


Abstract
Many diseases are associated with cytokine release after inflammatory infiltration, which perturbs organ function. Radioligands capable of noninvasive tracking to assess the integrity of specific biochemical pathways offer potent ways to establish such perturbing mechanisms. Methods: To demonstrate regulation of hepatobiliary transport in disease, we used 99mTc-mebrofenin in a carbon tetrachloride–induced liver injury model in Fischer 344 rats. Healthy rats served as control animals. Image analysis was used to determine 99mTc-mebrofenin handling. Liver tests and histologic analysis were used for grading liver injury and hepatic fibrosis. To address the role of inflammatory cytokines, we used in vitro assays with 99mTc-mebrofenin-loaded primary rat hepatocytes. Results: In healthy rats, 99mTc-mebrofenin was promptly excreted, and after 1 h only 20% ± 5% (mean ± SD) of peak 99mTc-mebrofenin activity remained in the liver. In contrast, rats treated with carbon tetrachloride for 1 or 3 mo showed 84% ± 5% and 80% ± 7% (mean ± SD), respectively, of peak 99mTc-mebrofenin activity in the liver after 1 h (P < 0.001). Abnormal 99mTc-mebrofenin transport was associated with necroinflammatory activity and not hepatic fibrosis. This was examined directly in animals, where withdrawal of carbon tetrachloride for 2 wk after significant liver injury produced loss of inflammatory activity without affecting hepatic fibrosis. In this situation, 99mTc-mebrofenin transport returned to normal, indicating a central role of inflammatory activity in this process. In vitro assays showed impairment in 99mTc-mebrofenin excretion after incubation of cultured hepatocytes with interleukin-6 and further impairment with interleukin-6 plus tumor necrosis factor-α. Conclusion: The findings indicate that inflammatory cytokines regulate 99mTc-mebrofenin transport. This cytokine-mediated process establishes a paradigm for identifying and monitoring organ inflammation, including in viral or alcoholic hepatitis, fatty liver disease, allograft rejection, and responses to gene therapy vectors.
The integrity of discrete biochemical and metabolic pathways in cells is disturbed during disease. Although various pathophysiologic alterations are often helpful in establishing the presence or extent of organ damage, specific metabolic radioligands are of major interest because they permit noninvasive assessment of molecular pathways (1,2). For instance, 99mTc-mebrofenin (99mTc-N-[3-bromo-2,4,6-trimethyacetanilide]iminodiacetic acid) and related compounds, widely used for assessing gallbladder and liver function, are incorporated exclusively in the liver followed by transport into bile (3–11). It is generally accepted that, because hepatic incorporation and biliary transport of 99mTc-mebrofenin depend on the prevailing mass of hepatocytes, 99mTc-mebrofenin handling is altered by the presence of liver disease. However, the mechanism of 99mTc-mebrofenin retention in chronic liver disease in the absence of gallbladder dysfunction or biliary obstruction is unclear. Because 99mTc-mebrofenin excretion in bile requires specific transporters (12), one possibility is that hepatic inflammation, cell damage, or fibrosis produces abnormal regulation of 99mTc-mebrofenin transporter activity. The roles of such mechanisms in the regulation of biliary transport activity offer potential for advancing insights into radiotracer biology.
We postulated that specific mechanisms could be established in animals, where progressive depletion of the parenchymal liver cell mass is associated with inflammatory and profibrogenic activity, as observed during repeated administration of carbon tetrachloride (CCl4) to rats (13). CCl4 induces peroxidative liver injury after conversion to toxic intermediates by cytochrome P450 enzymes. Whereas discrete exposure to CCl4 results in acute liver injury, continuous exposure to this toxin produces progressive liver injury and fibrosis, eventually causing cirrhosis, portal hypertension, and death (13,14). A noteworthy feature of CCl4-induced hepatotoxicity concerns reversal of liver disease after its withdrawal in rats. In that situation, necroinflammatory activity subsides rapidly, followed by regression of fibrosis, and liver histology gradually returns to normal (13). Here, we report studies in CCl4-treated rats to determine mechanisms regulating handling of 99mTc-mebrofenin transport in the liver. Our studies were aimed at establishing the role of hepatic necroinflammatory activity versus fibrosis and associated mechanisms in perturbing 99mTc-mebrofenin handling and analysis of disease-specific pathophysiologic events—for example, the effect of specific inflammatory cytokines.
MATERIALS AND METHODS
Animals
Thirty young adult Fischer 344 (F344) rats, 6–8 wk old and weighing 120–160 g, were used. Rats were divided into 5 groups of 6 animals each: 1 control group and 4 experimental groups. Rats in experimental groups were treated with CCl4 under phenobarbitone priming, according to previously established protocols (13). Briefly, 0.5 mL/kg CCl4 in mineral oil (1:1, v/v) was administered intramuscularly at 4- to 5-d intervals, along with 0.5 g phenobarbitone per liter of drinking water (Sigma Chemical Co.). Three groups of rats received CCl4 and phenobarbitone for 1, 2, or 3 mo. One group of rats received CCl4 and phenobarbitone for 3 mo followed by withdrawal of both for 2 wk. All animals were sacrificed after completing the studies.
Surgical Procedures
Rats were anesthetized with ketamine and xylazine. The spleen was isolated via laparotomy, and a 23-gauge butterfly needle was inserted into the splenic pulp to inject 7.4 MBq 99mTc-mebrofenin in 1 mL. Hemostasis was secured with a ligature around the splenic pole (9). Liver samples were obtained for histology after completion of imaging.
Nuclear Medicine Procedures
Mebrofenin was mixed with 185–222 MBq 99mTc-sodium pertechnetate in 3 mL normal saline according to the manufacturer’s instructions (Choletec; Bracco Diagnostics). A γ-camera (Argus; ADAC Laboratories), equipped with a low-energy, high-resolution, parallel-hole collimator and interfaced to a computer, was used for image acquisition. A 20% window centered on 140 keV was used for energy discrimination. Ten-second dorsal images were acquired for 60 min, using a 64 × 64 × 16 matrix at a zoom factor of 2. Data were analyzed using a conventional nuclear medicine workstation (Pegasys; ADAC Laboratories). Regions of interest were drawn over the liver, and hepatic time–activity curves were generated. The time at which maximal hepatic activity of 99mTc-mebrofenin occurred (Tpeak) as well as the time required for peak 99mTc-mebrofenin activity to decrease by 50% (T1/2 peak) were determined. The percent hepatic retention of the peak activity at 10, 20, 30, and 60 min after 99mTc-mebrofenin administration was also determined. Residual 99mTc-mebrofenin activity in excised livers was measured with a γ-counter.
Histologic Analysis
Liver samples were fixed in 10% buffered formalin. Paraffin-embedded sections were prepared and stained with hematoxylin–eosin or Gomori’s 1-step trichrome staining using standard procedures. Rats with chronic liver disease show hepatocellular necrosis or apoptosis, steatosis, hydropic change, megalocytosis, mitosis, and fibrosis. These changes were graded according to the criteria of Batts and Ludwig (15). Hepatic disease activity was graded by analysis of portal or lobular inflammation and necrosis: grade 0, no disease activity; grade 1, minimal portal inflammation with minimal or patchy lobular inflammation and necrosis; grade 2, mild portal inflammation with mild lobular inflammation and little hepatocellular change; grade 3, moderate portal inflammation, moderate lobular inflammation and necrosis with noticeable hepatocellular damage; and grade 4, severe portal and lobular inflammation, necrosis with prominent diffuse hepatocellular damage. Hepatic fibrosis was graded as follows: grade 0, no fibrosis; grade 1, portal fibrosis; grade 2, periportal fibrosis; grade 3, septal fibrosis; and grade 4, cirrhosis. The cumulative histologic scores could have ranged from 0 (entirely normal) to 8 (severe disease with cirrhosis).
Laboratory Tests
Sera stored at −20°C were analyzed for bilirubin, albumin, and alanine aminotransferase (ALT) with an automated microsystem (Bayer-Chem-1; Bayer Corp.).
Cell Culture Assays
Primary hepatocytes were isolated from F344 rats by 2-step collagenase perfusion as described previously (13). Cell viability was estimated by trypan blue dye exclusion. Cell culture dishes were coated with type-1 collagen isolated from the tail of F344 rats, as described previously (16). Cells were cultured in dishes with 5–6 × 104 cells per 24-well dish (Nalge Nunc International) in RPMI 1640 medium (Life Technologies) containing 100 U/mL penicillin, 100 μg/mL streptomycin, and 10% fetal bovine serum (Atlanta Biologicals). To analyze 99mTc-mebrofenin incorporation and release, 370 kBq 99mTc-mebrofenin were added to each well containing cells after culture for >40 h. Cells were incubated with 99mTc-mebrofenin for 30 min at 37°C and washed 3 times with cold phosphate-buffered saline, pH 7.4. Medium was collected from 6 replicate wells, each after 30, 60, 90, and 120 min, and cells were lysed in 10 mmol/L ethylenediaminetetraacetic acid and 0.5% sodium dodecyl sulfate for measuring 99mTc-mebrofenin activity. To determine nonspecific 99mTc-mebrofenin binding, cells were pretreated with 5 mmol/L of the nonhydroxamate iron chelator diethylenetriaminepentaacetic acid (DTPA) or potassium cyanide. The cellular protein content was measured by the Bradford assay (Bio-Rad).
Recombinant human tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) (Intergen Co.) were added to cells after 20–24 h of culture to perturb 99mTc-mebrofenin handling. In initial studies, cells were primed with 100 or 200 ng/mL actinomycin D (Sigma) for 30 min before adding 10–20 ng/mL cytokines, according to Jones et al. (16). To demonstrate perturbations in cell viability, utilization of the thiazolyl blue dye (3-[4,5-dimethylthiozol-2-yl]-2,5-diphenyl tetrazolium bromide [MTT]; Sigma) was analyzed, as described previously (16). In aliquots, total cellular protein was measured by the Bradford assay. Final cell culture studies used 10 ng/mL TNF-α and IL-6. To demonstrate that cytokines were bioactive, an initial experiment was performed, in which hepatocytes were incubated with TNF-α with or without actinomycin D priming. All studies used 6 replicate conditions and were performed with cells from separate normal F344 rats on 3 occasions.
Statistical Analysis
Data are presented as mean ± SD and were analyzed with statistical software (SigmaStat 2.0; Jandel Scientific). To analyze the significance of differences, the Student t test was used for intergroup comparisons and ANOVA was used for multigroup comparisons. Multiple linear regression was used to establish dependencies among discrete variables. P < 0.05 was considered significant.
RESULTS
Laboratory Tests
Serum bilirubin levels were normal in all animals studied. Serum albumin levels decreased with increasing duration of CCl4 treatment, such that albumin levels were significantly lower in CCl4 recipients treated for 2 and 3 mo than in normal untreated rats. Serum ALT levels were significantly elevated in CCl4-treated rats (Table 1).
Liver Test Abnormalities in Rats Studied
Histopathologic Changes
Animals treated with CCl4 for 1 mo showed macro- and microvesicular hepatic steatosis, hepatocellular necrosis, nuclear abnormalities, inflammatory cell infiltrates, and portal fibrosis (Fig. 1). Disease activity was most pronounced in perivenous areas of the liver lobule, as expected, since CCl4 is preferentially metabolized in perivenous hepatocytes with greater cytochrome P450 enzyme activity. The magnitude of liver injury increased after additional CCl4. Hepatic fibrosis was most pronounced in animals treated for 3 mo, with fibrotic septae extending from portal-to-portal or portal-to-perivenous areas (bridging fibrosis), along with fibrotic nodules. Extensive hepatic reorganization found in established cirrhosis and ascites or extensive nodules were not observed. Liver abnormalities were absent in normal untreated control rats. Histologic grading indicated significant disease activity in animals treated with CCl4 (Table 2). Similarly, the grade of fibrosis advanced with greater cumulative doses of CCl4. Analysis of histologic changes demonstrated that disease activity and fibrosis increased in animals between 1 and 3 mo after CCl4 (P = 0.005, ANOVA; Dunnett pairwise comparisons 1, 2, and 3 mo after CCl4 vs. controls, P < 0.05) (Table 2).
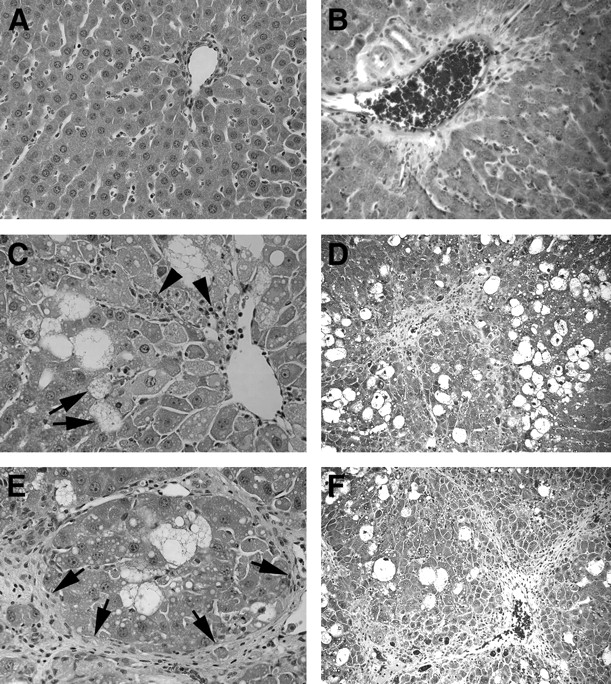
Liver injury in animals treated with CCl4. (A and B) Untreated control rats with normal liver histology. (C and D) Rat treated for 1 mo with CCl4. Note extensive fatty change (C, arrows), cell necrosis, and inflammatory activity (C, arrowheads), along with fibrosis. (E and F) Rat treated for 3 mo with CCl4 shows inflammation, cell necrosis, and extensive fibrosis (E, arrows). (A, C, and E: hematoxylin–eosin stain, ×200; B, D, and F: trichrome stain for fibrosis, ×100)
Histologic Grading of Liver Lesions
99mTc-Mebrofenin Handling
In normal control rats, 99mTc-mebrofenin appeared promptly in the liver and bile, with rapid clearance from the liver (Fig. 2). Hepatic 99mTc-mebrofenin uptake was not affected by CCl4 treatment, regardless of the treatment duration (Tpeak: 140 ± 7 s, normal controls vs. 165 ± 70 s, CCl4 for 3 mo, P = not significant [NS]). 99mTc-Mebrofenin clearance, in contrast, was markedly delayed in CCl4-treated rats, although the overall pattern of 99mTc-mebrofenin retention in the liver was not different in animals treated with CCl4 for various times. In normal untreated control rats, the T1/2 of 99mTc-mebrofenin excretion was 356 ± 86 s (Fig. 3). In contrast, 99mTc-mebrofenin excretion was severely impaired in animals treated with CCl4. Because of extremely slow 99mTc-mebrofenin excretion in these animals, as indicated in Figure 3, the slopes were not amenable for estimating T1/2 values of 99mTc-mebrofenin excretion.

Scintigraphy shows hepatic 99mTc-mebrofenin uptake and excretion. Sequential images are shown from same animals. (A) Normal rat shows rapid 99mTc-mebrofenin clearance. In contrast, rats treated with CCl4 for 1 mo (B) or 3 mo (C) show markedly delayed 99mTc-mebrofenin clearance.

Analysis of 99mTc-mebrofenin retention in liver of normal untreated rats and rats treated for 1 or 3 mo with CCl4. Note that largest fraction of 99mTc-mebrofenin was cleared from liver of normal rats within 10 min, whereas 99mTc-mebrofenin was retained in animals after liver injury.
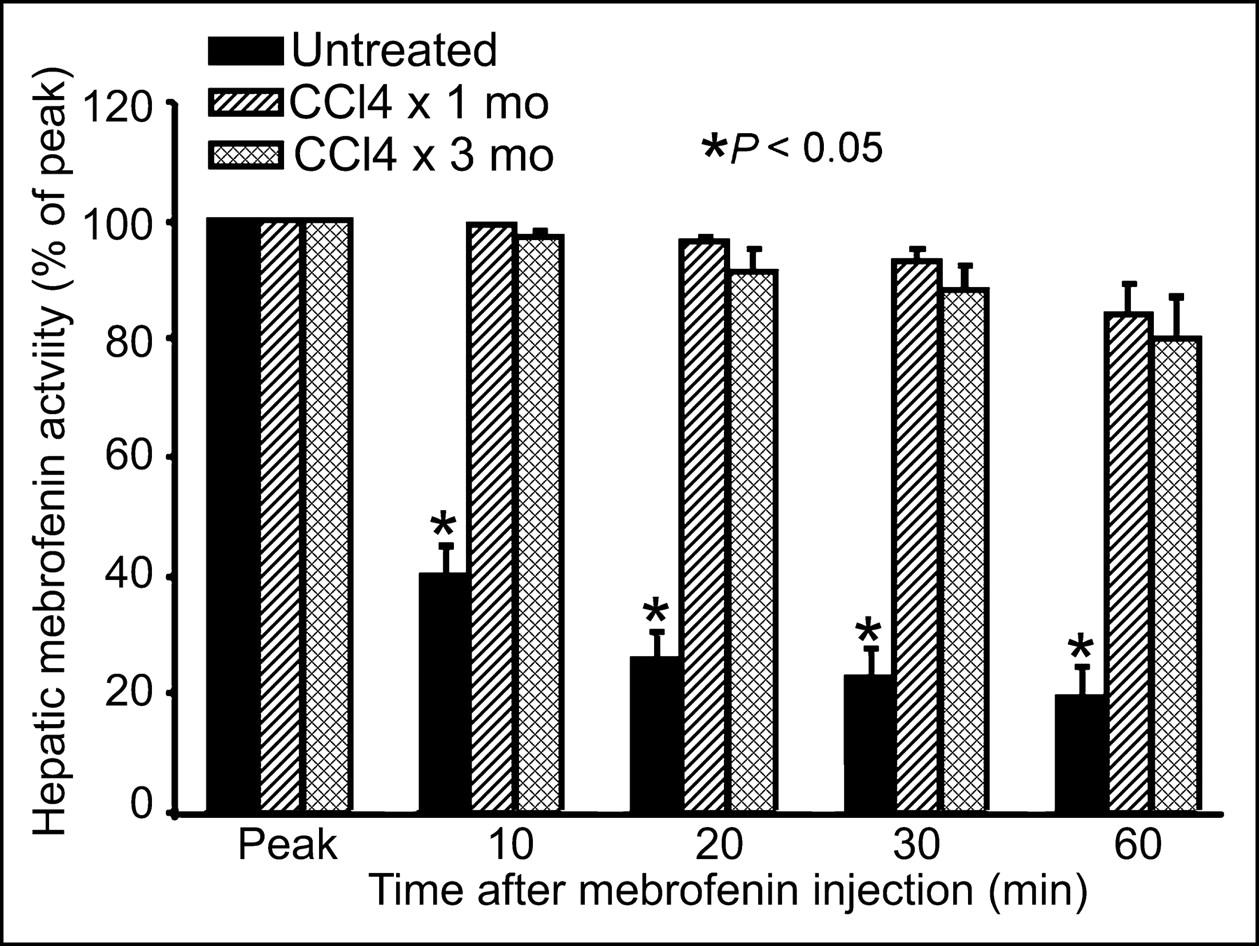
In normal control rats, at 20 and 60 min after intrasplenic injection, only 26% ± 3% and 20% ± 5%, respectively, of the peak 99mTc-mebrofenin activity remained in the liver. In contrast, 96% ± 1% and 91% ± 4% of the 99mTc-mebrofenin activity was still present in the liver after 20 min in animals treated with CCl4 for 1 and 3 mo, respectively (P < 0.001 vs. controls, t tests). At 60 min, animals treated with CCl4 for 1 and 3 mo still retained much of the 99mTc-mebrofenin in the liver, 84% ± 5% and 80% ± 7%, respectively (P < 0.001, t tests). Similarly, measurement of the retained 99mTc-mebrofenin activity in excised livers obtained after imaging revealed that rats treated with CCl4 for 1 or 3 mo contained on average 7.3-fold and 5.2-fold greater activity than that of untreated normal rats (P = 0.002, Mann–Whitney rank sum tests) (1.29 ± 1.09 × 105 counts per minute [cpm]/g liver, controls, vs. 9.43 ± 1.42 × 105 cpm/g liver and 6.71 ± 2.19 × 105 cpm/g liver, 1 and 3 mo after CCl4, respectively).
Multiple linear regression established a significant relationship between hepatic disease activity and 99mTc-mebrofenin retention (Table 3) when serum ALT, disease activity, and liver fibrosis were assigned as independent variables and 99mTc-mebrofenin retention was assigned as a dependent variable. These findings indicated that hepatic disease activity played a significant role in 99mTc-mebrofenin clearance. Study of rats administered CCl4 for 3 mo, followed by CCl4 withdrawal for 2 wk, showed extensive fibrosis but absence of liver inflammation (Fig. 4). In agreement with these findings, liver tests showed significant improvement, with nearly normal serum ALT (122 ± 116 IU/L), improved serum albumin (2.8 ± 0.1 g/dL), and normal serum bilirubin (0.3 ± 0.15 mg/dL) levels. The disease activity grade decreased to 1 ± 0, although the fibrosis grade remained elevated at 2.4 ± 0.5, which was virtually similar to that after 3 mo of CCl4. Hepatic 99mTc-mebrofenin clearance in these animals was normal (Fig. 4). The T1/2 of 99mTc-mebrofenin excretion was 322 ± 33 s, similar to that of normal untreated control rats (P = NS), and residual 99mTc-mebrofenin activity 20 and 60 min after peak 99mTc-mebrofenin accumulation was 25% ± 5% and 20% ± 6%, respectively, which was similar to that of normal untreated control rats (P = NS).
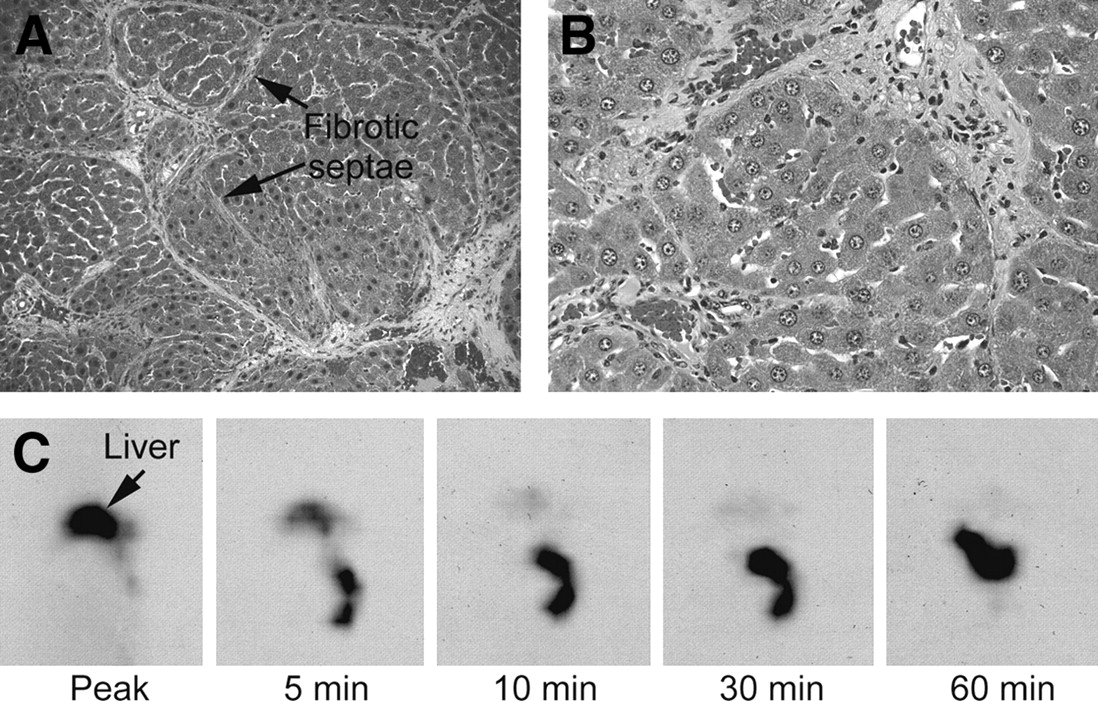
Normalization of 99mTc-mebrofenin excretion in animals after CCl4 withdrawal. (A and B) Rats show persistent fibrosis (A, arrows; trichrome stain, ×40) but resolution of inflammatory activity (B, hematoxylin–eosin stain, ×100). (C) Scintigraphy shows rapid 99mTc-mebrofenin clearance in a rat (compare with Fig. 2A).
Associations Between Clinical Parameters and 99mTc-Mebrofenin Excretion
Role of Cytokines in Hepatic 99mTc-Mebrofenin Excretion in Primary Rat Hepatocytes
Incubation of cells with TNF-α alone had no effect on MTT dye utilization. However, after culture with TNF-α plus actinomycin D, cell viability declined to 45% ± 4% of that of untreated controls or controls treated with TNF-α alone (P < 0.001, t test), which was an appropriate and expected response (17). In further studies, loading of cells with 99mTc-mebrofenin was successful and provided a suitable assay to analyze the kinetics of 99mTc-mebrofenin excretion in vitro.
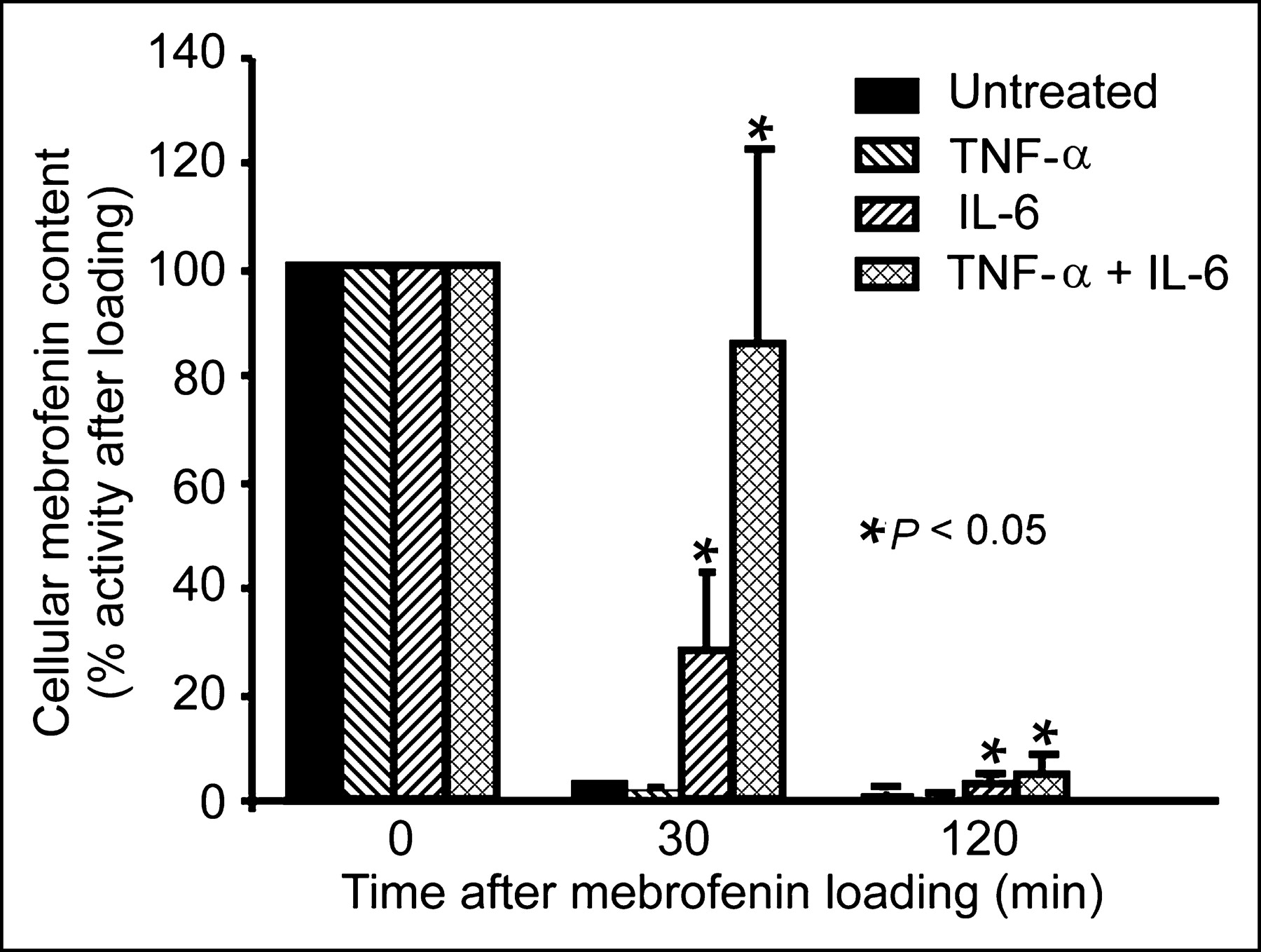
In untreated control hepatocytes, 96.8% ± 1.0% of the accumulated 99mTc-mebrofenin activity was cleared after 30 min and only 1.2% ± 0.5% of 99mTc-mebrofenin activity was left in cells after 3 h (Fig. 5). Similarly, cells treated with TNF-α alone promptly excreted 99mTc-mebrofenin. However, treatment of cells with IL-6, and especially with the combination of IL-6 and TNF-α, induced significant 99mTc-mebrofenin retention (P < 0.001, t tests). In this situation, 30 min after completion of cell loading, IL-6 was associated with retention of 28% ± 15% of 99mTc-mebrofenin activity, whereas IL-6 plus TNF-α resulted in retention of 85% ± 36% of 99mTc-mebrofenin activity (P < 0.001, t test). Essentially no 99mTc-mebrofenin was incorporated in cells incubated with either DTPA or potassium cyanide, indicating the absence of nonspecific 99mTc-mebrofenin adherence to cells under the assay conditions.

Cytokine-induced perturbation of 99mTc-mebrofenin handling in cultured hepatocytes. Data are from a typical experiment with prompt clearance of 99mTc-mebrofenin in cultured hepatocytes after 99mTc-mebrofenin loading for 30 min. Exposure of cells to TNF-α did not affect 99mTc-mebrofenin clearance, whereas exposure to either IL-6 alone or IL-6 plus TNF-α resulted in significant 99mTc-mebrofenin retention in cells.
DISCUSSION
Our findings demonstrate that abnormal retention of 99mTc-mebrofenin in the liver requires the presence of inflammation and is independent of chronic hepatocellular damage and liver fibrosis. The data further establish that the process affecting 99mTc-mebrofenin transport in inflammation is mediated by cytokines. This mechanism in the regulation of 99mTc-mebrofenin handling will enable further rational applications of 99mTc-mebrofenin imaging in health and disease.
We found that 99mTc-mebrofenin was rapidly extracted by the liver in both normal and diseased animals, as shown by the Tpeak values, which was similar to that of LEC rats with significant copper toxicosis-related chronic liver disease (9). In agreement with the 2-compartment modeling, taking into account fluxes of 99mTc-mebrofenin between systemic and hepatic blood pools and hepatocyte uptake (3,4), we previously showed that the hepatic extraction efficiency for 99mTc-mebrofenin follows first-order kinetics and is extraordinarily high. Also, our previous findings established that depletion of the reference systemic (cardiac) blood pool of 99mTc-mebrofenin at early times, before the commencement of biliary excretion in normal rats and LEC rats with liver disease, was similar, indicating that hepatic 99mTc-mebrofenin extraction was equally efficient in the presence of chronic liver disease (9). Our present findings provide further support that hepatic extraction of 99mTc-mebrofenin is unperturbed in chronic liver disease. The fractional excretion of 99mTc-mebrofenin in normal control rats, as judged by the T1/2 measurement of 99mTc-mebrofenin excretion, as well as 99mTc-mebrofenin retention at various times for up to 60 min, was also similar to that of previous studies (9) and was in general agreement with observations in healthy people (18). However, 99mTc-mebrofenin excretion was impaired markedly in animals with CCl4-induced liver disease. Of note was the extensive perturbation of 99mTc-mebrofenin excretion in animals at early stages of liver inflammation. 99mTc-Mebrofenin retention was obvious within 20 min and remained markedly elevated after 60 min, indicating profound perturbations in the regulation of 99mTc-mebrofenin transport in bile.
An examination of the molecular regulation of hepatic bile acid and organic anion transporters by cytokines should be appropriate for explaining the relationship between inflammatory activity and 99mTc-mebrofenin excretion. Although 99mTc-mebrofenin is a bifunctional chelating agent, with one end conjugated to 99mTc and the other to the lidocaine analog acetanilide, the identity of the specific transporter(s) involved in the hepatic excretion of 99mTc-mebrofenin is unknown. On the other hand, the list of bile salt and organic anion transporters has grown extensively and significant complexities have emerged (12). Among the hepatic uptake mechanisms, though the Na+/taurocholate cotransporting polypeptide (ntcp) is a major player in bile salt incorporation, a variety of sodium-independent transporters have also been found, including for bilirubin, organic anions, and multiple drugs—for example, organic anion transporters OAT-2, OATP-A, OATP-B, OATP-C, and OATP8. Because 99mTc-mebrofenin incorporation was unchanged in our rats with liver injury, these transporters are not likely incriminated, and adenosine triphosphate–dependent proteins involved in bile canalicular transport and basolateral export of organic anions may be more relevant. The bile canalicular transporters include the bile salt excretory pump (BSEP), which probably plays small role in organic anion transport, and the multidrug-resistance–related protein MRP2, as well as basolateral exporters, including MRP1 and MRP3, which may play more significant roles in organic anion transport.
Recent studies in rats treated with CCl4 showed that TNF-α downregulated expression of ntcp, OATP1, and OATP2 at the messenger RNA (mRNA) level (19). In mice, TNF-α affected mRNA levels of MRP2, MRP3, and OATP2 but not OATP1 or BSEP, whereas IL-6 decreased MRP2, OATP1, OATP2, and BSEP mRNA expression (20). Our results, which showed that TNF-α alone did not impair 99mTc-mebrofenin excretion, whereas IL-6 inhibited 99mTc-mebrofenin excretion, both directly and synergistically with TNF-α, suggest that the list of cytokine-regulated transporters responsible for these perturbations could be pared down. Thus, further analysis will be appropriate to determine what types of inflammatory activity will perturb hepatic 99mTc-mebrofenin handling in vivo. Similarly, manipulation of liver gene regulation by cytokines offers potent ways to identify the 99mTc-mebrofenin transporters. Although cultured hepatocytes have previously been shown to incorporate 99mTc-mebrofenin and related compounds (21), our in vitro assays should help demonstrate further mechanisms regulating 99mTc-mebrofenin excretion. For instance, nuclear hormone receptors regulate transcription of bile salt and organic anion transporters, including the farnesoid X receptor, which regulates BSEP transcription, and the hepatocyte nuclear factor (HNF)-1, HNF-4, retinoid X receptor/retinoic acid receptor, and other transcription factors, which are downregulated during cytokine-mediated acute liver inflammation, regulate expression of organic anion transporters, including MRP-2 (19).
Our findings indicate that fibrosis alone does not alter hepatic 99mTc-mebrofenin handling. Similarly, hepatic necrosis alone was unlikely to be responsible for altering 99mTc-mebrofenin handling, as suggested by the lack of an association between serum ALT levels and 99mTc-mebrofenin retention. On the other hand, numerous conditions are associated with cytokine release, ranging from inflammatory states—for example, acute cholecystitis, viral or alcoholic hepatitis, fatty liver disease, and so forth. It should be appropriate to undertake studies of 99mTc-mebrofenin handling in clinical situations, where hepatic inflammation is driven by specific populations of inflammatory cells—for example, neutrophilic infiltrates in alcoholic hepatitis, lymphocytic infiltrates in viral hepatitis, and plasma cell accumulation in autoimmune hepatitis.
CONCLUSION
Use of 99mTc-mebrofenin imaging in the context of cytokine activity will be effective for qualitative and quantitative analysis of liver inflammation. This will be potentially valuable for noninvasively assessing therapeutic responses in inflammatory liver diseases, which often necessitates repeated liver biopsies.
Acknowledgments
This research was supported in part by National Institutes of Health grants R01 DK46952 and P30 DK41296.
Footnotes
Received Jul. 16, 2004; revision accepted Aug. 30, 2004.
For correspondence or reprints contact: Kuldeep K. Bhargava, PhD, Division of Nuclear Medicine, Long Island Jewish Medical Center, 270-05, 76th Ave., New Hyde Park, NY 11042.
E-mail: bhargava{at}lij.edu
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Repurposing 99mTc-Mebrofenin as a Probe for Molecular Imaging of Hepatocyte Transporters
- PET with 64Cu-Histidine for Noninvasive Diagnosis of Biliary Copper Excretion in Long-Evans Cinnamon Rat Model of Wilson Disease
- Spontaneous origin from human embryonic stem cells of liver cells displaying conjoint meso-endodermal phenotype with hepatic functions
- Comparison Between the Values of the Hepatic Uptake Rate Obtained by 2 Methods, Using Hepatobiliary Scintigraphy in Patients with Nonalcoholic Steatohepatitis