


Abstract
The thyroid transcription factors TTF-1 and Pax8 cooperate in the transcriptional activation of thyroid-specific genes such as thyroglobulin (Tg), thyroperoxidase (TPO), and sodium/iodide symporter (NIS). Methods: Dog TTF-1 (dTTF-1) and the human Pax8 (hPax8) gene were transfected in Morris hepatoma (MH3924A) cells to investigate (a) the possible visualization of functional protein–protein interaction and (b) the induction of thyroid-specific gene expression. In MH3924A cell lines expressing dTTF-1, hPax8, or both, the activation of human Tg (hTg), human TPO (hTPO), or rat NIS (rNIS) promoter/enhancer was measured using firefly luciferase reporter constructs. Furthermore, the possible induction of thyroid-specific genes was investigated in iodide uptake and reverse transcription polymerase chain reaction (RT-PCR) experiments. Results: Low transcriptional activation of these constructs was observed in cells expressing either hPax8 or dTTF-1 alone. In contrast, the hTg and hTPO and, to a lesser extent, the rNIS regulatory region were significantly activated in cell lines expressing both transcription factors. Imaging the transcriptional activation of the thyroid-specific regulatory regions by Pax8 and TTF-1 was possible in nude mice implanted with MHhPax8dTTF-1 cells using a cooled charge-coupled device camera. Na125I uptake experiments and RT-PCR showed no effect of hPax8 and dTTF-1 on endogenous thyroid-specific gene expression in genetically modified cells. Conclusion: The activation of thyroid-specific promoter/enhancer elements in Morris hepatoma cells depends on the functional interaction of hPax8 and dTTF-1. The cooperation of these 2 transcription factors can be visualized in vitro as well as in vivo. With regard to a possible application for radioiodine therapy, further modifications are required.
- thyroid transcription
- thyroid transcription factors human Pax8 and dog TTF-1
- protein–protein interaction
- bioluminescence imaging
Tissue-specific transcriptional regulation is often mediated by a set of transcription factors whose combination is unique to specific cell types. The vast majority of genes expressed in a cell type–specific manner is regulated by promoters containing a variety of recognition sequences for tissue-specific and ubiquitous transcription factors. It is the precise functional interaction between these various regulating proteins and the regulatory DNA sequences that enables individual cell types to play their role within an organism.
Thyroid follicular cells, the most abundant cell population of the thyroid gland, are responsible for thyroid hormone synthesis and are characterized by the expression of thyroglobulin (Tg), thyroperoxidase (TPO), the thyrotropin-stimulating hormone receptor (TSHr), and the sodium/iodide symporter (NIS) (1,2). To date, 3 transcription factors that specifically regulate thyroid-specific gene expression have been identified: Thyroid transcription factor-1 (TTF-1) is a homeodomain-containing protein expressed in embryonic diencephalon, thyroid, and lung (3). Thyroid transcription factor-2 (TTF-2), a forkhead domain–containing protein, is expressed in the pituitary gland and thyroid; and Pax8, which belongs to the Pax family of paired domain–containing genes, is expressed in the kidney, the developing excretory system, and the thyroid (4,5). None of these transcription factors is expressed exclusively in the thyroid, but their combination is unique to this organ and is likely to be responsible for differentiation of thyrocytes. In recent years, a variety of studies focused on the molecular mechanisms of the TTF-1 and Pax8 activity and the functional role of TTF-1–and Pax8-binding sites within Tg and TPO promoters. Three TTF-1–binding sites are present in the Tg and the TPO promoter. Using reporter constructs with thyroid-specific promoters, TTF-1 was shown to activate transcription from the Tg promoter and, albeit at a much lower extent, from the TPO promoter in both thyroid and nonthyroid cells (6). Pax8 binds to a single site on the Tg and on the TPO promoters; in both cases, the Pax8-binding site overlaps with that of TTF-1 (7). In addition, Pax8 is involved in the regulation of the rat gene coding for the NIS (8). In transient transfection assays, Pax8 was shown to activate transcription from exogenous Tg and TPO promoters in nonthyroid cells (7). Both transcription factors directly interact and synergistically activate thyroid-specific transcription (6,9) and, therefore, represent a promising model system for the visualization of protein–protein interaction. Recently, new technologies based on reporter gene expression have been developed to monitor physical protein–protein interaction using complementation and reconstitution strategies (10–12). It was shown that luciferase expression can be imaged in living animals by using a cooled charge-coupled device (CCD) camera that is relatively sensitive and quantifiable and allows for noninvasive and repeated studies in the same animal (12).
In the present study, we were interested in the visualization of functional protein–protein interaction by the ability of dTTF-1 and hPax8 to induce reporter gene expression in rat Morris hepatoma (MH) cells. In addition, the ability of these transcription factors to induce endogenous thyroid-specific gene expression and iodide accumulation in nonthyroid tumor cells was investigated.
Here, we demonstrate the synergistic activation of human Tg (hTg), human TPO (hTPO), and rat NIS (rNIS) promoters in dog TTF-1 (dTTF-1)–and human Pax8 (hPax8)-expressing tumor cells in vitro and in vivo. However, even in presence of the histone deacetylase inhibitor (HDAC) trichostatin A, we failed to detect Tg, TPO, or NIS gene expression driven by endogenous promoters in the dTTF-1–and hPax8-expressing cell lines.
MATERIALS AND METHODS
Construction of Plasmids and Expression Vectors
To generate the reporter constructs phTPOluc and prNISluc, the enhancer and promoter region of the rNIS gene and hTPO gene were amplified by polymerase chain reaction (PCR) using the specific primers for the rNIS enhancer (reverse GTCTAGAAGAAGGTGTTTGGCC; forward ATCATGAAGGTGGAAGAGATTG), for the rNIS promoter (forward GCTTACGAGCCTGCCCTAGTC; reverse ATGGAGACAGGTGACTCGGTG), for the hTPO enhancer (forward (GGGATTCCATCTTTTATTCTAATGC; reverse CTCGAGTGGCCATCCATGATTTGGC), and for the hTPO promoter (reverse AATTGCCTTCCAGGGACTTG; forward CTCTTTTGATGGGTGCATGATG).
PCR was performed in 35 cycles under the following conditions: 2.5 U Taq DNA polymerase (Böhringer Mannheim), 200 μmol/L deoxynucleoside triphosphate, 1 μmol/L primer concentration, 1.5 mmol/L MgCl2, and 10% dimethyl sulfoxide (DMSO) in a total reaction volume of 50 μL. The denaturation was done at 95°C for 1 min; the annealing was at 59°C (rNIS enhancer), 62.5°C (rNIS promoter), and 62°C (hTPO enhancer and promoter) for 1 min; and the primer extension was at 74°C for 10 min.
The PCR products were cloned into the pGEM-T-Easyh (Promega) and sequenced, and the promoter sequences were inserted into the bluescript vector (pSK) with EcoRI, SpeI (hTPOprom) or SpeI, NotI (rNISprom), leading to the plasmids pSKhTPOprom and pSKrNISprom. The rNIS enhancer was cloned with SacII blunt, SpeI into the pSKrNISprom digested with EcoRV, SpeI, resulting in pSKrNISenh/prom. For construction of the luciferase expression vector (pluc), the pAP1-Luc vector containing the firefly luciferase gene from Photinus pyralis (Clontech) was used. The phTPOluc vector was constructed by first cloning the hTPOprom XbaI blunt, KpnI into the pAP1-Luc expression vector followed by inserting the hTPO enhancer upstream of the TPO promoter after EcoRI digestion. The prNISluc expression vector was obtained by inserting the rNIS enhancer/promoter fragment digested from the pSKrNISenh/prom into the pAP1-Luc.
To generate the reporter plasmid phTgenh(2)promluc, the 500-bp BstXI, SphI fragment excised from the entire hTg enhancer sequence (obtained from Daniel Christophe) (13) was blunted with Klenow enzyme and cloned in 2 copies into the HincII site and into the blunted EcoRI site, respectivley, of pSK (pSKhTgenh(2)prom). This was done to enhance the Tg promoter activity. The hTg enhancer fragments were excised from pSK with KpnI and BamHI and inserted upsteam of the hTg promoter (obtained from Daniel Christophe, Universite Libre de Bruxelles, Bruxelles, Belgique) (14) into the pSKhTg plasmid digested with KpnI, BglII, thereby replacing the complete Tg enhancer. The hTgenh(2)prom fragment excised with XbaI and KpnI was ligated to the pAP1-Luc vector digested with HindIII and KpnI after the XbaI terminus and the HindIII terminus had been blunted with the Klenow enzyme. The pTRUFCMVluc bearing the firefly luciferase gene controlled by the cytomegalovirus (CMV) promoter was cloned as described elsewhere (15).
To transfer the dTTF-1 gene (95% homology with human TTF-1) or the hPax8 gene and the neomycin (neo) or the hygromycin (hyg) antibiotic resisitance gene controlled by the elongation factor 1α (EF1α) promoter, the bicistronic retroviral vector M48 based on the Moloney murine leukemia virus vector (obtained from Olivier Danos, Généthon, Evry, France) (16) was constructed. To ensure simultaneous expression of both the transcription factor gene and the antibiotic resistance gene, and stabilization of the mRNA, a synthetic intron (IVS) and an internal ribosomal entry site (IRES) from encephalomyocarditis virus were inserted between the coding sequences (Fig. 1) (17,18). Briefly, the hPax8 open reading frame (obtained from Peter Gruss, Max-Planck Institute of Biophysical Chemistry, Göttingen, Germany) (19) was cloned with SmaI into a pSKEF1αIVSIRES1hyg plasmid and the EF1αhPax8IVSIRES1hyg fragment was inserted with SalI blunt NsiI into the M48 vector. To construct the M48EF1αdTTF-1IVSIRES1neo vector, the dTTF1 gene (provided by Christiane Christophe-Hobertus, Institut de Recherche Interdisciplinaire en Biologie Humaine et Nucleare, Bruxelles, Belgique) (20) was cloned MseI blunt into the SmaI site of pSK and inserted afterward into a pSKEF1αhPax8IVSIRES1neo by EcoRI and BamHI, thereby replacing the hPax8 gene. Finally, the EF1αdTTF1neo1 fragment was cloned by XbaI blunt, XhoI into the M48 vector prepared by BamHI blunt, XhoI.
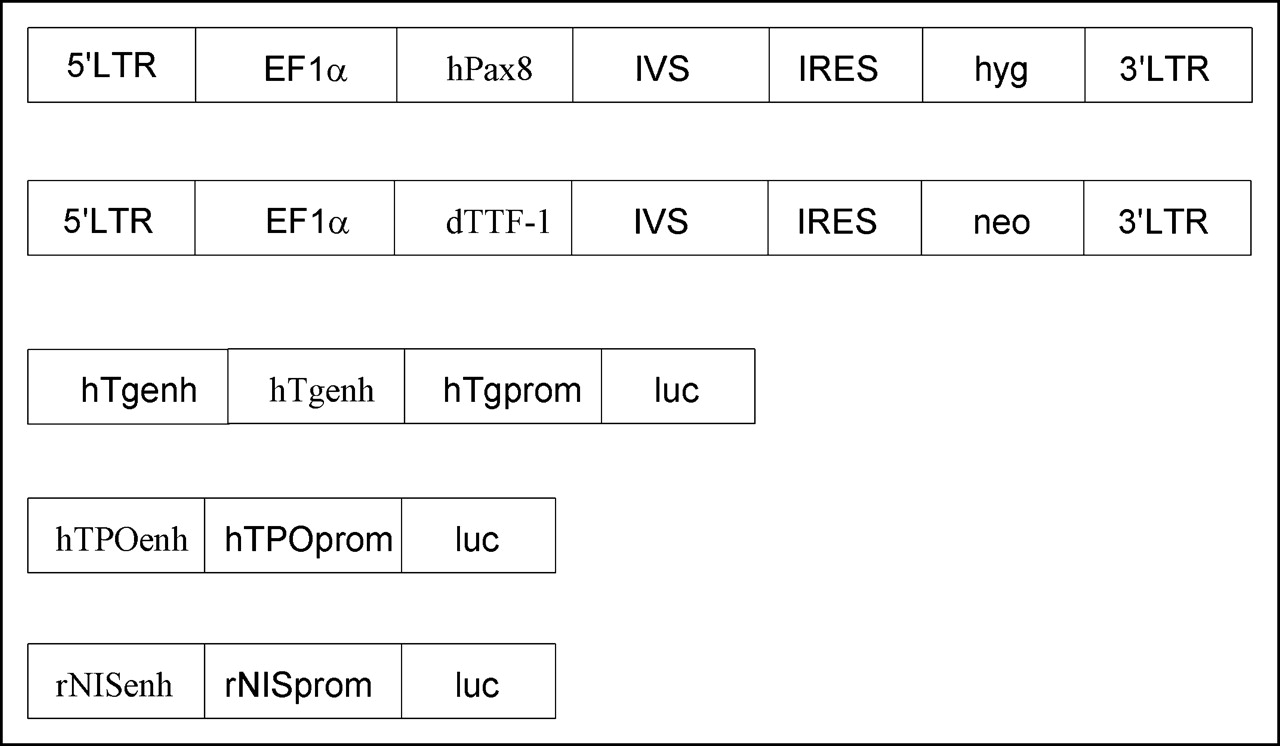
Structure of recombinant retroviral vector and reporter plasmids. M48EF1αhPax8IVSIREShyg and M48EF1αdTTF-1IVSIRESneo vectors are designed to transfer human Pax8 and hygromycin (hyg) resistance activity or dTTF-1 and neomycin (neo) resistance activity by use of internal ribosomal entry site (IRES) from encephalomyocarditis virus. A synthetic intron (IVS) is inserted to stabilize messenger RNA. Expression of genes is regulated by EF1α promoter. Reporter gene constructs prNISLuc, phTPOLuc, phTg(2)Luc, and pTRUFCMVluc were used for transient transfection of MH3924A and MHhPax8dTTF-1/14 cells. LTR = long terminal repeat.
Tranfection Procedures and Culture
The MH3924A cell line and the transient packaging cell line BOSC23 (21) used for the production of ecotropic retroviral particles were cultured in RPMI 1640 medium (Gibco BRL) supplemented with 10% (BOSC23) or 20% (MH3924A) fetal calf serum (FCS). The rat follicular thyroid cell line FRTL5, used as a positive control in the reverse transcription (RT)-PCR, was cultured in Ham’s F12 medium supplemented with somatostatin (10 ng/mL), Gly-His-Gly (10 ng/mL), transferrin (5 μg/mL), hydrocortisone (10 nmol/L), insulin (10 μg/mL), TSH (10 mU/mL), and 5% FCS. The cell lines were cultured at 37°C, in an atmosphere of 95% air and 5% CO2.
For transient packaging of the recombinant retroviral DNA, BOSC23 cells (60% confluent) were transfected with 2 μg DNA in 60-mm Petri dishes using the Lipofectamine Plus Reagent (Invitrogen, Life Technologies) according to the manufacturer’s instructions. After 2 d, the medium was centrifuged to remove detached BOSC23 cells and used for the infection of wild-type or recombinant MH3924A cells in the presence of Polybrene (Sigma; 8 μg/mL) overnight. After 2 d, the cells were cultured in hygromycin (0.5 mg/mL)–or G418 (0.8 mg/mL)–containing medium for at least 4 wk until resistent cell lines were established.
For assessment of the luciferase expression in various cell lines, 3 × 104 cells per well were plated in 24-well plates and transfected with 0.4 μg per well pTRUFCMVluc, phTg(2)luc, phTPOluc, or prNISluc 48 h later using the Lipofectamine Plus Reagent. Mock-transfected cells were used as the control. Twenty-four hours after transfection, the luciferase activity in the cells was determined using a luminometer as described.
For in vivo studies, MH3924A or MHhPax8dTTF-1/14 cells were plated in 100-mm Petri dishes, incubated until 80% confluence was reached, and transfected with 4 μg of different luciferase expression vectors. Twenty-four hours later, the cells were trypsinized and washed twice with Optimem (Gibco BRL); aliquots of 6 × 106 cells in 100 μL Optimem were prepared for imaging experiments as described.
Luminometer Measurements
To evaluate the luciferase activity in transiently transfected cells, a Luciferase Assay System (E1500; Promega) was used. Briefly, the culture medium was removed carefully from the 24-well-plates and the cells were washed with phosphate-buffered saline (PBS) before 200 μL lysis buffer per well were added. After centrifugation, 20 μL of the cell lysate were transferred to each well of a luminoscan plate and Luciferase Assay Reagent (Promega; 100 μL/well) was added immediately before the well was read over 10 s in a luminometer. The protein content of the cell lysates was determined with the Bio-Rad protein assay system in an Amersham spectrophotometer at 595 nm, and the luminescence results were reported as relative light units/μg protein. To account for differences in transfection efficiency, the luminescence was normalized to the signal obtained after transfection with the pCMVluc construct.
Northern Blot Analysis and RT-PCR
Five micrograms of RNA prepared by the trizol/chloroform procedure (Invitrogen) were electrophoresed on a 1% 3-morpholino-propanesulfonic acid (MOPS)-buffered agarose gel running in 2× loading dye solution (MBI Fermentas). RNA was blotted onto positively charged nylon membranes (Genescreen; PerkinElmer, Life Science) with 20× standard saline citrate, and hybridization and washing were performed according to Church and Gilbert (22). The blot was probed with complementary DNA (cDNA) fragments derived by restriction digestion of the dTTF-1 and hPax8 coding sequence and labeled using the HexaLabel DNA labeling kit (no. K0612; MBI Fermentas) and priming with [α-32P]deoxycytidine triphosphate. A 1760-bp fragment derived from human β-actin was used as the control.
For RT-PCR, 5 μg prepared from tichostatin A–treated (10 ng/mL) and nontreated MH3924A and MHhPax8/dTTF-1/14 cells and from the rat follicular thyroid cell line FRTL5 were digested with deoxyribonuclease I (DNase I) (RQ1 ribonuclease-free DNase [RNase-free DNase]; Promega) and used to synthesize cDNA using the Superscript reverse transcriptase kit with random hexamers as primers. The cDNA was subsequently used as the template for the PCR using specific primers for the rTg (forward AGCAGCTACATTAACAGCAC; reverse ACACCATTTCCATCACATCTC), the rTPO (forward TCTTCTCTACTGCTGCCTTC; reverse TCTCCCACCAAAACCTGTC), or the rNIS (forward ATTTTTGAGGATCTGCCCG; reverse GGGCAGGGCATGTATCAG) gene. One fifth of the total volume of the PCR was analyzed on a 1% agarose gel. As the control, recombinant β-actin primers were used for the RT-PCR, leading to a 660-bp fragment.
Imaging Luciferase Expression In Vivo Using a CCD Camera
Bioluminescence measurements of nude mice were performed under gas anesthesia in a proprietary dark chamber. Images were acquired in a constant experimental setup using a Hamamatsu OrcaII-ER cooled CCD camera (Hamamatsu Photonics GmbH) in combination with a wide-angle, high-aperture lens (CNG1.4/8; Jos. Schneider GmbH). The working distance was in the range of 5 cm. For the detection of luciferase expression, female mice (Swiss cCD1 nu/nu; Charles River Laboratories) were injected subcutaneously into the left (mock-transfected cells) or right (transfected cells) flank with an aliquot of 6 × 106 transiently transfected or mock-transfected MH3924A or MHhPax8/dTTF-1/14 cells in 100 μL Optimem. Immediately after cell implantation, 3 mg 1-(4,5-dimethoxy-2-nitrophenyl)ethyl ester (DNPE)-caged Luciferin (Molecular Probes) in 210 μL DMSO were injected intraperitoneally. After 40 min, the mice were imaged using a 5-min exposure time and a binning of 8 × 8 pixels for optimal sensitivity. A pause of 40 min between injection and imaging had been determined to yield maximum luminescence activity in vivo in prior experiments. Measured photon counts were superimposed in color on white light images of the respective animal. All units are photon counts after subtraction of read noise.
Measurement and Modulation of 125I Uptake
The iodide uptake was determined in triplicate vials as described previously (17,23) and the experiments were repeated twice. The recombinant MHhPax8, MHdTT-1, and MHhPax8dTTF-1 cell lines and the MH3924A wild-type cells were cultured for 48 h in 6-well plates before being incubated with 74 kBq Na125I (Amersham Buchler Germany; specific activity, 625.3 MBq/μg; radioactive concentration, 3.7 GBq/mL; radiochemical purity, 99.3%) for 4 h. For the modulation of the iodide uptake, the cell lines were incubated in Na125I medium supplemented with or without 50 μmol/L sodium perchlorate (Sigma). Furthermore, the HDAC inhibitor trichostatin A (10 ng/mL) was added to the cell cultures 24 h before the uptake began. After washing twice with ice-cold PBS, the cells were lysed with 0.3 mol/L sodium hydroxide on ice. Using an automated NaI(Tl) well counter (Cobra II; Canberra Packard), the radioactivity was measured in cell lysates and in the medium. The viable cell number was determined in a Coulter counter (Coulter Electronics) and by trypan blue staining (>94% viable cells). Using a Coulter counter, a mean cell volume of 5.66 ± 0.01 fL was measured, with no difference between wild-type and genetically modified cells.
Data Analysis and Statistics
Results are presented as mean + SD. Statistical comparisons between groups were performed by the Mann–Whitney U test–Wilcoxon rank sum W test or by the unpaired Student t test using the SIGMASTAT program (Jandel Scientific).
RESULTS
Generation and Characterization of Stable hPax8- and dTTF-1–Expressing MH Cell Lines
MH3924A cells were infected with recombinant retroviruses bearing the hPax8 and the hygromycin resistance gene or the dTTF-1 and the neomycin resistance gene. After selection with the respective antibiotic drug, 20 resistant cell lines were established for each single or both constructs. Northern blot analysis was done in 5 of 20 hPax8- or dTTF-1–transfected cell lines and in 7 of 20 hPax8/dTTF-1–transfected MH cell lines. Transcription of the hPax8 gene was found in 4 of 5 MHhPax8 cell lines (lines 3, 5, 13, and 20) and transcription of the dTTF1 gene was found in 3 of 5 MHdTTF-1 cell lines (lines 7, 11 and 21; data not shown). To establish MH cell lines expressing both hPax8 and dTTF-1, the MHhPax8/20 cells were transfected with the dTTF-1 gene and selected with neomycin. Northern blot analysis revealed transcription of both the hPax8 and the dTTF-1 gene in 5 of 7 MHhPax8/dTTF-1 cell lines (lines 2, 4, 5, 8, and 14; data not shown).
Thyroid-Specific Promoter Activity in hPax8- and dTTF-1–Expressing Hepatoma Cells
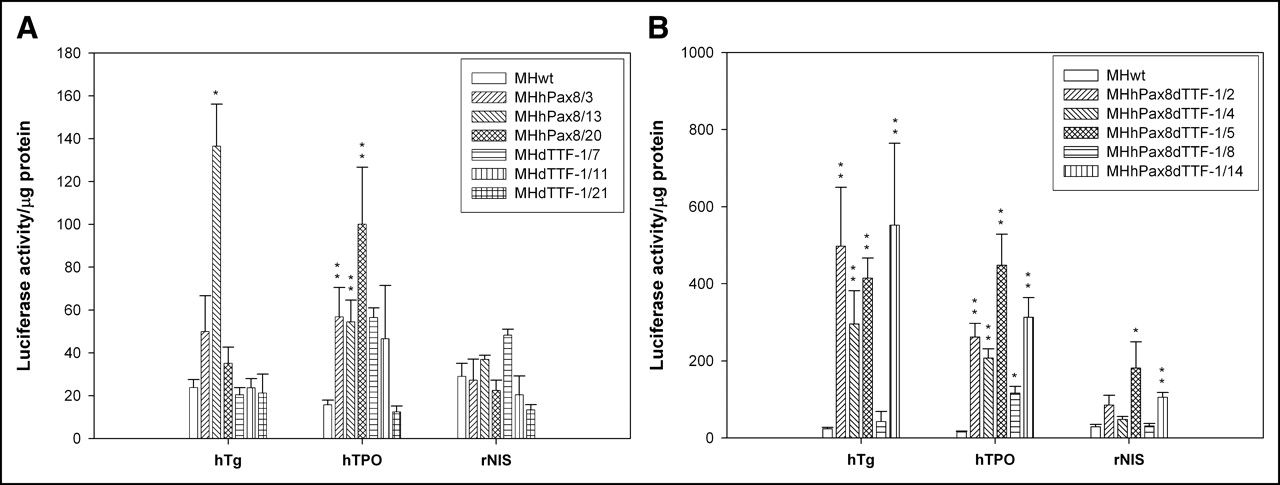
Individual MH cell lines expressing hPax8 or dTTF-1 or both transcription factors were compared with regard to their ability to transcriptionally activate the enhancer and promoter sequences of Tg, TPO, and NIS. To this end, the cells were transiently transfected with plasmids encoding the luciferase gene controlled by the regulatory sequence of one of the thyroid genes or by the CMV promoter. In lysates of the MHhPax8/3, MHhPax8/13, and MHhPax8/20 cell lines or MHdTTF-1/7, MHdTTF-1/11, and MHdTTF-1/21 cell lines, the expression of the reporter gene controlled by the thyroid-specific regulatory sequences showed variable changes in the different cell lines, which were in part statistically significant. The cell line MHhPax8/13 revealed a statistically significant induction of the hTg (P = 0.016) and the hTPO promoter (P = 0.001) but not of the rNIS promoter. In contrast, the cell line MHhpax8/20 showed no statistically significant increase in hTg (1.5-fold) and rNIS (0.8-fold) promoter induction but a 6.3-fold induction of the hTPO promoter (P = 0.01; Fig. 2A). The cell line MHhPax8/3 was characterized by statistically significant activation of the hTPO promoter (P = 0.003) but not of the hTg or rNIS regulatory regions. When compared with the MH3924A wild-type cells, we observed no induction or a low induction of the thyroid-specific regulatory regions in the dTTF-1–expressing cell lines MHdTTF-1/7 and MHTTF-1/11. In contrast, in cell lines expressing both transcription factors (except the MHhPax8/dTTF-1/8 cell line), we observed a statistically significant (P values < 0.001) activation of the hTg promoter (up to 23-fold) and the hTPO promoter (up to 28-fold) and, to a lesser extent, of the rNIS promoter (up to 6-fold; Fig. 2B), indicating that both hPax8 and dTTF-1 are required to induce thyroid-specific gene expression. The cell line MHhPax8dTTF-1/14, showing the highest hTg promoter activity, was selected for in vivo experiments.

Thyroid-specific promoter activity in hPax8- or dTTF-1–expressing MH cell lines (A) and in hPax8/dTTF-1–expressing MH cell lines (B). MH cell lines expressing hPax8 or dTTF-1 and MH3924A cells were plated in 24-well plates and transiently transfected with reporter constructs phTg(2)luc, phTPOluc, prNISluc, and pCMVluc (0.4 μg DNA per well) or mock-transfected 24 h before cells were lysed. Luciferase activity in 20 μL of cell lysate, as determined in a luminometer, is given as relative light units/μg protein normalized with luciferase expression obtained after pCMVluc transfection. Data are mean values and SD (n = 6). *P ≤ 0.05; **P ≤ 0.01.
Imaging of Pax8 and TTF-1 Cooperation by Luciferase Expression in Nude Mice
To demonstrate hPax8 and dTTF-1 activity on thyroid-specific regulatory sequences in vivo, we implanted nude mice in the left or right flank with 6 million transiently transfected (right flank) or mock-transfected (left flank) MH3924A wild-type or MHhPax8dTTF-1/14 cells containing the constructs pCMVluc, phTg(2)luc, prNISluc, and phTPOluc. Immediately after the cell implantation, DNPE-caged Luciferin was injected intraperitoneally and the mice were imaged after 40 min using a cooled CCD-camera. Mice implanted with the phTPOluc- and prNISluc-transfected MHhPax8dTTF-1/14 cells revealed a significantly higher bioluminescence at the right flank compared with those implanted with the equally transfected MH3924A wild-type cells (Fig. 3; Table 1). We also observed a higher signal in animals implanted with phTg(2)luc-transfected MHhPax8dTTF-1/14 cells than in mice implanted with phTg(2)luc-transfected MH3924A wild-type cells. In this case, the luminescence was lower than that of phTPOluc- and prNISluc-transfected MHhPax8dTTF-1/14 cells. A significant luciferase expression was imaged in mice bearing the pCMVluc-transfected cells, and no bioluminescence was detectable in the left flanks implanted by mock-transfected recombinant or wild-type MH3924A cells or other organs of the animals. These results demonstrate that it is possible to image hPax8 and dTTF-1 cooperation in the induction of thyroid-specific promoter activity in living mice.
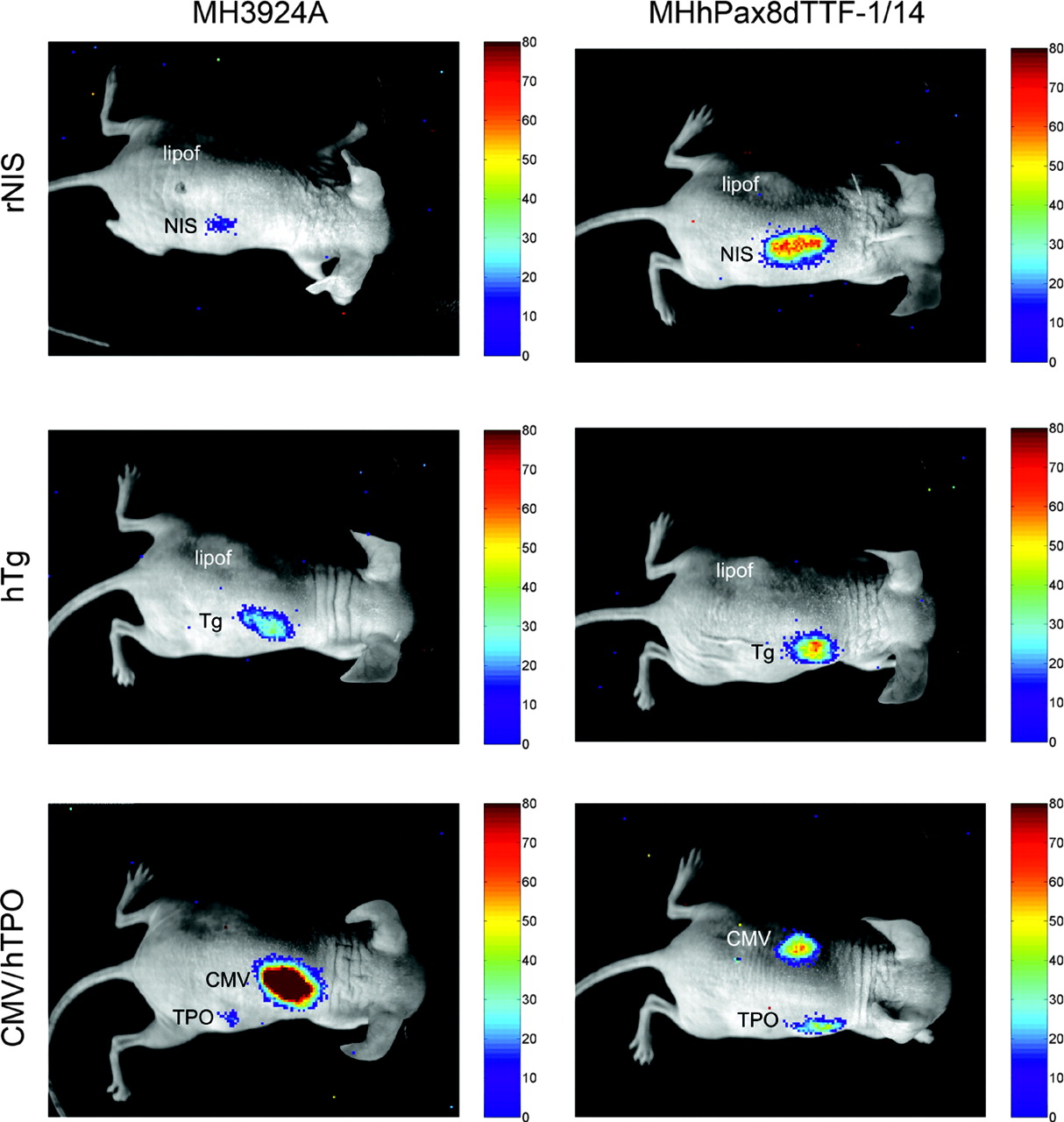
In vivo optical CCD imaging of mice carrying MH3924A and MHhPax8dTTF-1/14 cells transiently transfected with different reporter constructs. All images shown are the visible light image superimposed on the optical CCD bioluminescence image. Mice were imaged in supine position after intraperitoneal injection of DNPE-caged Luciferin. All units are CCD counts measured for 5 min in a constant experimental setup and after subtraction of noise. Animals carrying cells transfected with rNIS and hTg constructs (right flank) also received mock-transfected cells (left flank). Animals implanted with phTPOluc-transfected cells (right flank) were implanted with pCMVluc-transfected cells (left flank). lipof = control; NIS = rNIS promoter/enhancer construct; Tg = hTg promoter/enhancer construct; CMV = CMV promoter; TPO = hTPO enhancer/promoter.
Evaluation of In Vivo Bioluminescence Data
Effect of hPax8 and dTTF-1 on Endogenous Thyroid-Specific Gene Expression
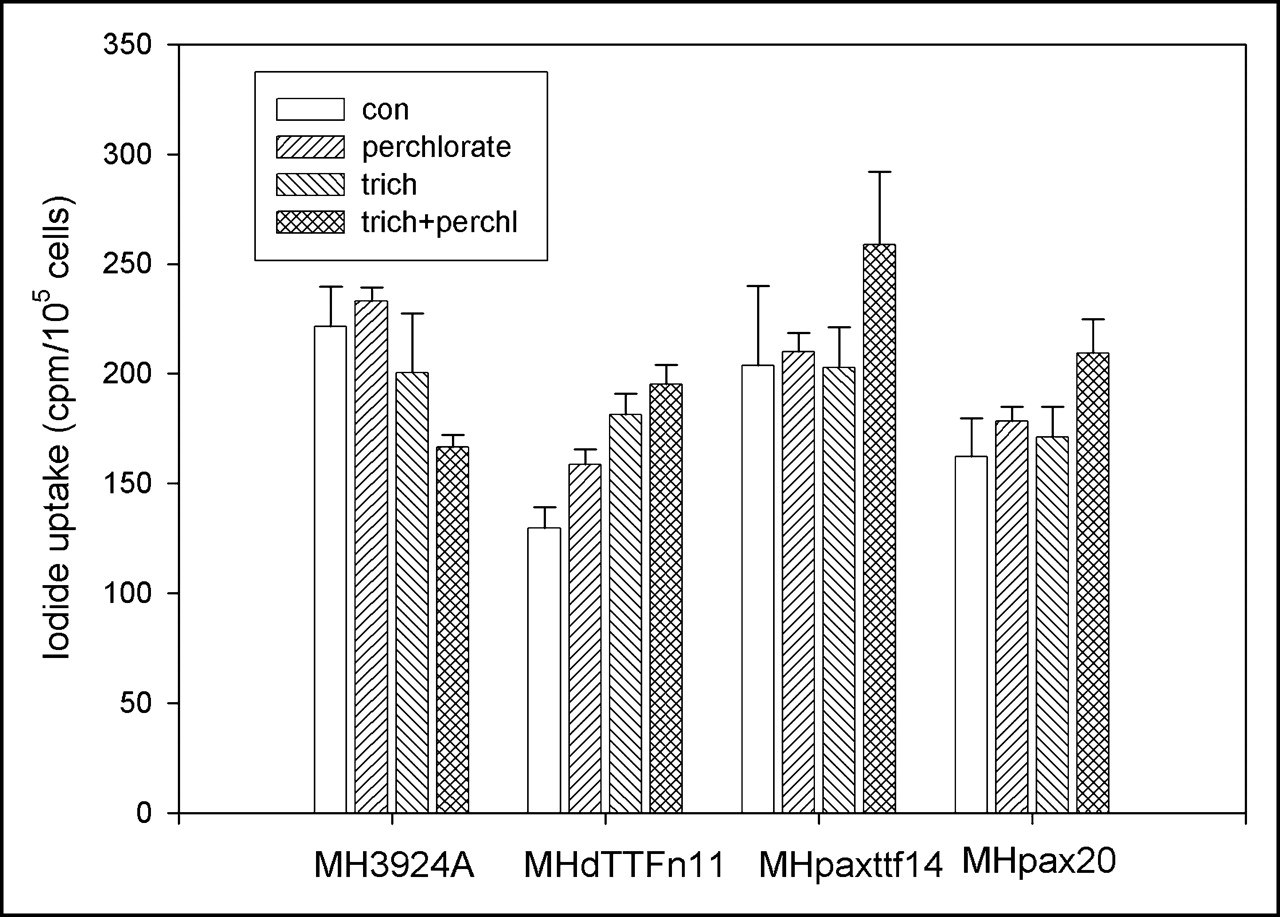
We also investigated the effect of hPax8 or dTTF-1 or both on the transcriptional activation of the endogenous thyroid-specific promoters in recombinant MH cell lines. The expression and functional activity of the NIS gene in these cell lines was analyzed in Na125I uptake experiments. hPax8- or dTTF-1–expressing MH cell lines were cultured in Na125I-containing medium for 4 h with or without sodium perchlorate, which selectively blocks the iodide transporter. In the recombinant and as well as in the wild-type MH3924A cells, only a minimal accumulation of iodide was measured. Because sodium perchlorate did not affect the Na125I uptake, we suggest an influx via diffusion in these cells is more likely than an active iodide transport. Even the addition of the deacetylase inhibitor trichostatin A, which is known to induce chromatin remodeling and to facilitate the access of transcription factors to promoter sequences, did not lead to an increased iodide uptake in any of the cultures (Fig. 4).
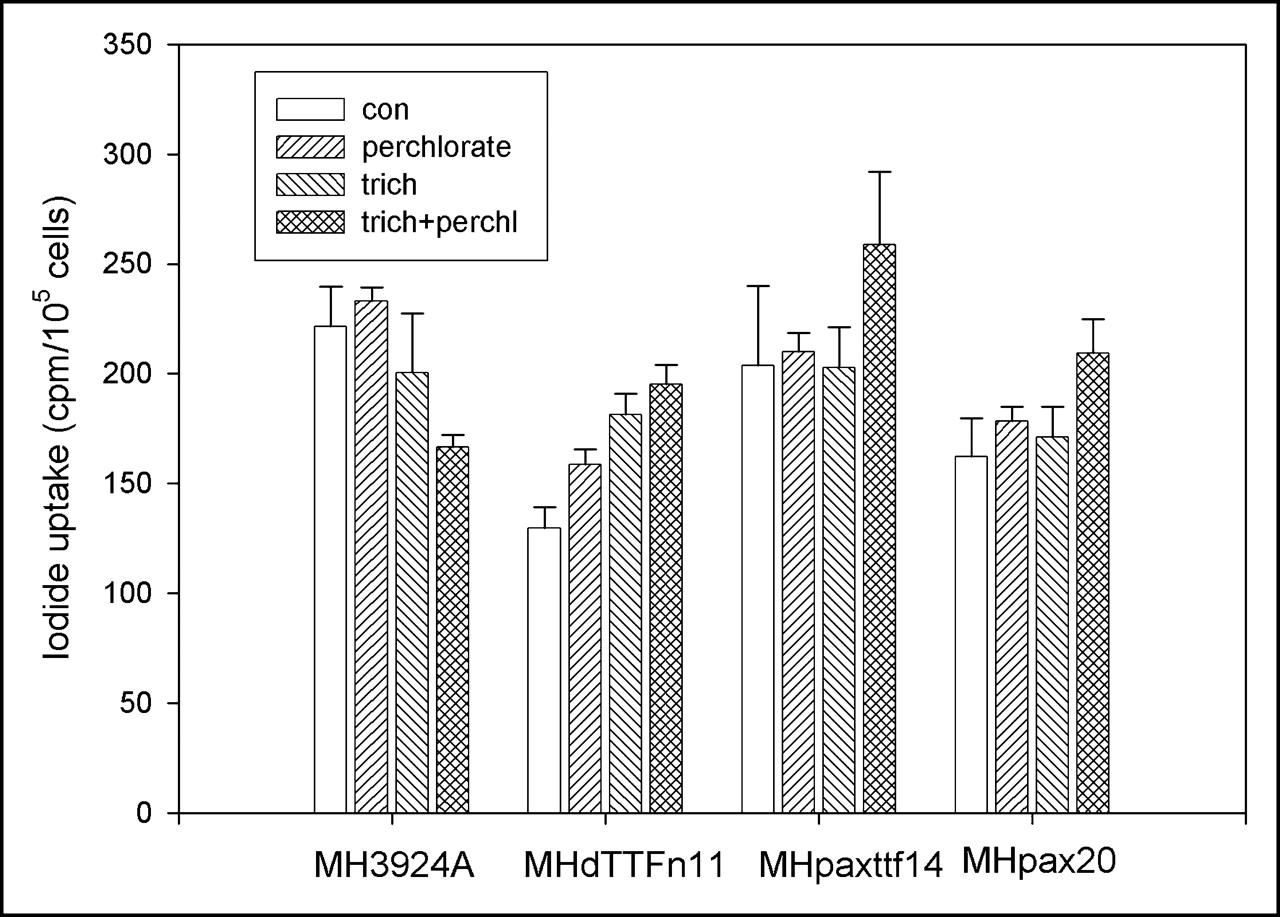
125I uptake in hPax8- and hPax8dTTF-1–expressing cells or MH3924A cells. Recombinant MH cell lines and MH3924A cells were cultured in medium with or without trichostatin A (10 ng/mL for 24 h) before 74 kBq Na125I were added for 4 h. Uptake studies were done in presence or absence of 50 μmol/L sodium perchlorate. Data are mean values and SD (n = 3). con = control; trich = trichostatin A; trich+perchl = trichostatin A and perchlorate.
The inability of the hPax8- and dTTF-1–expressing MH cell lines to accumulate Na125I might be due to the functional inactivity of the NIS or TPO in hepatoma cells or to the lack of thyroid-specific gene transcription. Therefore, we performed a RT-PCR using rTPO-, rTg-, and rNIS-specific primers to study thyroid-specific gene expression in trichostatin A–treated or nontreated MHhPax8dTTF-1/14 cells shown in Figure 5. In contrast to the differentiated thyroid FRTL5 cell line, characterized by TPO, Tg, or NIS gene expression, none of the genes was found to be transcribed in the MHhPax8dTTF-1/14 cell line, pointing to an inaccessibility of the respective genes for the thyroid-specific transcription factors.

RNA from FRTL5 cells and from trichostatin A (10 ng/mL)–treated and nontreated MHhPax8dTTF-1 cells was used to synthezise cDNA that was used for a PCR using specific primers for rTg (Tg), rTPO (TPO), and rNIS (NIS).
DISCUSSION
The aim of this study was to image the functional interaction between the thyroid transcription factors dTTF-1 and hPax8 and their target genes as well as to investigate their ability to induce a thyroid-specific genotype in nonthyroid tumor cells. Pax8 and TTF-1 expression in MH cells presents a useful system to demonstrate functional protein–protein interaction in vivo for 2 reasons. First, the regulatory sequences of the thyroid-specific genes Tg, TPO, and the NIS have been extensively studied and promoter regions responsible for binding of the transcription factors have been identified. Second, hepatoma cells provide an intracellular milieu unrelated to thyroid follicular cells lacking molecules such as the transcription factor TTF-2, which might interfere with Pax8–TTF-1 interactions.
Here, we demonstrate that the dTTF-1 and the hPax8 gene product show a cross-species functional activity in rat MH cells. In cell lines expressing both transcription factors, the hTg, hTPO, and rNIS promoter/enhancer are activated to induce a significant higher luciferase gene expression compared with that of cell lines expressing either hPax8 or dTTF-1 alone. To our knowledge, this is the first report on the functional cooperation between thyroid transcription factors and thyroid-specific genes monitored in nude mice.
Procedures for the noninvasive dynamic in vivo monitoring of protein–protein interactions have been applied using model systems where the proteins were in direct physical contact. Using a CCD camera, Ray et al. adapted the commonly used yeast 2-hybrid technique for in vivo detection of luciferase expression driven by GAL4 and VP16 proteins associated by the interaction of the MyoD and Id proteins (24). Imaging of the interaction between MyoD and Id was also demonstrated by a complementation and a reconstitution strategy using a CCD camera and split reporter constructs of firefly luciferase (11). In this experimental setup, 40%–60% of the luciferase activity was obtained as compared with the transfection of a plasmid bearing the full-length reporter gene (11).
On closer view, not only physical but also functional protein interactions exist, which may be studied by analysis of promoters or promoter modules (25) or by using combinations of specific promoters and reporter genes (26). Recently, the interaction between transcription factors and regulatory elements has been visualized in vitro using individual regulatory promoter and enhancer elements driving reporter genes, such as the secreted alkaline phosphatase (SEAP), which had been cloned downstream of several copies of specific transcription factor–binding sequences (27). In our study, interactions between combinations of 3 proteins were studied: dTTF-1 and hPax8 on one side and Tg, TPO, or NIS on the other side. Tg, TPO, and NIS interact functionally with dTTF-1 and hPax8 because their expression is regulated by the binding of dTTF-1 and hPax8 to the corresponding promoter elements. Furthermore, dTTF-1 and hPax8 interact in the activation of the promoter/enhancer regions, either physically or functionally by recruiting cofactors. In vitro, only a weak induction of the thyroid-specific reporter constructs by either dTTF-1 or hPax8 alone was observed, indicating a weak functional interaction of single transcription factors with these promoter/enhancer elements. In contrast, the cooperation of dTTF-1 and hPax8 resulted in a much stronger luciferase activity in vitro as well as in vivo. Obviously, the regulation of thyroid-specific genes requires a more complex mechanism based on the functional protein–protein interaction between both transcription factors and the target protein. Transfection of wild-type cells with the construct containing the hTg enhancer and promoter elements and the luciferase gene resulted in a higher luminescence compared with transfection with the rNIS or hTPO constructs. This may be due to nonspecific activation, because binding sites for nonspecific factors such as cyclic adenosine monophosphate (cAMP)–responsive elements have been described for the hTg promoter and enhancer sequences (13,28). Since 2 hTg enhancer elements were used to enhance the reporter gene expression, this may result in an increased activation by cAMP or other nonspecific factors.
TTF-1 and Pax8 colocalize in the nucleus of thyroid cells and a direct biochemical interaction of the 2 factors was observed in in vitro as well as in vivo assays (6). According to the current hypothesis, Pax8 and TTF-1 can interact with each other or can act separately or in cooperation with various factors or cofactors. This depends largely on the promoter context within the target gene to regulate thyroid-specific gene expression. However, it is still unclear whether Pax8 and TTF-1 physically interact during induction of Tg and TPO, since their binding sites within the promoters overlap (7).
The features described here are different from the properties of the 5′-flanking region of the NIS gene, where 2 TTF-1–binding sites have been localized within the promoter (29) within 0.56 kb upstream of the translation start and 2 Pax8-binding sites within the NIS upstream enhancer (8). Obviously, in this case, interaction of the 3 proteins dTTF-1, hPax8, and NIS occurs functionally without any physical interaction between the transcription factors. Individual cell lines expressing both thyroid transcription factors showed varying levels of thyroid-specific promoter induction. This may be due to variations in the dTTF-1 or hPax8 protein concentration. Since the integration of the retroviral genome occurs randomly at different sites of the cellular DNA, the genes coding for dTTF-1 and hPax8 may be expressed in variable amounts. Alternatively, this effect may be caused by different gene copy numbers. Therefore, the ratio between dTTF-1 and hPax8 may vary in the different cell lines, leading to the observed differences in luciferase expression.
Recently, it has been shown that transfection of TTF-1 in the Pax8-expressing rat thyroid tumor cell line FRT induced activation of the endogenous Tg promoter, although to a very low extent (30). Pasca di Magliano et al. transfected the Pax8 gene into the transformed thyroid cell line PCPy and, thereby, restored the endogenous Tg, TPO, and, to a lesser extent, NIS expression at wild-type levels (31).
In the present work, we performed Na125I uptake experiments as well as RT-PCR to investigate the endogenous expression of Tg, TPO, or NIS in hPax8- and dTTF-1–transfected MH cells, without evidence for the existence of a functionally active iodide transport molecule or thyroid-specific transcripts. There might be several reasons to account for this. First, it seems to be difficult to reproduce the appropriate concentration of the transfected protein in a cell transfection system. It has been shown for several regulating proteins that a small change in their expression may have quite large biologic effects (32,33). In addition, there is evidence that TTF-1 and Pax8 cooperate with several ubiquitous transcription factors, forming complexes on the regulatory region of specific genes (34–36). Since the transcription factors were shown to be functionally active in our experiments, it is possible that cofactors required for thyroid-specific gene expression are not provided by MH cells but by thyroid cells.
Alternatively, the endogenous thyroid-specific regulatory regions may be inaccessible to Pax8 and TTF-1. The appropriate regulation of gene expression requires the interplay of complexes that remodel chromatin structure and, thereby, regulate the accessibility of individual genes to sequence-specific transcription factors and the basal transcription machinery. In hepatoma cells, thyroid-specific genes may be packed in a more condensed chromatin structure. It is generally accepted that transcriptional activity is correlated with histone acetylation. According to one model of transcription, the expression of genes depends primarily on their ability to recruit either histone acetylase or histone deacetylases (HDACs) to the promoter. Transcriptional repression results from the activity of HDACs removing acetyl groups from nucleosomes, causing the tightening of nucleosomal integrity and a suppression of transcription (37). Therefore, we investigated whether transcriptional activation of thyroid-specific genes in hPax8- and dTTF-1–expressing cells is induced by the HDAC inhibitor trichostatin A, which is known to induce the accumulation of acetylated histone H4 (38,39). According to our 125I-uptake experiments and RT-PCR performed with trichostatin A–treated hPax8- and dTTF-1–expressing cells, there was no evidence for a functionally active iodide transport protein or for transcription of the genes for Tg, TPO, or NIS. Our findings suggest that the inhibition of deacetylation per se is not sufficient to enable the access of the thyroid-specific transcription factors to the Tg, TPO, or NIS regulatory regions. Alternatively, MH cells may not be sensitive toward trichostatin A. However, in separate experiments, we observed a dose-dependent decrease in cell number after exposure to trichostatin A.
CONCLUSION
The transduction of genes encoding the thyroid transcription factors hPax8 and dTTF-1 in MH cells leads to activation of thyroid-specific promoter/enhancer elements in vitro and in vivo, demonstrating the functional interaction between TTF-1, hPax8, and Tg, TPO, or NIS. The functional activity of the transcription factors per se, however, is not sufficient to induce the expression of endogenous thyroid-specific genes in nonthyroid tumor cells. With regard to the development of therapeutic procedures based on the accumulation of radioiodide in tumors, the expression of genes involved in the uptake as well as the organification of iodide is required. Studies investigating the effect of chromatin- remodeling factors on endogenous gene expression in Pax8- and TTF-1–expressing cells are in progress.
Acknowledgments
The authors thank Ursula Schierbaum and Karin Leotta for their help in performing the animal experiments. This study was supported by a grant from the Tumorzentrum Heidelberg/Mannheim.
Footnotes
Received Oct. 10, 2004; revision accepted Jan. 5, 2005.
For correspondence or reprints contact: Annette Altmann, PhD, Clinical Cooperation Unit of Nuclear Medicine, German Cancer Research Center, Im Neuenheimer Feld 280, 69120 Heidelberg, Germany.
E-mail: a.altmann{at}dkfz.de
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Upregulation of thyroid transcription factor-1 and human leukocyte antigen class I in Hashimoto's disease providing a clinical evidence for possible triggering autoimmune reaction
- Expression of sodium-iodide symporter mRNA in the thyroid gland of Xenopus laevis tadpoles: developmental expression, effects of antithyroidal compounds, and regulation by TSH