


Abstract
99mTc-Annexin V is used to image cell death in vivo via high-affinity binding to exposed phosphatidylserine. We investigated how changes in membrane-binding affinity, molecular charge, and method of labeling affected its biodistribution in normal mice and its uptake in apoptotic tissues. Methods: An endogenous Tc chelation site (Ala-Gly-Gly-Cys-Gly-His) was added to the N-terminus of annexin V to create annexin V-128. The membrane-binding affinity of annexin V-128 was then progressively reduced by 1–4 mutations in calcium-binding sites. In addition, mutations were made in other residues that altered molecular charge without altering membrane-binding affinity. All mutant proteins were labeled with 99mTc at the same N-terminal endogenous chelation site. Wild-type annexin V was also labeled with 99mTc after derivatization with hydrazinonicotinamide (HYNIC). Radiolabeled proteins were tested for biodistribution in normal mice and in mice treated to induce apoptosis of the liver. Results: Comparison of 99mTc-annexin V-128 with 99mTc-HYNIC-annexin V showed that the protein labeled at the endogenous chelation site had the same or higher uptake in apoptotic tissues, while showing 88% lower renal uptake at 60 min after injection. The blood clearance of annexin V was unaffected by changes in either the membrane-binding affinity or the molecular charge. Kidney uptake was unaffected by changes in binding affinity. In marked contrast, uptake in normal liver and spleen decreased markedly as affinity decreased. The same pattern was observed in animals treated with cycloheximide to induce apoptosis. Control experiments with charge mutants showed that the effects seen with the affinity mutants were not due to the concomitant change in molecular charge that occurs in these mutants. Conclusion: (a) All four domains of annexin V are required for optimal uptake in apoptotic tissues; molecules with only 1 or 2 active domains are unlikely to be suitable for imaging of cell death in vivo. (b) Uptake in normal liver and spleen is specific (dependent on phosphatidylserine-binding affinity), whereas renal uptake is nonspecific. (c) 99mTc-Annexin V-128 detects cell death as well as 99mTc-HYNIC-annexin V, while showing 88% less renal retention of radioactivity due to much more rapid urinary excretion of radioactivity.
Apoptosis is important in normal physiology and numerous disease states. We have developed annexin V as a means to image cell death in vivo (1,2). This is based on the protein’s high affinity for phosphatidylserine (PS) exposed on the extracellular face of cell membranes (3), which occurs when cells undergo apoptosis (4). Annexin V contains 4 similar domains of 70–80 amino acids arranged in a compact water-soluble structure (5). High-affinity membrane binding depends on multiple calcium-binding sites present in all 4 domains of the molecule (6). Clinical applications for annexin V imaging are being developed in oncology (7), organ transplantation (8), and cardiovascular disease (9).
Although 99mTc-HYNIC-annexin V (HYNIC is hydrazinonicotinamide) has been successfully used in many studies, it has certain limitations—most notably, its very high uptake in the kidney, which results in high radiation dose and may limit imaging in the abdomen (10). Alternative approaches for radiolabeling annexin V with technetium have been described (11–14). Annexin V has also been labeled with 18F for PET (15). Although there is some variation in biodistribution and metabolism of these different radiolabeled annexins, many properties of the protein are the same regardless of the method of labeling. Initial blood clearance is relatively rapid (11,16,17). Organ uptake is almost always highest in the kidney, followed by the liver; there is moderate uptake in the spleen, lung, and bowel; uptake is very low in the brain, heart, and skeletal muscle. Recently, novel approaches based on modifications of the protein itself have also been described. There has been a report of a single-domain “mini-annexin” that may be useful for imaging (18). Annexin V has also been modified with polyethylene glycol to prolong its circulation time and, thereby, improve its availability over extended periods for uptake by tumors (19).
As part of an effort to develop improved annexin V–based imaging agents, we wanted to address several questions. First, why do some organs have much higher uptake of annexin V than others? Second, what structural features of annexin V control uptake in both normal and apoptotic tissues? Third, how much loss of membrane-binding affinity can be tolerated before annexin V loses its ability to localize in apoptotic tissues? To address these questions, we prepared a series of site-directed mutants of annexin V that alter properties such as charge and membrane-binding affinity. We took advantage of the method we previously developed for 99mTc labeling of annexin V via an N-terminal endogenous chelation site consisting of the amino acid sequence Ala-Gly-Gly-Cys-Gly-His (12). All structural mutants of annexin V contained the same sequence in 1:1 stoichiometry; therefore, the site of 99mTc labeling was exactly the same for all mutant proteins. Thus, any differences observed could be attributed to the specific mutations introduced rather than differences in the site or degree of labeling. We show that the new annexin V-128 recombinant protein has substantially improved biodistribution properties compared with HYNIC-derivatized annexin V. We also show that alterations in membrane-binding affinity change biodistribution in normal animals and target uptake in an animal model of apoptosis. These results provide guidance in the development of improved annexin V–based apoptosis imaging agents radiolabeled with either 99mTc or positron emitters as well as annexin V–based agents used for fluorescence or MRI.
MATERIALS AND METHODS
Preparation and 99mTc Labeling of Mutant Proteins
Recombinant human annexin V-128 and derivative mutant proteins were prepared and purified as described (6). In preparation for technetium labeling, protein (0.6 mg) was diluted with phosphate-buffered saline ([PBS] containing 10 mmol/L sodium phosphate, pH 7.4, 140 mmol/L NaCl) to a final volume of 0.2 mL and reduced with 1 mmol/L dithiothreitol at 37°C for 15 min under anaerobic conditions. The protein was applied to a Sephadex G-25 column (NAP-5; Pharmacia), which had been preequilibrated with deoxygenated buffer (20 mmol/L trisodium citrate, pH 5.4, 100 mmol/L NaCl), and the breakthrough (0.2 mL) was discarded. Elution was performed with the same buffer; the first 0.5 mL of eluate was discarded, and the subsequent 0.5 mL, containing the protein, was collected in argon-flushed glass vials. Aliquots of 0.1 mL (containing 0.1 mg protein, 0.59 mg trisodium citrate, and 0.58 mg NaCl) were stored frozen at −20°C in glass vials with Teflon-sealed screw caps (DuPont). Tin reagent was prepared as described (12). Each vial of lyophilized reagent contained 5 mg of sodium glucoheptonate, 0.128 mg SnCl2·2H2O, and 0.128 mg of sodium gentisate. Just before use, the vial was reconstituted with 0.2 mL of deoxygenated water.
99mTc-Annexin V-128 was prepared as follows. 99mTcO4 (185–370 GBq [5–10 mCi] in 0.1 mL of 0.9% NaCl) was added to a polypropylene microfuge tube, followed by 0.005 mL of tin reagent and 0.05 mL (0.05 mg) of reduced protein. After a 60-min anaerobic incubation at 37°C, the reaction mixture was brought to a volume of 0.4 mL with PBS, pH 7.4, and applied to a Sephadex G-25 column (NAP-5; Pharmacia) equilibrated and eluted with PBS, pH 7.4. The labeled protein was collected in a volume of 1.4 mL (0.4 mL of breakthrough and the first 1.0 mL of eluate). The percentage incorporation of technetium determined by instant thin-layer chromatography (ITLC) was routinely 85%–97%, corresponding to a specific activity of about 3.7 GBq/mg (100 mCi/mg). The labeling reaction can also be performed at room temperature for 20–30 min with minimal loss of labeling efficiency (12). A dose of 1.85–7.4 MBq of tracer (0.05–0.2 mCi; 0.1–0.2 mL, 0.05–0.1 mg protein/kg body weight) was injected via tail vein for all animal studies. 99mTc-HYNIC-annexin V was prepared and labeled with the tin-tricine method as previously described (1).
Membrane-Binding Affinity Measurement by Quantitative Calcium Titration
The affinity of labeled protein for red blood cells (RBCs) with exposed PS was determined by the calcium titration assay previously described for fluorescently labeled annexin V (20). Annexin V-128 was labeled with 99mTc as described and diluted to a working concentration of 200 nmol/L in 50 mmol/L N-(2-hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) (HEPES)-sodium, pH 7.4, 100 mmol/L NaCl, 3 mmol/L NaN3, and 5 mg/mL bovine serum albumin (BSA). The final concentration of 99mTc-annexin V-128 in the assay was 1 nmol/L, RBCs were added at 1 × 108 per milliliter and calcium was varied from 0 to 5 mmol/L, in an assay buffer containing 50 mmol/L HEPES-sodium, pH 7.4, 100 mmol/L NaCl, 3 mmol/L NaN3, with 1 mg/mL BSA as carrier protein. After an 8-min incubation at room temperature, cells were pelleted by centrifugation, resuspended in 1 mL of assay buffer containing the same concentration of calcium used in the binding step, and centrifuged again. The supernatant was removed and the 99mTc-annexin V-128 bound to the RBCs was released with 0.7 mL of assay buffer containing 5 mmol/L ethylenediaminetetraacetic acid. The RBCs were pelleted by centrifugation and the amount of 99mTc-annexin V-128 bound to the RBCs at each calcium concentration was determined by counting an aliquot (0.3–0.6 mL) of the supernatant on a γ-counter (RIASTAR Jr; Hewlett Packard). Calcium titration curves were analyzed to determine the binding affinity as described (20); results are expressed as a pK value, which is the logarithm of the equilibrium constant for protein-membrane-calcium binding.
TUNEL (Terminal Deoxyribonucleotidyl Transferase-Mediated Deoxyuridine Nick-End Labeling) Assay
Livers from normal and cycloheximide-treated mice were evaluated for the presence of apoptotic nuclei in formalin-fixed, paraffin-embedded tissue sections with a fluorescent TUNEL assay kit following the manufacturer’s directions (Oncogene Research Products Fluorescein-FragEL kit; Calbiochem-Novabiochem Corp.). The number of apoptotic nuclei was determined from 60 fields per slide, containing an average of 36 nuclei per field, examined under 60× magnification.
Blood Clearance and Biodistribution Studies in Normal and Treated Mice
All mice used were adults (6- to 8-wk old) of the BALB/c strain obtained from the breeding facility of the Department of Comparative Medicine, Stanford University. Hepatic apoptosis was induced by intraperitoneal injection of cycloheximide (21) at a dose of 50 mg/kg dissolved in 0.5 mL of PBS. All animals were anesthetized with a cocktail of ketamine (100 mg/kg intraperitoneally) (Fort Dodge Animal Health) and xylazine (10 mg/kg, intraperitoneally) (Butler) before clearance, biodistribution, and imaging studies. Blood clearance studies were performed by serial retroorbital blood samples obtained after tail vein injection of tracer. Biodistribution assay was performed immediately after sacrifice. Tissue and organ samples were analyzed with a scintillation well counter along with 4 samples of standard activity (1/100 of injected dose) at an energy level of 140 keV with an energy window of ±20 keV. Results are expressed as the average of the percentage injected dose (%ID) or %ID per gram (g) of tissue (% ID/g) ± 1 SD of the mean.
Dynamic Imaging
Dynamic planar images of tracer activity were obtained using the A-SPECT dedicated small animal imaging SPECT γ-camera (Gamma-Medical Instruments) with a 128 × 128 matrix and a high-resolution, high-sensitivity, parallel-hole collimator. Sedated animals were placed in the anterior supine position and planar whole-body images were collected starting immediately after injection of tracer via tail vein (animals received the same dose of tracer used for static biodistribution studies); serial 1-min images were obtained for 35 min, and serial 3-min images were obtained for another 60 min. Region-of-interest image analyses were performed on planar images using commercially available software (Mirage Software, version 5.3; Segami Corp.). The results of the dynamic image analysis for the activity of each region of interest expressed as counts/pixel are reported as the percentage of whole-body activity also expressed as counts/pixel.
RESULTS
Characterization of 99mTc-Annexin V-128
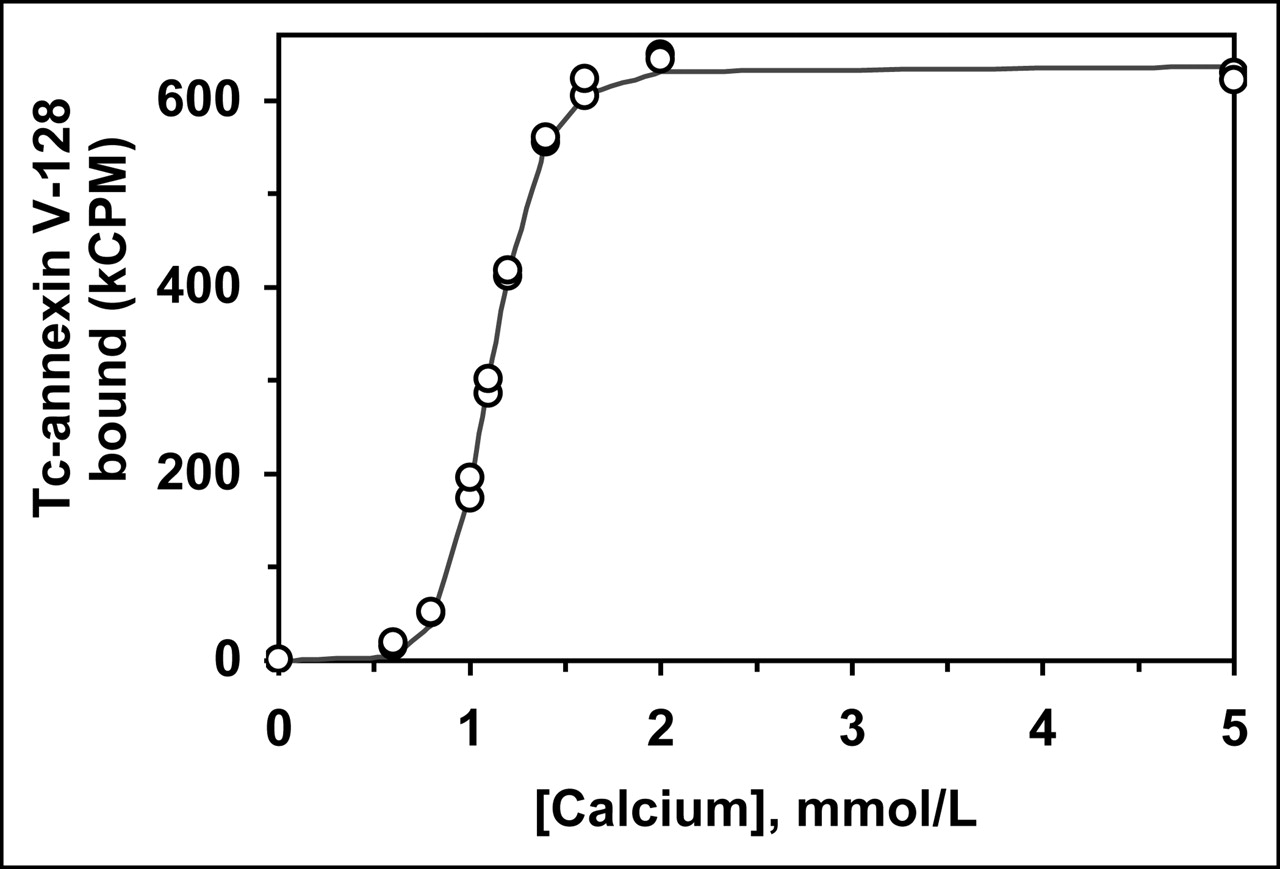
All mutant proteins were built on the base structure of annexin V-128 (6). This protein has essentially the same structure as wild-type human annexin V, except for an N-terminal extension of Ala-Gly-Gly-Cys-Gly-His to provide a site for Tc chelation and a Cys316Ser point mutation. Figure 1 shows a calcium titration curve performed with 99mTc-annexin V-128 and RBCs with exposed PS. The curve is identical to that previously obtained with fluorescently labeled annexin V-128. The pK value for binding of 99mTc-annexin V-128 to RBCs is 30.8 ± 1.0 (mean ± SD, n = 3), identical to that previously reported for both for wild-type annexin V and for annexin V-128 labeled with fluorescein (6,20). Thus, 99mTc labeling at the N-terminus does not alter binding affinity, consistent with our earlier work with the annexin V-117 protein (12), which has a nearly identical structure.
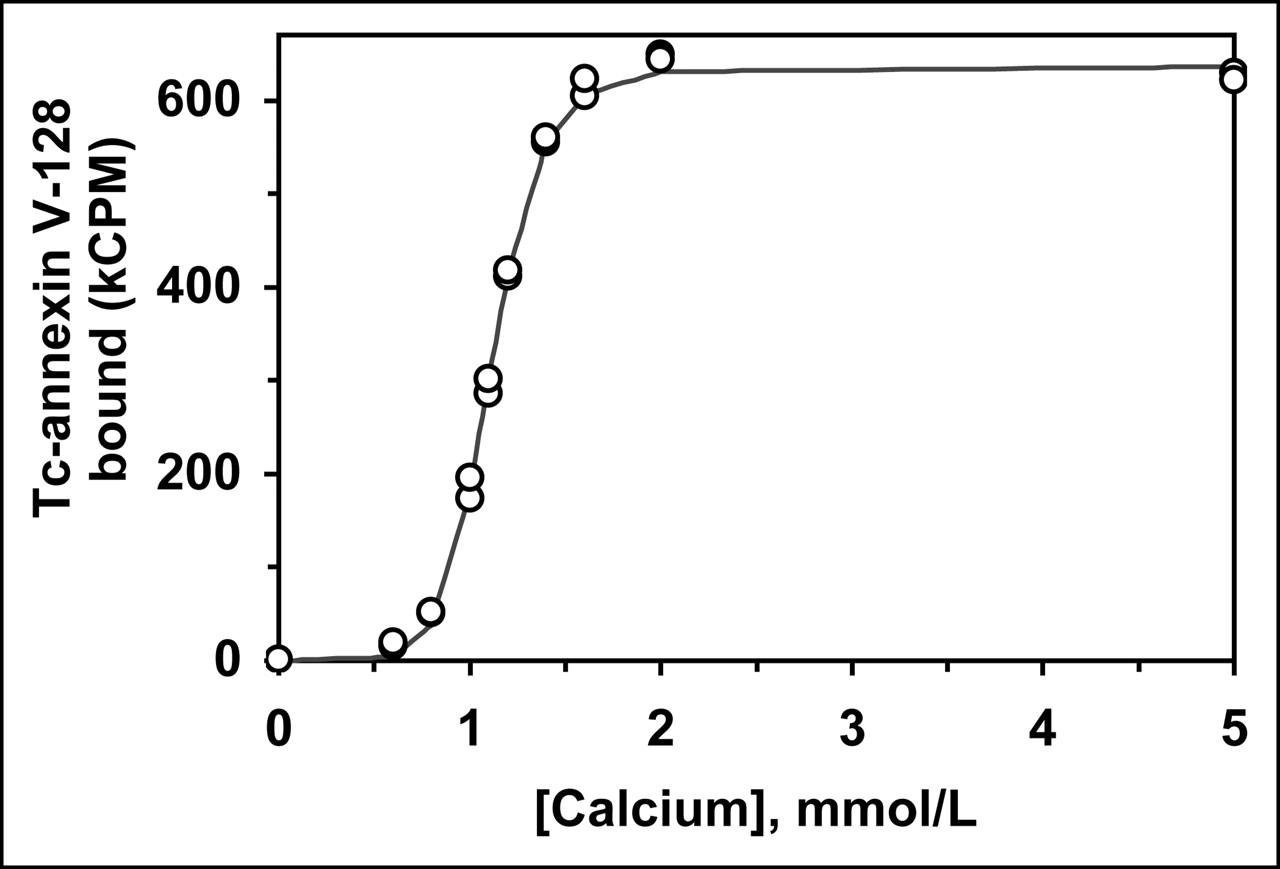
Calcium titration of 99mTc-annexin V-128 binding to RBCs with exposed PS. Line is the fitted function used to determine the median effective concentration (EC50) and Hill coefficient values, from which logarithm of the binding constant (pK value) is calculated (20).
Comparison of HYNIC-Annexin V and Annexin V-128
To verify that 99mTc-annexin V-128 was suitable for detection of cell death, it was compared with 99mTc-HYNIC-annexin V in normal mice. As seen in Table 1, the biodistribution of the 2 proteins was similar overall, with a few exceptions. The most striking exception was the kidney: 99mTc-annexin V-128 showed 88% less renal uptake than 99mTc-HYNIC-annexin V. Liver and spleen uptake was also somewhat lower for 99mTc-annexin V-128, possibly reflecting less aggregated material. Small bowel was the only organ showing higher uptake of 99mTc-annexin V-128 (3.9 %ID) compared with 99mTc-HYNIC-annexin V (1.6 %ID). However, this was greatly outweighed by the lower overall abdominal uptake of 99mTc-annexin V-128 (17 %ID vs. 60 %ID for 99mTc-HYNIC-annexin V).
Biodistribution of 99mTc-HYNIC-Annexin V and 99mTc-Annexin V-128
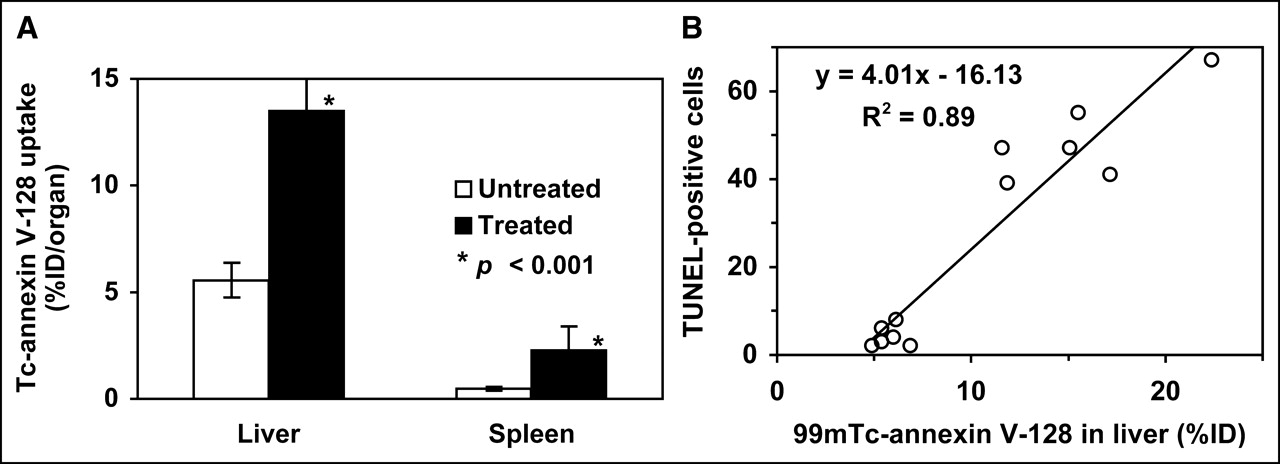
When mice were treated with cycloheximide to induce apoptosis, the liver and spleen showed a severalfold increase in uptake of 99mTc-annexin V-128 (Fig. 2A). We also verified that uptake of 99mTc-annexin V-128 was consistent with an independent measure of apoptosis. As shown in Figure 2B, in vivo uptake of 99mTc-annexin V-128 in the liver correlated closely with histologic analysis of the degree of apoptosis as judged by the TUNEL assay.

(A) Uptake of 99mTc-annexin V-128 in liver and spleen of mice treated with cycloheximide. Groups of 6 mice were either untreated or treated with cycloheximide (50 mg/kg, intraperitoneally). Two hours later, 99mTc-annexin V-128 was injected via tail vein, and organs were harvested 60 min after injection. Significance of the difference between treated and untreated animals was assessed by a 2-tailed t test assuming unequal variances; error bars indicate SD. (B) Correlation between TUNEL positivity and 99mTc-annexin V-128 uptake in liver for treated and untreated mice (6 in each group).
Dynamic Imaging with HYNIC-Annexin V and Annexin V-128
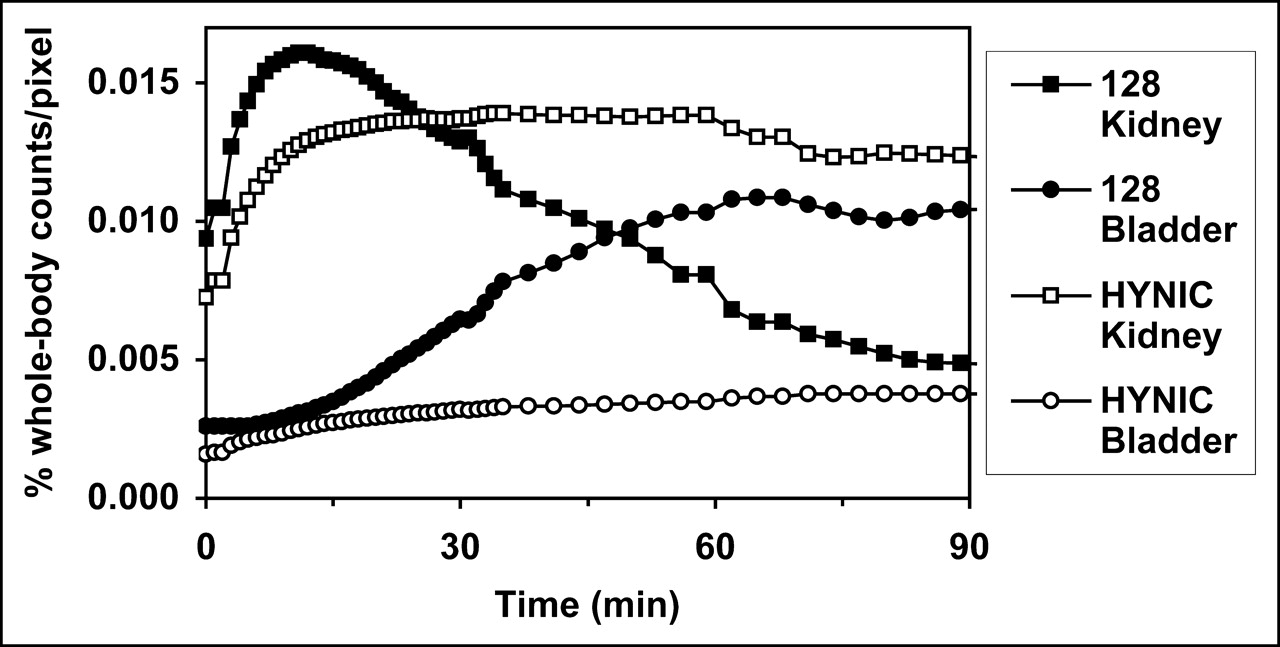
The reason for the striking difference in renal uptake of 99mTc-annexin V-128 compared with that of 99mTc-HYNIC-annexin V was further explored via dynamic imaging studies (Fig. 3). Renal uptake of each protein is comparable up to about 15 min after injection. After that time, there is a progressive decline in renal radioactivity for the 99mTc-annexin V-128 protein, accompanied by a corresponding increase in radioactivity in the bladder. In contrast, for 99mTc-HYNIC-annexin V, radioactivity in the kidney stays roughly constant, and there is minimal bladder excretion during this time. These results were confirmed by static biodistribution measurements performed at 15 min after injection, which confirmed that the renal uptake of 99mTc-annexin V-128 and 99mTc-HYNIC-annexin V was comparable at that time (data not shown).
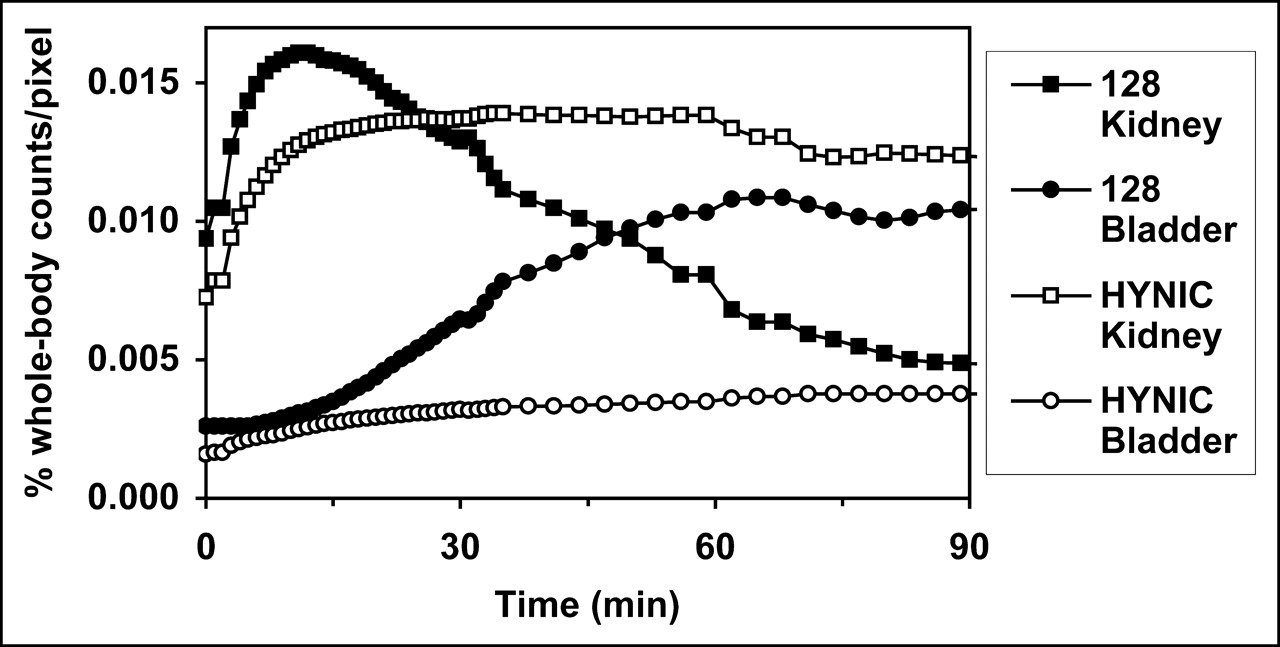
Serial dynamic imaging of kidney and bladder uptake of 99mTc-annexin V-128 and 99mTc-HYNIC-annexin V. Each curve is mean of 4 animals; coefficient of variation at each point is approximately 20%.
Characterization of Charge and Affinity Mutants
To determine how PS-binding affinity and molecular charge affect biodistribution and target uptake of annexin V, a series of mutants were prepared (Fig. 4; Table 2). The PS-binding affinity of annexin V can be reduced in a step-wise fashion by making point mutations in aspartate or glutamate residues contributing to the AB-helix calcium-binding sites present once in each of the 4 domains (6). These mutants span a very broad range of membrane-binding affinity (as summarized on a logarithmic scale by the pK affinity value) but they all have essentially the same molecular mass as the parent protein, since each mutation changes molecular mass by only 1 Da. Since mutations in the AB-helix calcium-binding sites also alter molecular charge by converting a negatively charged residue to a neutral residue, a second set of mutant proteins was prepared with unaltered affinity but altered molecular charge as control proteins to evaluate the specificity of changes observed with the affinity mutants. Some of these mutant proteins have been previously reported (6), and others were developed specifically for this study.

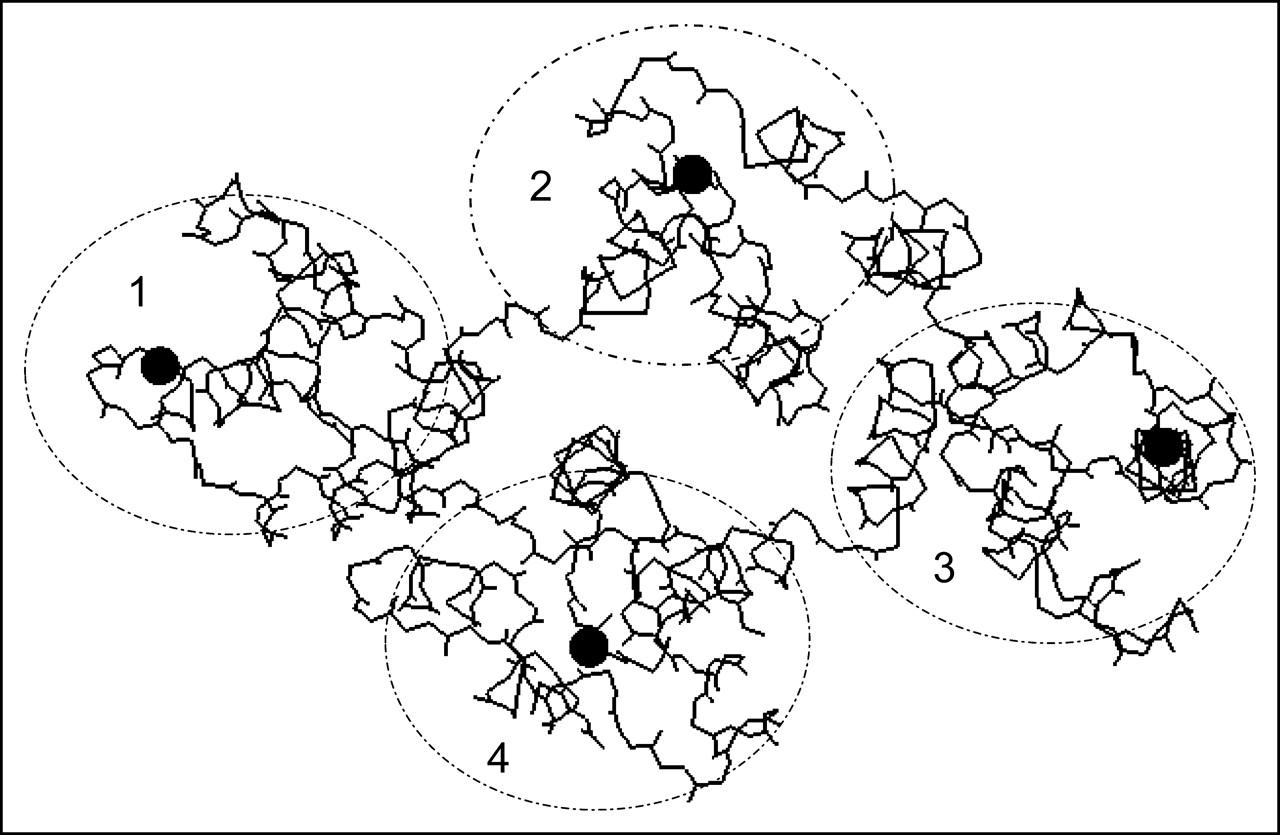
Schematic view of 3-dimensional structure of annexin V-128. View is downward onto membrane-binding face of protein. The 4 domains of protein are numbered, and their approximate locations are indicated by ellipses. Black spheres indicate approximate locations of calcium ions bound to AB-helix calcium-binding sites that have been mutated in this study. Polypeptide backbone is shown as a continuous black line. Location of N-terminal technetium chelation site cannot be seen in this projection because it is on opposite face of molecule, beneath the plane of the page. Structure is based on preliminary coordinates provided by Dr. Barbara Seaton (personal communication, May 2004).
Summary of Affinity and Charge Mutants
Blood Clearance of Affinity Mutants
The blood clearance of 99mTc-annexin V-128 and selected affinity mutants was measured in mice (Fig. 5). All proteins showed the same rapid clearance kinetics, with only 4% remaining in blood at 60 min. These clearance curves are essentially the same as those previously reported for unlabeled annexin V (16), 125I-annexin V and 123I-annexin V (11,17), and HYNIC-annexin V (22). Thus, changes in PS-binding affinity do not alter the kinetics of blood clearance of annexin V. Similarly, changes in molecular charge did not alter blood clearance (data not shown).
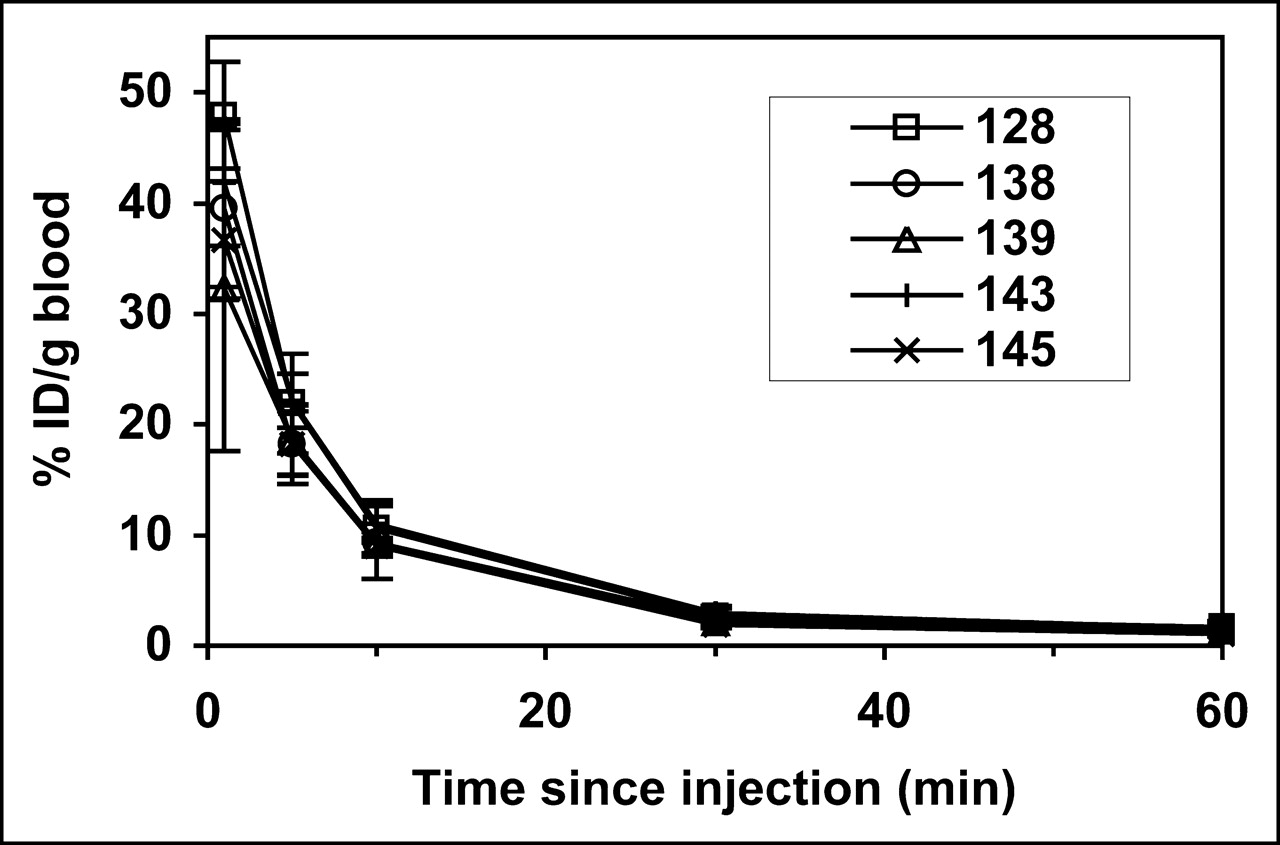
Blood clearance of affinity mutants. Groups of 4 mice received intravenous injections of 99mTc-labeled protein at time zero. Results are given as mean ± SD for each mutant.
Uptake in Apoptotic Organs
To determine the impact of affinity changes on uptake in apoptotic organs, we tested the series of affinity mutants in mice treated with cycloheximide to induce apoptosis of the liver and spleen (Fig. 6A). There was a clear dose–response relationship between the number of mutations and the reduction in organ uptake in the liver and spleen. A single mutation decreased uptake by 50%–60%, and the presence of 2 or more mutations reduced uptake by 80%–90%. However, the specific location of the mutation in the molecule (Fig. 4) did not influence the results: Single-site mutants in domains 1 and 4 were equivalent, double mutants in domains 1 + 4 and domains 2 + 3 were equivalent, and triple mutants in domains 1 + 2 + 4 and domains 2 + 3 + 4 were equivalent. Thus, the dominant factor determining the uptake in apoptotic tissue is the total number of active domains rather than the specific site of the mutation(s); in other words, membrane-binding sites in each of the 4 domains are equivalent in their influence on target uptake in vivo.

(A) Dose–response relationship between number of affinity-reducing mutations and liver and spleen uptake in mice treated with cycloheximide. Each affinity mutant in Table 2 was tested in groups of 4 or 5 mice. Mean values are given for liver and spleen uptake, normalized to uptake observed with wild-type protein. Mutants tested were 128 (wild-type/zero mutations); 131 and 138 (1 mutation); 136 and 139 (2 mutations); 137 and 143 (3 mutations); and 145 (4 mutations). (B) Dose–response relationship between number of affinity-reducing mutations and liver and spleen uptake in normal mice.
As a control, we also determined the effect of affinity mutations on the uptake in normal liver and spleen (Fig. 6B). Somewhat unexpectedly, a similar pattern was observed: Organ uptake was progressively reduced by an increasing number of mutations. To verify that this effect on normal organ uptake was not an artifact of concomitant changes in molecular charge that occur in the affinity mutants, a second series of mutant proteins was tested that altered charge without affecting PS-binding affinity (Fig. 7). Charge differences from −2 to +2 relative to wild-type protein did not affect the uptake in normal liver or spleen, indicating that the effects observed in Figure 8 were due to affinity changes rather than changes in charge per se.
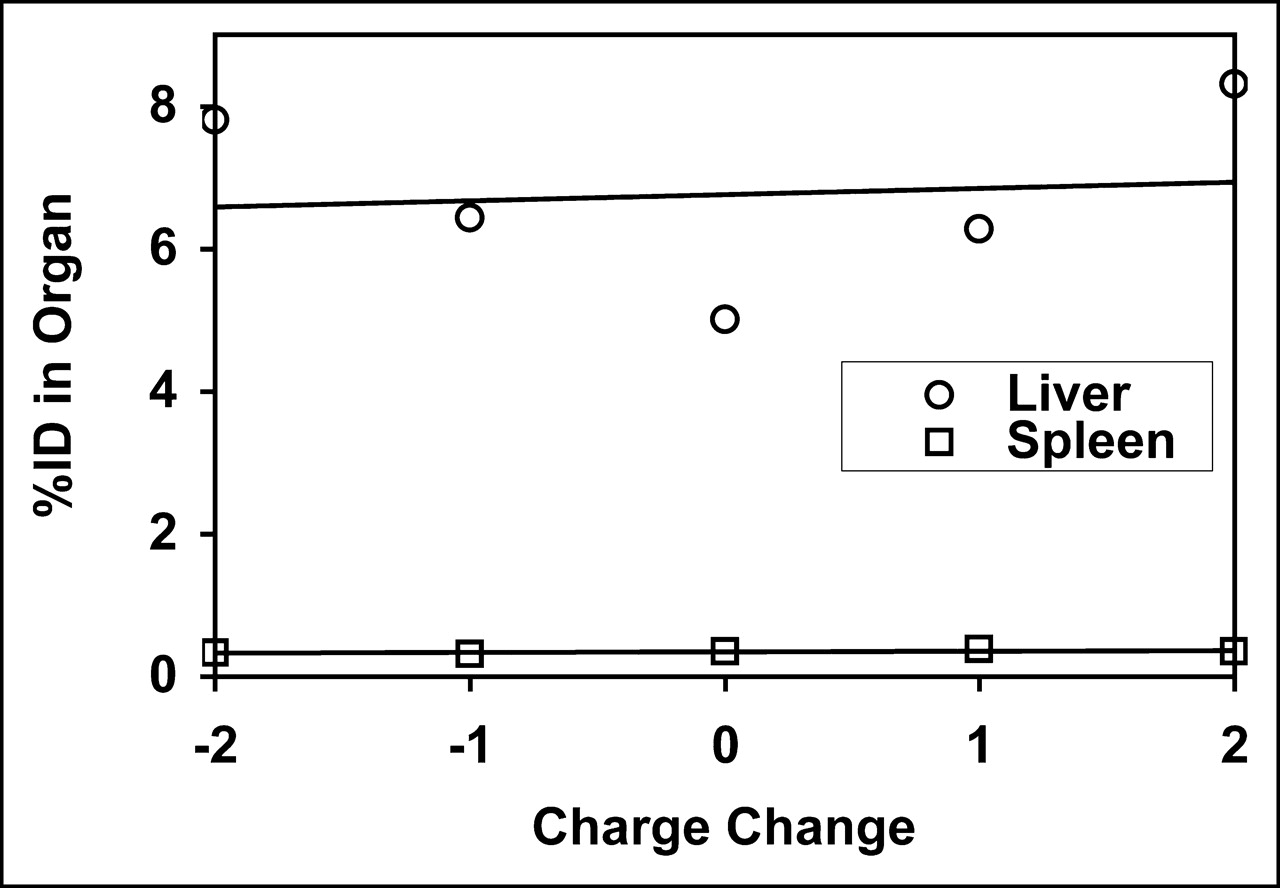
Effect of molecular charge on liver and spleen uptake of 99mTc-annexin V-128 in normal mice. Each charge mutant in Table 2 was tested in groups of 4 or 5 mice. Mean values are given for liver and spleen uptake.
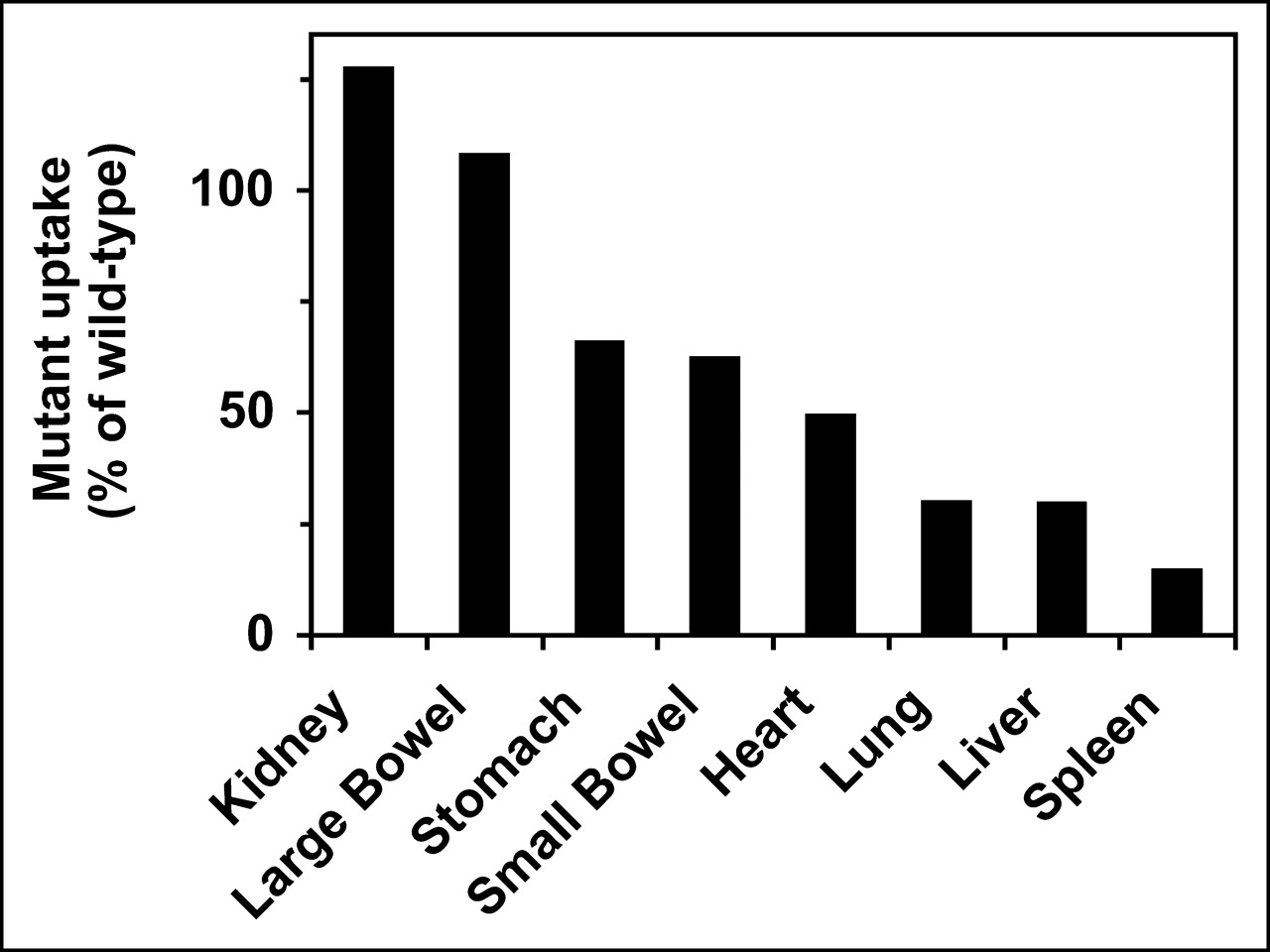
Organ uptake of affinity mutants in normal mice. Triple and quadruple mutants in Table 2 (annexin V-137, -143, and -145) were tested in groups of 4 or 5 mice for each mutant. Results were averaged for all 3 mutant proteins and expressed as percentage of organ uptake observed with wild-type protein (annexin V-128).
In view of the results obtained with normal liver and spleen, we performed a more extensive evaluation of the effect of PS-binding affinity on biodistribution in normal animals (Fig. 8). Biodistribution was measured with triple and quadruple mutant proteins (i.e., annexin V-137, -143, and -145). Results for individual organs fell into 3 groups. For the spleen, liver, and lung, 75%–85% of uptake was eliminated by eliminating the PS-binding activity of annexin V. For the stomach, small bowel, and heart, about 40%–50% of uptake was lost in the mutants compared with the wild-type protein. For the kidney and large bowel, uptake was unaffected by changes in affinity.
DISCUSSION
Characterization of 99mTc-Annexin V-128 and Comparison with 99mTc-HYNIC-Annexin V
We have recently developed a novel form of annexin V with an endogenous 99mTc chelation site, termed annexin V-128 (6), that has significant advantages over 99mTc-HYNIC-annexin V. There is a striking difference in the renal handling of 99mTc-labeled annexin V depending on the method of labeling. Initial renal uptake is the same, but subsequent urinary excretion of 99mTc is far more rapid for the 99mTc-annexin V-128 derivative compared with that of the 99mTc-HYNIC-annexin V derivative. The most likely explanation is as follows. Both proteins are filtered equally at the glomerulus, and both are taken up in the proximal tubules by relatively nonspecific uptake systems designed to capture low-molecular-weight proteins (23). Endocytosed proteins are then delivered to the lysosome and degraded by proteases to oligopeptides or amino acids. However, the end products differ greatly in their retention in the kidney. The end product of the degradation of 99mTc-HYNIC-annexin V is probably the lysine adduct of 99mTc-HYNIC complexed to tricine, which has a high degree of retention in lysosomes (24,25). The end product of the metabolism of 99mTc-annexin V-128 is not known but may consist either of a small oligopeptide or possibly pertechnetate alone. In any event, the radioactive end product is much more rapidly excreted from the kidney. Although we did not study metabolism of 99mTc-annexin V-128 in other organs, it seems unlikely that the rapid degradation seen in the kidney would also occur in apoptotic tissues because apoptotic bodies will have lost the multiple systems required for endocytosis, intracellular transport, and degradation of proteins bound to the cell surface. Thus, annexin V is likely to remain intact when bound to the surface of dead cells or their fragments.
The improvements in biodistribution achieved with annexin V-128 were obtained without any compromise in its ability to detect apoptosis in vivo, as shown by Figure 2. This protein also has significant advantages for manufacturing compared with HYNIC-annexin V: The end product of recombinant protein production is ready to be labeled directly with 99mTc and does not need to undergo further chemical derivatization, purification, and characterization. The single labeling site at the N-terminus is uniform and well characterized; this region of the protein is on the opposite side of the molecule from the membrane-binding region (5). In contrast, it can be difficult to obtain a reproducible preparation of chemically modified annexin V when amine-directed reagents are used; inevitably, the product is heterogeneous with respect to sites and stoichiometry of labeling. The Tc-labeling procedure is comparable in simplicity to that used with HYNIC-annexin: One simply mixes pertechnetate, tin reagent, and protein and incubates for 20–60 min at 22°C or 37°C (this study (12)). Overall, 99mTc-annexin V-128 is an attractive candidate to replace 99mTc-HYNIC-annexin V in many applications.
Factors Regulating Normal Organ Uptake of Annexin V
This study shows that uptake of annexin V in normal organs is differentially regulated. In some organs, such as liver, spleen, and lung, uptake is an inherent property of the protein and is primarily dependent on the PS-binding affinity of the molecule. In other organs, such as kidney and large bowel, uptake is relatively nonspecific and is not regulated by the PS-binding affinity of the molecule. Some organs, such as stomach, small bowel, and heart, are in-between, with about half their normal uptake being PS specific. Control experiments showed that these effects were not simply due to charge changes introduced by the point mutations used to reduce binding affinity. Similarly, the observed effects were not due to molecular weight changes, since the point mutations change the protein’s molecular mass by only 1–4 Da (6).
There are several possible explanations for the differential effects of PS-binding affinity on organ uptake. It is possible that the results reflect different levels of physiologic apoptosis in different organs. It is also possible that annexin V is binding to apoptotic cells or cell debris that is in the process of being engulfed by phagocytic cells in the liver, spleen, and lung. It is also possible that annexin V is binding to non-PS substances in some normal organs—for example, annexin V is known to bind to anionic polysaccharides such as heparan sulfate under some conditions (26). Finally, annexin V could be taken up by an organ-specific endocytic mechanism that is somehow dependent on its membrane-binding affinity. In any event, the results of this study show that the factors governing the biodistribution of annexin V are complex and are not simply a function of nonspecific factors, such as molecular size, charge, or vascular permeability.
Affinity Requirements for Annexin V Uptake in Apoptotic Tissues
Our results indicate clearly that all 4 domains of annexin V are needed for maximum uptake in apoptotic organs in vivo. Although previous work had suggested that most of the in vitro binding affinity of annexin V was concentrated in the first domain (27), our recent work with improved assay methods showed clearly that all 4 domains contribute equally to in vitro binding affinity (6). The results in this study now extend these observations to organ uptake in vivo. It is clear that organ uptake is a function of only the total number of active domains and is not affected by the specific domain(s) that have been inactivated by mutation. For example, single-site mutants in domains 1 and 4 behaved equivalently, as did double mutants in domains 1 + 4 versus domains 2 + 3; there was no difference between mutants containing different combinations of individual mutations, provided they contained the same total number of mutations.
This study also confirms the value of our recently developed methods for measuring the affinity of annexin V for membranes in vitro (20). These methods rely on quantitative calcium titration at low ratios of protein to membrane, which are probably most relevant to the in vivo situation. Although we have previously used this method with fluorescently labeled protein, the results of Figure 1 show that the method works equally well for cell-binding assays performed with radioactively labeled protein. We believe that these methods are the best overall means of measuring the membrane-binding affinity of modified annexins and will provide the best overall prediction of in vivo uptake. The logarithm of the binding constant (pK value) quantitatively predicts in vivo uptake: pK values 25% below wild-type are associated with significant loss of in vivo uptake, and pK values below 50% of wild-type are associated with complete loss of in vivo uptake (Table 2; Fig. 6A). It should be noted that the pK value is logarithmic; a decrease of 1 pK unit corresponds to a 10-fold reduction in the equilbrium constant.
CONCLUSION
There are several implications of this work for the further development of annexin V as an imaging agent for cell death in vivo. First, it appears that annexin V derivatives based on a single active domain (18), instead of the full 4 domains of the native protein, will probably lack the binding affinity required to provide effective target localization in vivo. Second, since the binding surface of annexin V encompasses contributions from all 4 domains (6,20), caution should be exercised in amine-directed labeling strategies that affect multiple residues on the same protein molecule, since there are multiple lysine residues present on or near the membrane-binding face of the molecule (5). However, modest changes in molecular charge per se, as can typically occur with modification of single lysine residues or the N-terminus, are unlikely to alter the clearance, biodistribution, or metabolism of the molecule. Finally, the use of the N-terminal endogenous 99mTc chelation sequence appears to have major advantages over the HYNIC chelator with regard to renal retention of 99mTc, with attendant decreased abdominal background and renal radiation dose.
Acknowledgments
We thank Donald Gibson for preparing experimental materials and Dr. Jean-Luc Vanderheyden of Theseus Imaging Corp. for the North American Scientific 2020 HYNIC-annexin V and tin-tricine kits. This research was supported by U.S. Public Health Service National Institutes of Health grants CA-102348 and EB-000898.
Footnotes
Received Oct. 18, 2004; revision accepted Jan. 16, 2005.
For correspondence or reprints contact: Jonathan F. Tait, MD, PhD, Department of Laboratory Medicine, University of Washington, Box 357110, Seattle, WA 98195-7110.
E-mail: tait{at}u.washington.edu
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Whole-Body Imaging of High-Dose Ionizing Irradiation-Induced Tissue Injuries Using 99mTc-Duramycin
- Apoptosis Imaging Probe Predicts Early Chemotherapy Response in Preclinical Models: A Comparative Study with 18F-FDG PET
- Recent Advances in the Molecular Imaging of Programmed Cell Death: Part I--Pathophysiology and Radiotracers
- Evaluation of Chemotherapy Response in VX2 Rabbit Lung Cancer with 18F-Labeled C2A Domain of Synaptotagmin I
- Time Course of Paclitaxel-Induced Apoptosis in an Experimental Model of Virus-Induced Breast Cancer
- The Role of Radiotracer Imaging in the Diagnosis and Management of Patients with Breast Cancer: Part 2--Response to Therapy, Other Indications, and Future Directions
- Imaging of Apoptosis
- 99mTc-Labeled Duramycin as a Novel Phosphatidylethanolamine-Binding Molecular Probe
- In Vivo Detection of Apoptosis
- Imaging Acute Cardiac Cell Death: Temporal and Spatial Distribution of 99mTc-Labeled C2A in the Area at Risk After Myocardial Ischemia and Reperfusion
- Molecular Position of Radiolabels and Its Impact on Functional Integrity of Proteins
- Improved Detection of Cell Death In Vivo with Annexin V Radiolabeled by Site-Specific Methods
- 99mTc-Labeled C2A Domain of Synaptotagmin I as a Target-Specific Molecular Probe for Noninvasive Imaging of Acute Myocardial Infarction
- Counting Heads in the War against Cancer: Defining the Role of Annexin A5 Imaging in Cancer Treatment and Surveillance
- Past, Present, and Future of Annexin A5: From Protein Discovery to Clinical Applications