


Abstract
Disseminated, metastatic cancer is frequently incurable. Targeted α-particle emitters hold great promise as therapeutic agents for disseminated disease. 225Ac is a radionuclide generator that has a 10-d half-life and results in α-emitting daughter elements (221Fr, 217At, 213Bi) that lead to the emission of a total of 4 α-particles. The aim of this study was to develop approaches for stable and controlled targeting of 225Ac to sites of disseminated tumor metastases. Liposomes with encapsulated 225Ac were developed to retain the potentially toxic daughters at the tumor site. Methods: 225Ac was passively entrapped in liposomes. To experimentally test the retention of actinium and its daughters by the liposomes, the γ-emissions of 213Bi were measured in liposome fractions, which were separated from the parent liposome population and the free radionuclides, at different times. Under equilibrium conditions the decay rate of 213Bi was used to determine the concentration of 225Ac. Measurements of the kinetics of 213Bi activity were performed to estimate the entrapment of 213Bi, the last α-emitting daughter in the decay chain. Results: Stable pegylated phosphatidylcholine-cholesterol liposomes of different sizes and charge were prepared. Multiple (more than 2) 225Ac atoms were successfully entrapped per liposome. 225Ac retention by zwitterionic liposomes was more than 88% over 30 d. Retention by cationic liposomes was lower. A theoretical calculation showed that for satisfactory 213Bi retention (>50%), liposomes of relatively large sizes (>650 nm in diameter) are required. 213Bi retention was experimentally verified to be liposome-size dependent. For large liposomes, the measured 213Bi retention was lower than theoretically predicted (less than 10%). Conclusion: This work supports the hypothesis that it may be possible to develop 225Ac-based therapies by delivering multiple 225Ac atoms in liposomes. Improvements in the retention of 225Ac daughters will likely be necessary to fulfill this potential. Because of the size of the liposomal structures required to contain the daughters, the approach is ideally suited for locoregional therapy (e.g., intraperitoneal, intrahepatic artery, or intrathecal).
Disseminated micrometastatic disease is rarely cured by current treatment options. Targeted α-particle emitters hold great promise as therapeutic agents for micrometastases. α-Particles are highly potent cytotoxic agents, potentially capable of tumor-cell kill without limiting morbidity. The increased effectiveness of α-particles is due to the amount of energy deposited per unit distance traveled (linear energy transfer, or LET). For α-particles, this LET is approximately 400 times greater than that of β-particles (80 keV/μm vs. 0.2 keV/μm). Cell survival studies have shown that α-particle–induced killing is independent of oxygenation state or cell-cycle during irradiation and that as few as 1–3 tracks across the nucleus may result in cell death (1–3). In addition, the 50- to 100-μm range of α-particles is consistent with the dimensions of micrometastatic disseminated disease, allowing for localized irradiation of target cells with minimal normal-cell irradiation.
Human use of antibody-targeted α-particle emitters has been reported in 2 trials (4–6). The first injection of an α-particle emitter to humans for radioimmunotherapy was of 213Bi conjugated to the anti-CD33 antibody HuM195, targeting myeloid leukemia (5–7). This trial demonstrated feasibility and anticancer activity with minimal toxicity. In the second of the 2 human trials, the antitenascin antibody 81C6, labeled with the α-particle emitter 211At, was injected into surgically created cavities in patients with malignant gliomas. This trial has demonstrated substantially better tumor control relative to 131I-labeled 81C6 antibody (4). In the setting of minimal disease, animal studies have shown that α-particle emitters yield superior tumor control relative to β-emitters (8–13).
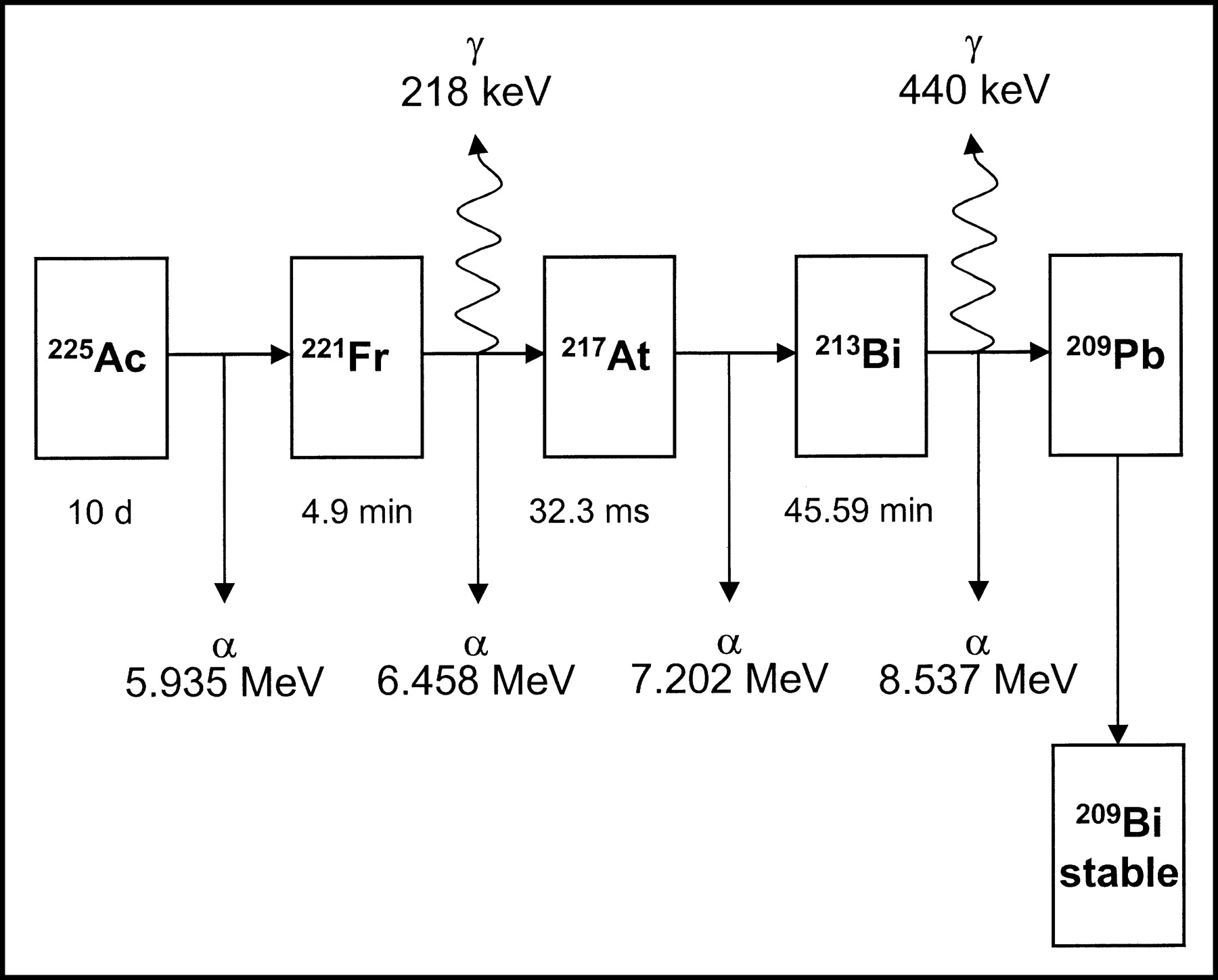
In this study, the α-particle–emitting element generator 225Ac was proposed for potential use in treatment of micrometastatic disease. 225Ac has a 10-d half-life, and its decay, and those of its daughters, lead to the emission of 4 α-particles (Fig. 1).

Simplified 225Ac decay cascade with associated particulate decays and half-lives.
To deliver an increased number of 225Ac atoms at the target site, liposomes with encapsulated multiple 225Ac atoms were proposed. Liposomes are closed structures defined by a phospholipid bilayer membrane that encloses an aqueous compartment (14). α-Particles traverse the phospholipid membrane without energy dissipation. Liposomes have been studied for more than 30 y, in particular as vehicles for drug delivery (15,16). Their application in drug delivery has been made possible by the development of sterically stabilized structures that use polyethylene glycol (PEG) chains to reduce uptake and catabolism of intravenously administered liposomes by the reticuloendothelial system, thereby increasing circulatory half-life. Such liposomes are typically 100–150 nm in diameter since this size range reduces reticuloendothelial system uptake while retaining adequate aqueous volume for drug delivery. Liposome tumor localization is dependent on the differential permeability of normal- versus tumor-tissue capillaries (17–19). Liposomes have also been used to deliver radionuclides, primarily for tumor diagnosis and infectious site imaging (20,21).
Studies in animals have shown that 225Ac is substantially more potent and more toxic than 213Bi (12,22,23). The toxicity arises because conjugation to antibodies or other molecular vehicles for targeting of this radionuclide can stably deliver only the first of the 4 α-particle–emitting atoms for several reasons: First, the bond between the targeting vehicle and the chelate holding the radionuclide may be broken on transformation of the parent to new daughter and emission of the first α-particle; second, the new daughter has chemistry different from that of the parent; and third, there is recoil of the parent atom away from the chelate. Subsequent α-particle–emitting intermediates (daughters) are, therefore, free to distribute throughout the body and to irradiate healthy organs and tissues.
Liposomes with encapsulated 225Ac should also retain the α-emitting intermediates at the tumor site. Because of their position in the periodic table, the daughters of 225Ac are likely to be ionic (Fr+1, At−1, Bi+3). The hydrophobic region of liposomal membranes provides a barrier to loss by diffusion of ionic species entrapped in the internal aqueous compartment (24). Therefore, if 225Ac is initially entrapped within the aqueous compartment of liposomes, the daughters are expected to be retained within the liposomes. Some loss is unavoidable, however, because the recoil distance traveled by daughter atoms, on decay of the parent, is on the order of the liposome size.
In this study, the feasibility of engineering liposomal vesicles capable of retaining 225Ac and its daughters for cancer therapy was investigated.
MATERIALS AND METHODS
Reagents
The lipids l-α-phosphatidylcholine (egg), 1,2-dipalmitoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] (ammonium salt) (PEG-labeled lipid), 1,2-dipalmitoyl-sn-glycero-3-ethylphosphocholine (chloride salt), l-α-phosphatidylethanolamine-N-(4-nitrobenzo-2-oxa-1,3-diazole) (egg) (NBD-PE), and l-α-phosphatidylethanolamine-N-(lissamine rhodamine B sulfonyl) (egg) (purity > 99%) were purchased from Avanti Polar Lipids. Cholesterol, ascorbic acid, phosphate-buffered saline (PBS), fluorexon (calcein), and Sephadex G-50 were purchased from Sigma. 1,4,7,10-Tetraazacyclododecane-1,4,7,10-tetraacetic acid (DOTA) was obtained from Macrocyclics. Dithionite (sodium hydrosulfite, technical grade, approximately 85%) was obtained from Acros Organics. 225Ac was obtained from Oak Ridge National Laboratory.
Liposome Preparation
Mixtures of phosphatidylcholine, cholesterol (1:1 molar ratio) and PEG-labeled lipids (6-mol percentage of total lipid) in CHCl3 were dried in a rotary evaporator. For stability measurements, the lipids were resuspended in calcein solution (55 mmol/L calcein in phosphate buffer, isosmolar to PBS, pH = 7.4). For 225Ac passive entrapment, the lipids were resuspended in PBS containing chelated actinium complexes (225Ac-DOTA, 1.85–3.70 MBq) (12) and diethylenetriaminepentaacetic acid. The lipid suspension was then annealed to 55°C for 2 h (25). To make liposomes, the lipid suspension was then taken through 21 cycles of extrusion (LiposoFast; Avestin) through 2 stacked polycarbonate filters (100-, 400-, and 800-nm filter pore diameter). Unentrapped contents were removed by size exclusion chromatography (SEC) in a Sephadex G-50 packed 1 × 10 cm column, eluted with an isotonic buffer. Ascorbic acid (8 mmol/L) was coentrapped to minimize lipid oxidation due to radiation (26).
To make unilamellar liposomes for radionuclide binding measurements, the above procedure was repeated. The lipids were resuspended in sucrose solution (138 mmol/L, pH = 7.4) isosmotic to PBS and extruded through filters with a 100-nm pore diameter. Unentrapped sucrose was removed by dilution of the liposome suspension in PBS, followed by ultracentrifugation (2 h, 142,000g, 25°C). The pellet (10% v/v) was resuspended in PBS and used for the 225Ac binding measurements. The unilamellar character of the zwitterionic and cationic liposomes was tested using dithionite. Dithionite ion S2O4− and the spontaneously produced SO2− radical react with the NBD-PE–labeled lipids of the outer membrane layer and produce nonfluorescent derivatives (27). They diffuse very slowly through the bilayer and thus allow the quantitative distinction of the inner- and outer-layer lipids. In our NBD-PE–labeled vesicle preparations, unilamellarity was verified by a 51% ± 4% (zwitterionic liposomes) and 55% ± 4% (cationic liposomes) decrease of the initial fluorescence on dithionite addition.
Liposome Stability Measurements
To study the liposome stability (retention of entrapped contents), a fluorescent dye (calcein) was encapsulated at self-quenching concentrations. Different liposome sizes (100-, 400-, and 800-nm filter pore diameter) were examined. The fluorescence intensity of liposome suspensions was measured by a fluorescence microplate reader (excitation wavelength, 485 nm; emission wavelength, 538 nm). Destabilization of the liposomal membrane causes calcein leakage from the liposomes. Calcein leakage is followed by dilution of calcein into the surrounding solution and relief of the fluorescence self-quenching effect, which results in increase of fluorescence intensity. To normalize and properly compare different samples, Triton X-100 (4.5% w/w) was added to the suspension to achieve complete calcein release. To characterize liposome stability, the fluorescence self-quenching efficiency q of liposome suspensions was compared over the period of 30 d (q = Imax/I, where I is the measured fluorescence intensity before Triton X-100 addition and Imax is the maximum fluorescence intensity after Triton X-100 addition).
Liposome Size Distribution Determination
Dynamic light scattering (DLS) of liposome suspensions was studied with an N4 Plus autocorrelator (Beckman-Coulter), equipped with a 632.8-nm He–Ne laser light source. Scattering was detected at 15.7°, 23.0°, 30.2°, and 62.6°. Particle size distributions at each angle were calculated from autocorrelation data analysis by CONTIN (28). The average liposome size was calculated to be the y-intercept at zero angle of the measured average particle size values versus sin2(θ) (29). All buffer solutions used were filtered with 0.22-μm filters just before liposome preparation. The collection times for the autocorrelation data were 1–4 min.
Transmission Electron Microscopy
To observe the size and external morphology of liposomes, a transmission electron microscope (JEOL) was used at 80 kV following the negative staining method. Liposome suspensions were added dropwise to a 400-mesh copper grid coated with polyvinyl formal. After allowance for liposome adhesion, excess sample was removed with filter paper. Staining was obtained with isotonic uranyl acetate solution (2%) in phosphate buffer.
Actinium and Bismuth Retention Measurements
As shown on Figure 1, 225Ac (t1/2 = 10 d) decays to the following α-emitting radionuclides (respective half-lives shown): 221Fr (4.9 min), 217At (32 msec), and 213Bi (45.6 min). To experimentally test the retention of 225Ac and its daughters by the liposomes, the γ-emissions of 221Fr and 213Bi were measured using a Cobra γ-counter (Packard Instrument Co., Inc.). Under steady-state conditions (after 24 h), the decay rate of each species in the decay chain must be equal; thus, at steady state, the decay rate of either francium or bismuth can be used to determine the actinium concentration. Measurements of the kinetics of 213Bi activity (t1/2 is 45.6 min) in liposome fractions separated from the parent liposome population and the unentrapped radionuclides, at different times, allow for estimation of the stability of 213Bi entrapment. In particular, if 213Bi is not leaking, then the 213Bi activity concentration (measured by γ-counting), after SEC, would be the same over several hours of measurement. If 213Bi is leaking, the 213Bi γ-emissions from the liposome fraction will be low initially, since 213Bi has been lost from the liposomes: Had bismuth been retained, the radioactivity of the vesicle fraction would have been at steady state with 225Ac (i.e., at ∼100%). In this newly separated liposome fraction, the bismuth activity rises with monoexponential kinetics (with kinetic constant almost equal to 213Bi half-life) as ongoing actinium decay gradually brings the entire fraction into steady state. Extrapolation of the monoexponential fit curve to the time point of chromatographic separation gives the fractional 213Bi retention by the liposomes. For each liposome population, 213Bi activity was measured after repeated SEC (Sephadex G-50) at various time points. The statistical significance of differences in 213Bi retention for different-sized liposomes was determined by obtaining the area under the retention-versus-time curve for each measurement set. The statistical significance of differences in area under the retention-versus-time curve were then evaluated by a Student (2-tailed) t test (SigmaPlot, SPSS Inc.)
Theoretical Model
To evaluate the theoretical limits of daughter entrapment, a model was developed using nuclear theory and geometry. The nuclear recoil distances of the α-decays of the actinium daughters are not well established in aqueous media, but estimates to within 10%–20% were made (30) using a standard computer model. The range of the recoil distances for francium, astatine, and bismuth were estimated as 81.7, 86.5, and 94.7 nm, respectively. Because these differ by less than the experimental uncertainties, both in the recoil range estimates and in our experimental measurements, a single recoil distance was used to simplify further calculations. With this model, unilamellar liposomes were considered. Each entrapped radionuclide was distributed uniformly within the aqueous volume of a liposome. Thus, each disintegration has a fixed probability f for daughter ejection. The probability of ejection fr,rd for a disintegration, which occurs at a distance r from the center of a liposome of radius rl, may then be calculated from geometry (rd is the recoil distance). The probability f is calculated by averaging the ejection probability fr,rd over the possible locations of the radionuclide, r:
Retention of bismuth requires 3 successive decays without escape; therefore, the probability is f3.
Binding Measurements of 225Ac and 225Ac-DOTA to Liposomes
Different activities of 225Ac and 225Ac-DOTA, ranging from 10 Bq to 6 kBq, were mixed in PBS with sucrose-loaded liposomes (2 mmol/L total lipid), and the system was allowed to equilibrate for at least 1 h at room temperature. Rhodamine-labeled zwitterionic and cationic liposomes were present at the same concentrations as those used for the measurement of 225Ac and 213Bi retention by the above liposomes. After ultracentrifugation for 2 h at 142,000g, (25°C), the supernatant (the top 90% of the sample volume) was promptly removed. Lipid in both supernatant and pellet was measured using the rhodamine fluorescence intensity (excitation wavelength, 544 nm; emission wavelength, 590 nm). 225Ac activity concentration was measured by the γ-emissions of 213Bi decay at equilibrium. The calculated amount of membrane-bound 225Ac was corrected for the amount of lipid that remained in the supernatant as follows: β, the fraction of bound 225Ac, was defined as the ratio of the bound 225Ac to the total 225Ac in the sample. It was assumed that the concentrations of unbound 225Ac in the pellet [Ac]p,f and in the supernatant were equal, since the distribution of free 225Ac atoms along the tube axis should not be influenced by the separation process. Thus, for the pellet, the average amount of 225Ac bound per lipid (B) would be B = ([Ac]p,t –[Ac]p,f)/[L]p, where [Ac]p,t is the total concentration of 225Ac in the pellet and [L]p is the lipid concentration in the pellet. Because not all the liposomes were found in the pellet, B was used to calculate the bound 225Ac in the supernatant and the true free 225Ac concentration.
RESULTS
Liposome Characterization
Liposomes were prepared by the extrusion method with different filter pore sizes. Liposome size distributions were determined with DLS. The measured average liposome sizes for the zwitterionic composition were 183 ± 83 nm (100-nm filter diameter), 402 ± 185 nm (400-nm filter diameter), and 646 ± 288 nm (800-nm filter diameter). For the cationic composition the corresponding values were 184 ± 74 nm (100-nm filter diameter), 415 ± 153 nm (400-nm filter diameter), and 602 ± 385 nm (800-nm filter diameter). Liposome size distributions were stable during a period of 30 d (data not shown). Liposomes were also imaged by transmission electron microscopy using the negative staining method (Fig. 2).

Negative-staining transmission electron microscopy images of liposomes prepared with 800-nm (A) and 100-nm (B) filter diameter.
To evaluate the stability of the liposomal membrane over time, calcein was entrapped into liposomes at high concentration. Release of calcein, because of liposome instability, results in dilution of the fluorophore into the surrounding buffer solution, relief of self-quenching, and an increase in the fluorescence intensity. The fractional fluorescence self-quenching decrease, Δq × 100, is shown in Table 1 for zwitterionic and cationic liposomes during the 30 d. Both liposome formulations and all liposome sizes showed a small release of calcein within the first 24 h after preparation; this may have been caused by differences in osmolarity between the encapsulated calcein solution and the liposome surrounding solvent or by membrane defect relaxation. After the second day, and during the 30 d, all types of liposomes were stable. The same behavior was observed for liposomes in media with 1%–10% serum at 37°C (data not shown). The effect of radiation on the stability of liposomes was studied by DLS. The liposome size distributions of samples that contained 225Ac did not change over time.
Percentage Decrease in Self-Quenching with Time
Retention of 225Ac by Liposomes
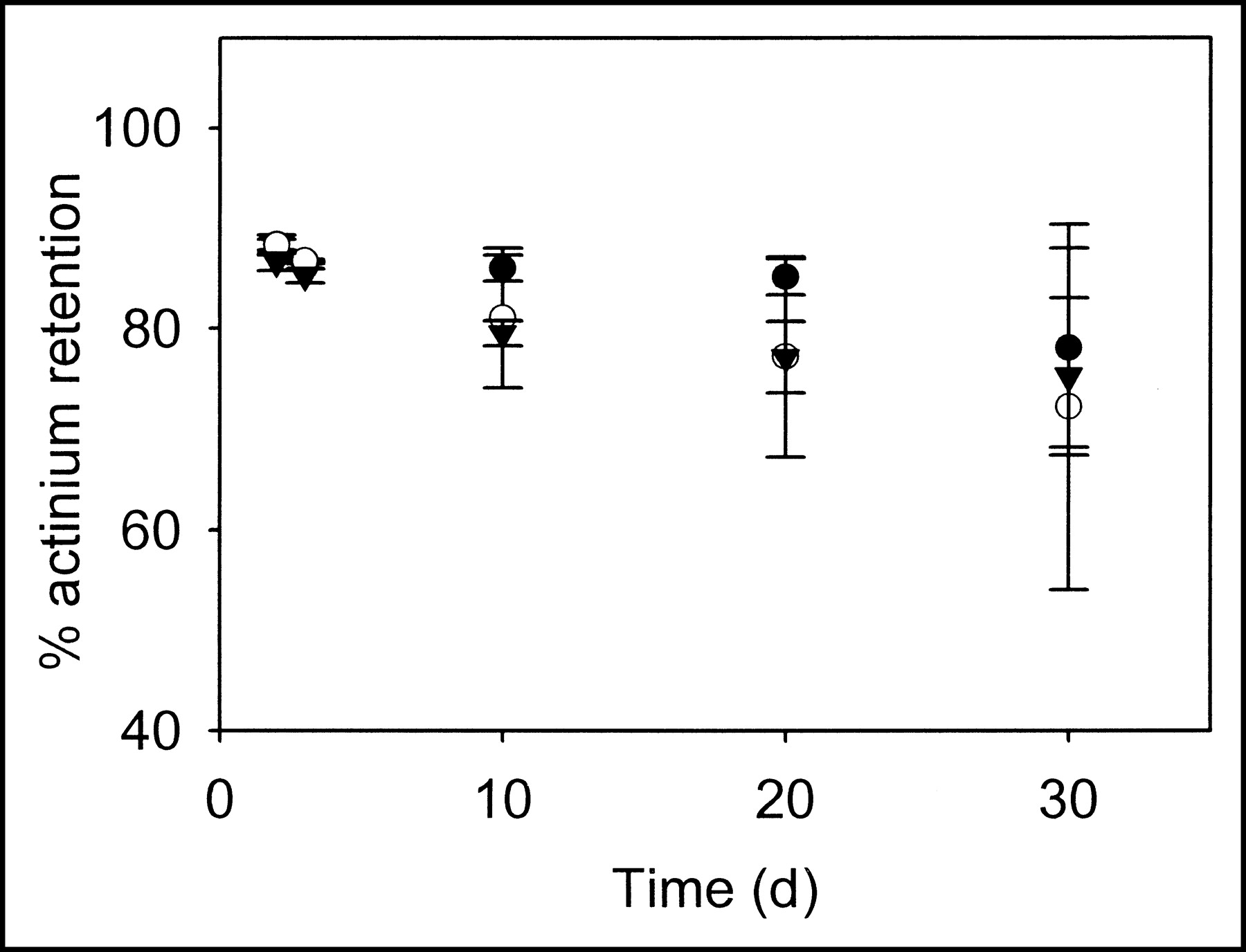
As stated in the Materials and Methods, 225Ac retention in liposomes can be determined by the 221Fr or 213Bi activities at steady state. At different time points, liposome fractions were separated from free 225Ac by SEC, and the γ-photons of 213Bi decay at steady-state were used to determine 225Ac retention. Independent of time, the 225Ac activity retained in the zwitterionic liposome fractions was >88% (Fig. 3). 225Ac retention decreased (Fig. 4) over time in the cationic liposomes (cationic lipid: 10-mol percentage of total lipid), but even after 30 d the retention was more than 54%. 225Ac mean encapsulation efficiency in liposomes, using passive entrapment, was 6.4%, with a maximum value of 10.0%.

Fraction of 225Ac retention by zwitterionic liposomes (filter diameters: 100 (•), 400 (○), and 800 (▾) nm) during 30 d. The error bars correspond to SEs of repeated measurements.
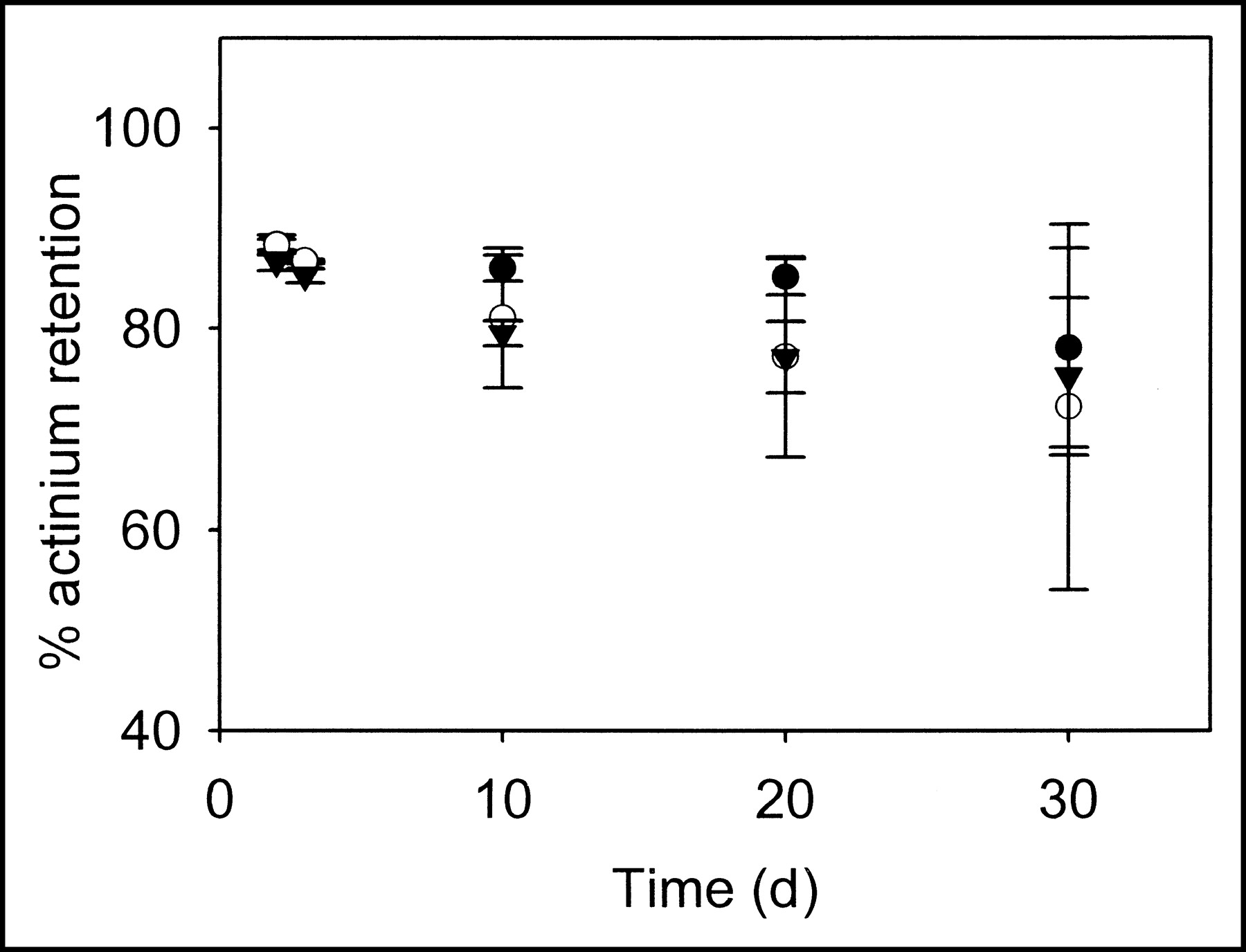
Fraction of 225Ac retention by cationic liposomes (filter diameters: 100 (•), 400 (○), and 800 (▾) nm) during 30 d. The error bars correspond to SEs of repeated measurements.
Theoretical Calculations
The toxicity of 225Ac is in part due to release of its α-particle–emitting daughters. Retention of daughters within the liposomes and at the tumor site would enhance the potential of 225Ac toward α-particle therapy.
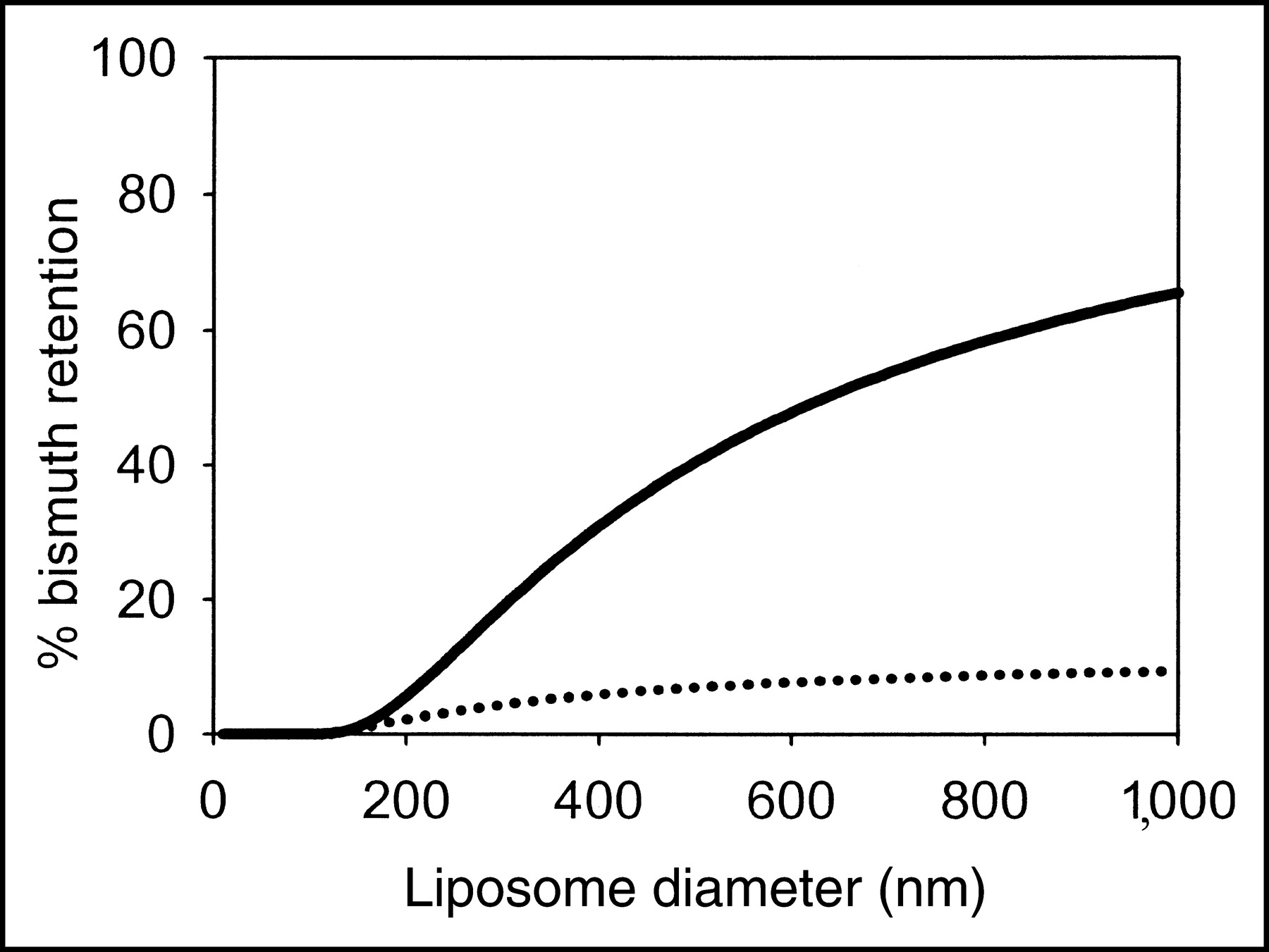
The mechanism for daughter loss from the liposome interior is likely to be nuclear recoil. The theoretical fraction of 213Bi retention as a function of liposome size is shown in Figure 5 assuming a recoil distance of 87.6 nm—the average recoil distance in water calculated using the software program Stopping and Range of Ions in Matter, or SRIM. 213Bi is the last α-emitting intermediate on the 225Ac scheme, and thus retention of 213Bi requires retention of both 221Fr and 217At. Adequate 213Bi retention (>50%) requires large liposomes (>650 nm in diameter) (Fig. 5). Theoretical retention was also calculated assuming that 225Ac and its daughters localize at the inner liposome membrane; this represents a worst-case scenario. In this case, the maximum 213Bi retention approaches 1/23 = 12.5% for large liposomes (8.1% for 650-nm-diameter liposomes). Half the recoils from the surface of an infinitely large vesicle would result in daughter ejection. Thus, if even a fraction of the daughters of 225Ac (or of 225Ac itself) associates with the liposomal membrane, the 213Bi retention will be significantly reduced.
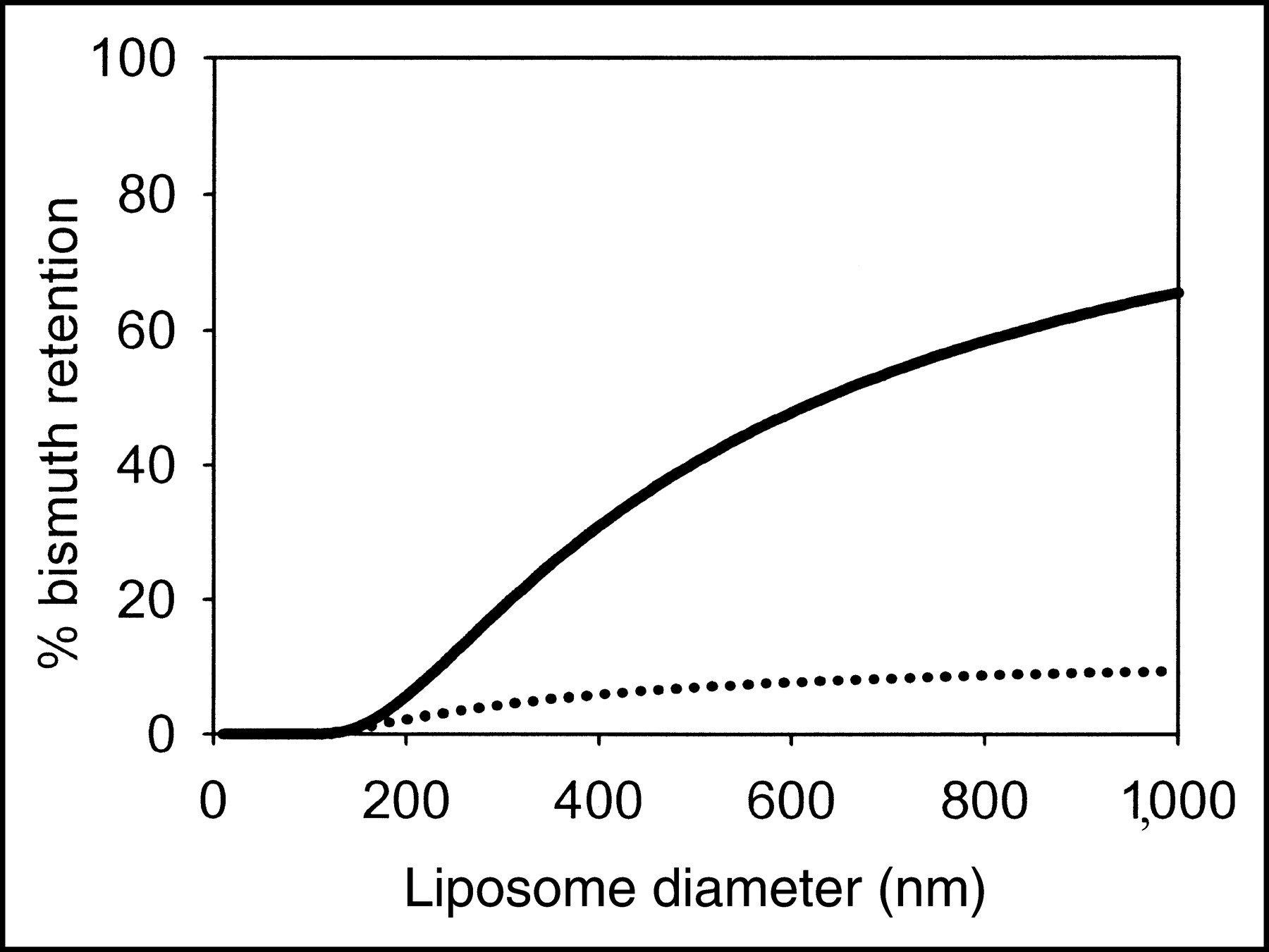
Theoretical predictions of 213Bi retention for different liposome sizes (solid line). Radionuclides were assumed to be uniformly entrapped into the liposomal aqueous compartment. The average recoil distance of 87.6 nm was used for all α-emitting intermediates to simplify calculations. Binding of radionuclides to the liposomal membrane will significantly reduce retention (dotted line).
Retention of 213Bi by Liposomes
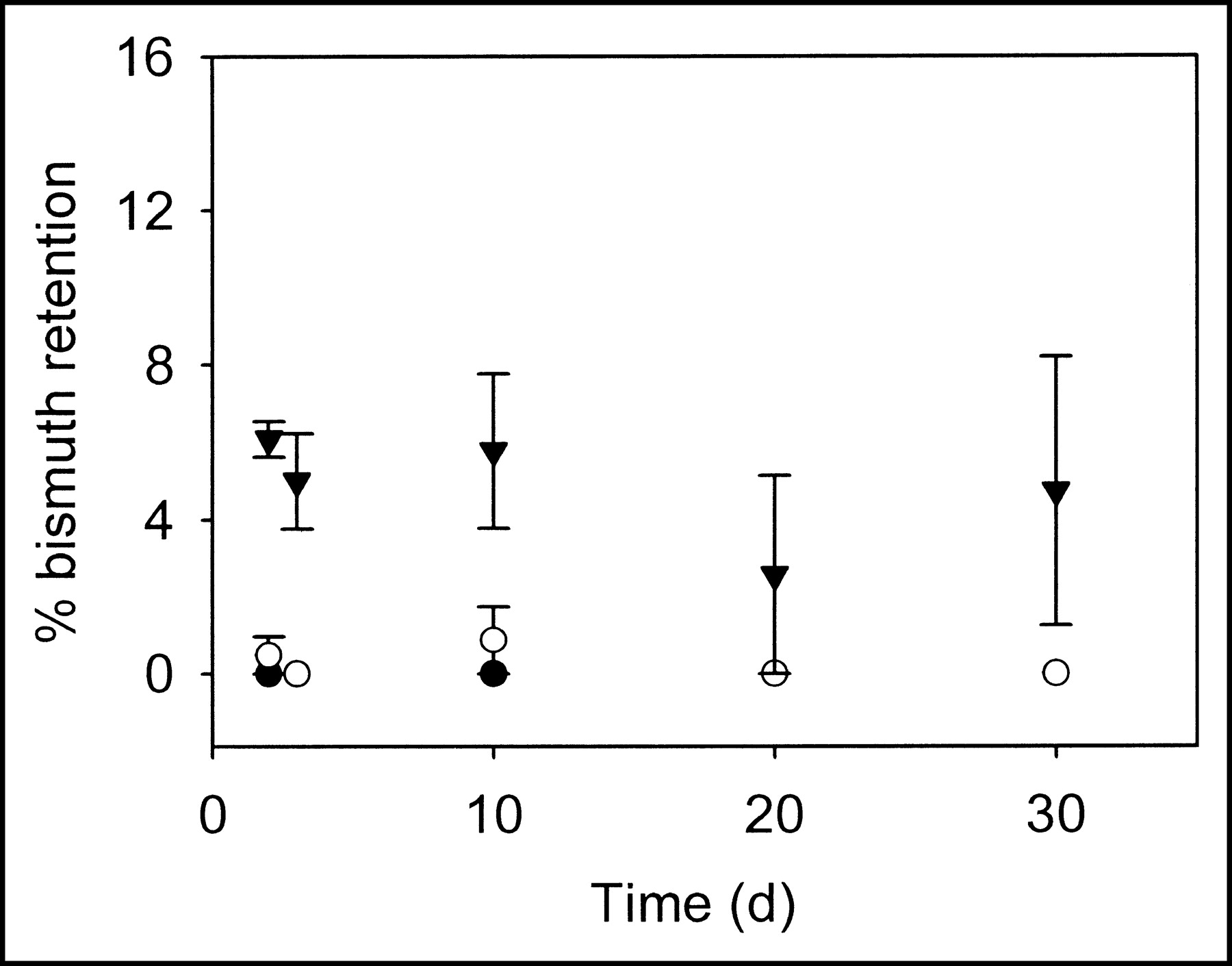
The retention of 213Bi by liposomes was studied as a function of liposome size and composition. For each liposome population, measurements of 213Bi activity were made after repeated chromatography at various time points (Figs. 6 and 7). For large liposomes, retention was much less than theoretically predicted. For both liposome compositions and all sizes examined, the values of 213Bi retention were consistent with the theoretical results for localization of the radionuclides on the liposome membrane.

Fraction of 213Bi retention by zwitterionic liposomes (filter diameters: 100 (•), 400 (○) and 800 (▾) nm) during 30 d. The error bars correspond to SEs of repeated measurements (for some data points, error bars are smaller than the symbol sizes). For day 30, 213Bi retention for 100- and 400-nm liposomes is 0%, and data points overlap. 213Bi retention in 800-nm liposomes was significantly higher than that in 100-nm liposomes (P = 0.0044).
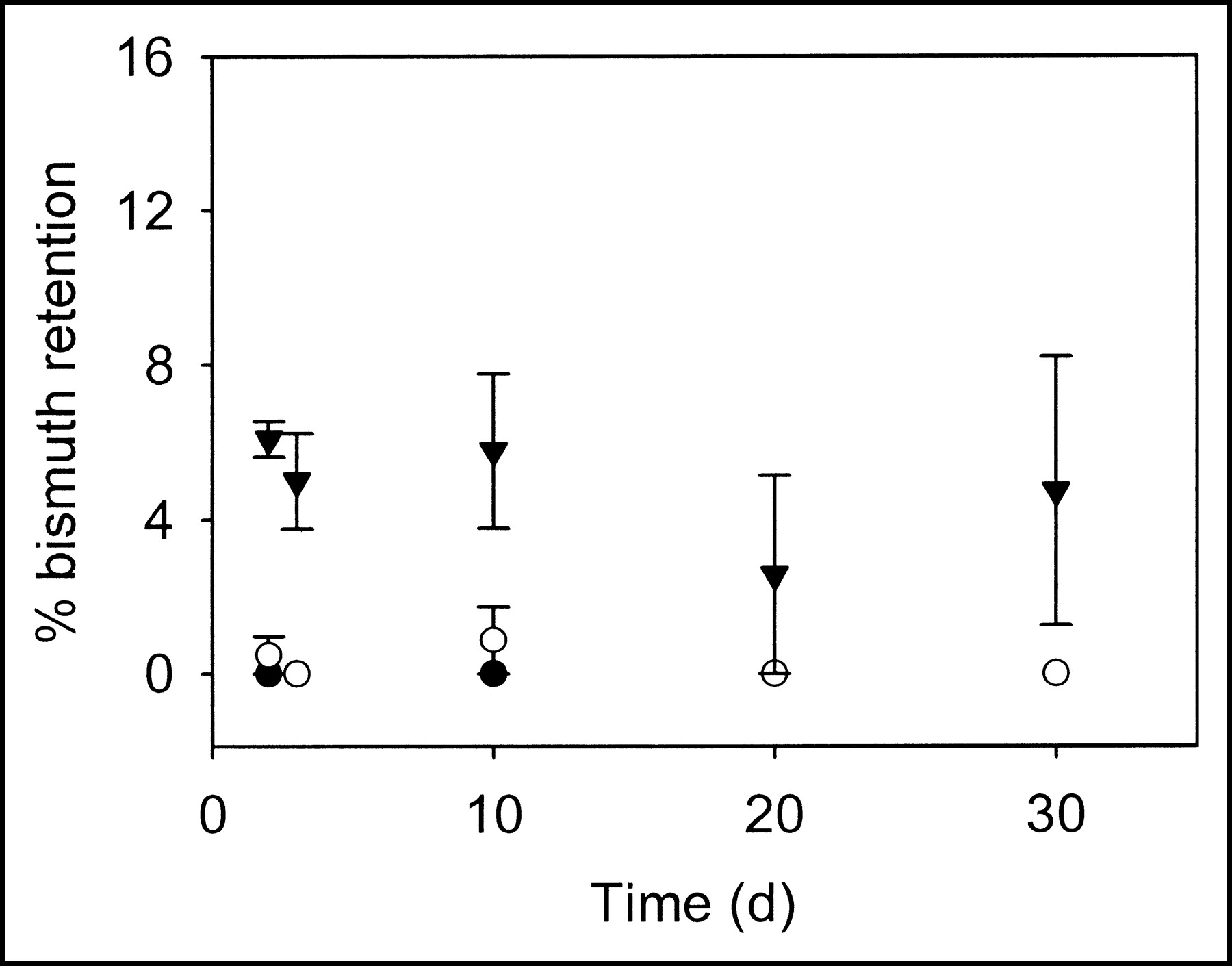
Fraction of 213Bi retention by cationic liposomes (filter diameters: 100 (•), 400 (○) and 800 (▾) nm) during 30 d. The error bars correspond to SEs of repeated measurements (for some data points, error bars are smaller than the symbol sizes). For days 20 and 30, 213Bi retention for 100- and 400-nm liposomes is 0%, and data points overlap.
Binding of 225Ac and 225Ac-DOTA to Liposomal Membranes
Binding of 225Ac and 225Ac-DOTA to zwitterionic and cationic liposomes was measured using an ultracentrifugation assay. In this binding assay, sucrose-loaded liposomes were permitted to bind 225Ac, in free or chelated form, for at least 1 h at room temperature and then were separated by ultracentrifugation. Measurement of 225Ac and lipids was performed on the pellet and supernatant, as described in the Materials and Methods. The liposome concentration was kept constant and equal to that used in the retention experiments, and the radionuclide activity was varied over 4 orders of magnitude, encompassing the activities used in the retention experiments. Significant membrane binding of 225Ac was observed with zwitterionic and cationic liposomes. The number of radionuclide atoms was low compared with the number of binding sites present (concentration of lipid molecules); thus, no saturation on the binding profile was observed. The partition constants Kp (Kp = Cbound C−1free L−1, C, L [=] mol/L) were expressed as the ratio of bound to free radionuclide atoms, normalized to the concentration of lipid present (L). For the zwitterionic membranes, Kp values were 420 (for 225Ac) and 60 (for 225Ac-DOTA), and for the cationic membranes, 550 and 50, respectively. In our measurements, 30%–35% of 225Ac was bound to membrane under the experimental conditions (accessible lipid L = 1 mmol/L).
DISCUSSION
The 225Ac atomic generator is a promising α-particle–emitting radionuclide for cancer therapy. 225Ac and its α-emitting daughters lead to the emission of a total of 4 α-particles. Loss and widespread dissemination of daughters from the site of parent generator decay will lead to toxicity, whereas confinement to the targeted site will increase efficacy (7,31). This work is part of a study that ultimately aims to target 225Ac to sites of disseminated tumor micrometastases while also retaining the 225Ac α-emitting daughters at the targeted sites. Toward this goal, the use of liposomes is proposed to deliver multiple 225Ac atoms per liposome and to retain the 225Ac progeny in the liposomes and consequently at the targeted sites.
Pegylated liposomes of different membrane charge (zwitterionic and cationic) and size were prepared to entrap 225Ac. They were characterized for content leakage (calcein release) and size (DLS) over time. Liposomes were stable during a period of 30 d. Even in the presence of radiation (from 225Ac), the liposome size distributions were stable over time. Zwitterionic liposomes retained more than 88% of 225Ac for more than 30 d. 225Ac retention in cationic liposomes was lower but still above 54%.
Passive entrapment for 225Ac encapsulation was used in this work. This approach yielded a maximum efficiency of 10% of the initial radioactivity. At initial radioactivity concentrations in the range of 11–37 MBq/mL, this efficiency translates into 10–40 actinium atoms per liposome. Immune-mediated endocytosis of such antibody-coated liposomes would deliver a much larger number of radioactive atoms to a tumor cell than would be possible with radiolabeled antibodies, given currently achievable specific activities.
An important, yet theoretically anticipated, finding of this work is that liposome size is critical in determining daughter retention, implicating nuclear recoil as a critical mechanism for daughter loss in the liposomal encapsulation strategy. Because 225Ac daughters are most probably ionic (Fr+1, At−1, Bi+3), passive permeation (diffusion) through the lipid membrane will be negligible. Based on a geometric model, large, 650-nm-diameter, liposomes are required to yield >50% retention of 213Bi, the last α-emitting daughter in the decay chain. For both liposome compositions and all sizes of liposomes examined, the values of 213Bi retention were consistent with the theoretical results for localization of the radionuclides on the liposome membrane. 213Bi retention was significantly higher in the 800- versus the 100-nm zwitterionic liposomes. No statistically significant difference was seen between 400- and 800-nm zwitterionic liposomes. Likewise, no statistically significant difference in 213Bi retention was seen between the 800- and 100-nm liposomes among the different cationic liposomes. Measurement of the extent of association of Fr+1, At−1, and Bi+3 to liposomal membranes is not possible because of their short half-lives compared with the length of the measurement process. Nevertheless, partitioning of the ionic α-emitting intermediates to the liposomal membrane is possible. Binding or association of inorganic monovalent ions to noncharged hydrophilic surfaces, such as phospholipid membranes, has been treated theoretically (32) and verified experimentally (33). In addition, certain anions have the tendency to associate with phosphatidylcholine lipids (32). The partition of Ac+3 and Ac+3-DOTA was measured, however, and only a small fraction of the chelated 225Ac partitioned to the liposomal membrane. On the contrary, the measured partitioning of unchelated Ac+3 to zwitterionic and cationic membranes was significantly higher. Continuing work focuses on the improvement of 213Bi retention by modification of the membrane affinity.
The theoretical calculations, confirmed by the experimental results, showed that relatively large liposomal sizes are required for adequate daughter retention. (The relatively large liposome sizes required, 650 nm in diameter, are still much smaller than the average cell size, 20 μm. Cells are typically 10,000-fold greater in volume than the largest liposomes considered in this work.) The application of these liposomal systems will focus on locoregional and intracavitary therapy of disseminated micrometastases (34). Liposomes of large size are rapidly cleared from blood circulation (35). Future developments may contribute to better understanding of the clearance mechanism. Targeted delivery can in principle be achieved with appropriate antibodies attached at the terminal end of the PEG chains; this geometry provides exposed antibody molecules that protrude from the liposome for unhindered antigen recognition. Similarly engineered liposomes have been shown to achieve increased cellular uptake by receptor-mediated endocytosis (36).
CONCLUSION
225Ac is a highly potent, but also a potentially highly toxic, α-emitting atomic generator. 225Ac is the parent in a chain of 4 α-particle emitters (221Fr, 217At, 213Bi) and is thus a good candidate radionuclide for cancer therapy. To investigate this approach, multiple 225Ac atoms per liposome were successfully encapsulated within phosphatidylcholine-cholesterol pegylated vesicles and were stably retained for more than 30 d.
Retention of 225Ac radioactive daughters at the tumor site will provide 225Ac with exceptional potential toward α-particle therapy. It was shown that daughter retention depends on liposome size because of daughter nuclear recoil. Measurements of 213Bi retention by liposomes suggested that the α-emitting daughters apparently localize at the liposomal membrane and thus significantly reduce 213Bi retention. Further, improvement of 213Bi retention is being investigated. This work will make it possible to retain the high potency of 225Ac while reducing the toxicity, thereby making it a more feasible approach for cancer therapy.
Acknowledgments
We thank Dr. William Tong (Rockefeller Research Laboratories), Prof. Iwao Teraoka (Polytechnic University), and Dr. Hedy Druskin (Rockefeller Research Laboratories) for use of their laboratory facilities; Drs. Richard Enmon and Stig Palm for many helpful discussions; and Dr. Kostas Kostarelos for drawing our attention to the use of liposomes as carriers of radioactivity. This study was supported by concept award DAMD170010657 from the U.S. Army Medical Research and Materiel Command, grant R01 CA55349 from the National Institutes of Health, the Doris Duke Charitable Foundation, the Experimental Therapeutics Center, the Goodwin Commonwealth Foundation for Cancer Research, and the Dr. Frederick E.G. Valergakis Graduate Research Grant of the Hellenic University Club of New York.
Footnotes
Received May 12, 2003; revision accepted Oct. 9, 2003.
For correspondence or reprints contact: Stavroula Sofou, PhD, Department of Medical Physics, Memorial Sloan-Kettering Cancer Center, 1275 York Ave., New York, NY 10021.
E-mail: sofous{at}mskcc.org
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Anti-Prostate-Specific Membrane Antigen Liposomes Loaded with 225Ac for Potential Targeted Antivascular {alpha}-Particle Therapy of Cancer
- Immunoliposomal Delivery of 213Bi for {alpha}-Emitter Targeting of Metastatic Breast Cancer
- Radioimmunotherapy of Breast Cancer Metastases with {alpha}-Particle Emitter 225Ac: Comparing Efficacy with 213Bi and 90Y
- Cancer Nanotargeted Radiopharmaceuticals for Tumor Imaging and Therapy
- Liposome-Mediated Radiotherapeutics Within Avascular Tumor Spheroids: Comparative Dosimetry Study for Various Radionuclides, Liposome Systems, and a Targeting Antibody