectives: On successful completion of this activity, participants should be able to describe (1) mechanisms of renal accumulation of radiolabeled peptides, proteins, and antibody fragments; (2) different techniques to reduce kidney damage in peptide-receptor radionuclide therapy and antibody fragment radioimmunotherapy; and (3) factors determining the risk of nephrotoxicity in radionuclide therapies.
-
Financial Disclosure: The authors of this article have indicated no relevant relationships that could be perceived as a real or apparent conflict of interest.
-
CME Credit: SNM is accredited by the Accreditation Council for Continuing Medical Education (ACCME) to sponsor continuing education for physicians. SNM designates each JNM continuing education article for a maximum of 1.0 AMA PRA Category 1 Credit. Physicians should claim only credit commensurate with the extent of their participation in the activity.
-
For CE credit, participants can access this activity through the SNM Web site (http://www.snm.org/ce_online) through July 2011.
References
- 1.↵
- 2.↵
- 3.↵
- 4.↵
- 5.↵
- 6.↵
- 7.↵
- 8.↵
- 9.↵
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.↵
- 28.↵
- 29.↵
- 30.
- 31.↵
- 32.↵
- 33.↵
- 34.↵
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.↵
- 57.↵
- 58.↵
- 59.↵
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.↵
- 79.↵
- Received for publication January 16, 2010.
- Accepted for publication March 24, 2010.





{kind=link}
Jump to section
Related Articles
Cited By...
- Preclinical Characterization of DPI-4452: A 68Ga/177Lu Theranostic Ligand for Carbonic Anhydrase IX
- Pre-clinical Evaluation of Biomarkers for Early Detection of Nephrotoxicity Following Alpha-particle Radioligand Therapy
- Choosing the Right Metrics for Evaluation of Radiopharmaceutical Therapy Dosimetry Methodologies
- Phase I Study of [68Ga]Ga-Anti-CD206-sdAb for PET/CT Assessment of Protumorigenic Macrophage Presence in Solid Tumors (MMR Phase I)
- 225Ac-MACROPATATE: A Novel {alpha}-Particle Peptide Receptor Radionuclide Therapy for Neuroendocrine Tumors
- Optimizing Immuno-PET Imaging of Tumor PD-L1 Expression: Pharmacokinetic, Biodistribution, and Dosimetric Comparisons of 89Zr-Labeled Anti-PD-L1 Antibody Formats
- Toward a Patient-Specific Traceable Quantification of SPECT/CT-Based Radiopharmaceutical Distributions
- Normal-Tissue Tolerance to Radiopharmaceutical Therapies, the Knowns and the Unknowns
- Imaging Reveals Importance of Shape and Flexibility for Glomerular Filtration of Biologics
- Effect of Modulating FcRn Binding on Direct and Pretargeted Tumor Uptake of Full-length Antibodies
- Recombinant {alpha}1-Microglobulin Is a Potential Kidney Protector in 177Lu-Octreotate Treatment of Neuroendocrine Tumors
- Preclinical Investigation of 212Pb-DOTAMTATE for Peptide Receptor Radionuclide Therapy in a Neuroendocrine Tumor Model
- 90Y-DOTATOC Dosimetry-Based Personalized Peptide Receptor Radionuclide Therapy
- Theranostic Radiolabeled Anti-CD20 sdAb for Targeted Radionuclide Therapy of Non-Hodgkin Lymphoma
- Predicting the response to CTLA-4 blockade by longitudinal noninvasive monitoring of CD8 T cells
- Approaches to Improve the Pharmacokinetics of Radiolabeled Glucagon-Like Peptide-1 Receptor Ligands Using Antagonistic Tracers
- 89Zr-Labeled Versus 124I-Labeled {alpha}HER2 Fab with Optimized Plasma Half-Life for High-Contrast Tumor Imaging In Vivo
- Comparative Studies of Three 68Ga-Labeled [Des-Arg10]Kallidin Derivatives for Imaging Bradykinin B1 Receptor Expression with PET
- The Somatostatin Analog 188Re-P2045 Inhibits the Growth of AR42J Pancreatic Tumor Xenografts
- Improved Tumor Targeting of Anti-HER2 Nanobody Through N-Succinimidyl 4-Guanidinomethyl-3-Iodobenzoate Radiolabeling
- Synthesis and In Vitro and In Vivo Evaluation of Hypoxia-Enhanced 111In-Bombesin Conjugates for Prostate Cancer Imaging
- Synthesis, Preclinical Validation, Dosimetry, and Toxicity of 68Ga-NOTA-Anti-HER2 Nanobodies for iPET Imaging of HER2 Receptor Expression in Cancer
- DOTA Conjugate with an Albumin-Binding Entity Enables the First Folic Acid-Targeted 177Lu-Radionuclide Tumor Therapy in Mice
- Cohort Study of Somatostatin-Based Radiopeptide Therapy With [90Y-DOTA]-TOC Versus [90Y-DOTA]-TOC Plus [177Lu-DOTA]-TOC in Neuroendocrine Cancers
- Different Radioactivity Uptake between Somatostatin Analogues Labelled with 111In and 90/88Y in Rat Kidney
- Receptor Affinity and Preclinical Biodistribution of Radiolabeled Somatostatin Analogs
- Therapeutic strategies for advanced neuroendocrine carcinomas of jejunum/ileum and pancreatic origin