


Abstract
Recent studies have revealed that the wild-type hemochromatosis protein (HFE) interacts with the transferrin receptor (TfR) and modulates TfR-mediated iron uptake by cells. Because of similarities in the transport of gallium and iron and the use of 67Ga scanning in lymphoid malignancies, we examined the effect of HFE expression on 67Ga uptake. Methods: 67Ga and 59Fe uptakes were measured in HeLa cells transfected with a FLAG-tagged wild-type HFE (fHFE) gene under control of a tetracycline-repressible promoter. fHFE and TfR protein levels were measured by Western blotting; cellular transferrin (Tf) binding sites were measured by 125I-Tf binding assay. Results: Induction of fHFE expression produced an increase in TfR protein that was accompanied by a decrease, rather than an increase, in cellular 67Ga and 59Fe uptake. The difference in 67Ga uptake between fHFE-expressing and fHFE-nonexpressing cells was markedly increased in the presence of Tf. Although fHFE expression produced an increase in cellular TfR protein, cell surface and intracellular Tf binding sites were actually decreased in these cells. Conclusion: Our studies suggest that expression of wild-type HFE in cells produces a decrease in 67Ga uptake due to a reduction in available Tf binding sites for 67Ga-Tf on the TfR. These results imply that 67Ga uptake by cells with wild-type HFE may differ from cells with the HFE C282Y mutation.
Scanning with 67Ga is of value in the detection of viable malignant cells in tumor masses that persist after treatment of patients with Hodgkin’s and non-Hodgkin’s lymphoma (1,2). The mechanisms involved in gallium uptake by cells have been the subject of considerable investigation. Gallium resembles iron in many ways in that it circulates in the blood bound to transferrin (Tf), the iron transport protein, and is incorporated into cells by Tf receptor (TfR)-mediated endocytosis of 67Ga-Tf complexes (3–5). In addition, TfR-independent uptake of 67Ga by cells has been described (3–5).
Hemochromatosis, an inherited disorder of iron overload, is one of the most common autosomal recessive diseases in individuals of northern European ancestry, with an occurrence of 1 in 300 and a carrier state of 1 in 10 persons (6). It is characterized by increased iron absorption from the gut and the deposition of excess iron in certain organs, including the liver, heart, and pancreas, resulting in tissue damage, morbidity, and mortality (7). Recent studies of the hemochromatosis (HFE) gene have revealed that in the cell, wild-type HFE protein binds β2-microglobulin and trafficks to the cell surface in association with the TfR (8,9). Wild-type HFE protein bound to the TfR competitively interferes with the binding of Tf-Fe to the TfR, thus decreasing TfR-mediated uptake of iron (10,11). The HFE C282Y mutation (substitution of cysteine by tyrosine at amino acid 282) is the predominant HFE mutation responsible for hemochromatosis (12); it results in the production of an HFE protein that fails to bind to the TfR (13).
The modulating effect of HFE on cellular iron uptake raises important questions regarding the potential impact that this gene may have on the uptake of gallium by cells. To address this issue, we conducted studies to examine 67Ga uptake by HeLa (human cervical cancer) cells under conditions of inducible expression of the wild-type HFE gene. Our studies show that changes in the expression of HFE significantly affect the uptake of gallium by these cells.
MATERIALS AND METHODS
Materials and Cells
Human Tf, anti-FLAG monoclonal antibody M2, puromycin, and doxycycline were purchased from Sigma Chemical Co. Geneticin (G418) was purchased from GIBCO BRL. Anti-TfR monoclonal antibody was purchased from Zymed Laboratories. 67Ga citrate was obtained from NycoMed-Amersham. 59Fe and Na125I were purchased from Amersham, and 59Fe-Tf and 125I-Tf were prepared as described (14). The fwtHFE/tTA HeLa cell line expressing FLAG epitope-tagged wild-type HFE (fHFE) under control of the tetracycline-responsive promoter was used in our studies and was generously provided by Dr. Caroline A. Enns (University of Oregon). These cells have been described (8). Cells were grown in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum, 400 μg/mL geneticin, and 200 ng/mL puromycin. fHFE gene expression in cells was turned off by the addition of 1 μg/mL doxycycline to the culture medium and turned on by the removal of doxycycline. Cells were incubated at 37°C in an atmosphere of 6% CO2.
Western Blotting
fHFE and TfR protein levels in cells were determined using an ECL Western blotting detection system (Amersham). Cell lysates were prepared, and total cellular proteins were resolved on a 10% (for HFE) or 7.5% (for TfR) NaDodSO4-polyacrylamide gel and transferred to nitrocellulose membrane using a Transblot system (BioRad, Richmond, CA). Membranes were incubated in blocking buffer (phosphate-buffered-saline [PBS] with 0.1% Tween-20 containing 10% nonfat dry milk), followed by sequential washes in PBS-Tween. Membranes were then incubated in PBS-Tween containing 2 μg/mL of monoclonal anti-FLAG antibody or anti-TfR antibody and then washed and incubated for 1 h at room temperature in buffer containing rabbit antimouse immunoglobulin (1:10,000 dilution) conjugated to horseradish peroxidase. For detection of bands, membranes were immersed in ECL detection solution and exposed to XAR-5 film for autoradiography.
Cellular Uptake of 67Ga and 59Fe and 125I-Tf Binding Studies
fwtHFE/tTA HeLa cells grown to confluency in flasks with or without 1 μg/mL doxycycline were harvested and replated at a concentration of 5 × 105 cells/mL in 24-well plates. After cells had been incubated for 24 h in a CO2 incubator, 67Ga citrate (0–74 kBq [0–2.0 μCi]) and various concentrations of Tf (0, 1, or 10 μg/mL) were added to the wells. After 24 h of incubation with 67Ga citrate, wells were washed with ice-cold PBS to remove unincorporated 67Ga. Cells were removed from the wells, and the total amount of 67Ga radioactivity in the cells was counted using a gamma counter.
For studies of 59Fe uptake, cells grown in the presence or absence of doxycycline were replated in 24-well plates at a concentration of 5 × 105 cells/mL. 59Fe-Tf (final Fe concentration, 8 ng/mL) was added to each well at the start of the incubation. After 24 and 48 h of incubation, cells were harvested and washed by centrifugation with PBS. The total 59Fe cpm in the cell pellet was counted to determine the amount of radioactive iron taken up by cells.
Tf binding to cells grown with or without doxycycline was measured using an 125I-Tf binding assay. Cells were harvested and washed twice with ice-cold PBS containing 0.1% bovine serum albumin, and binding studies were performed on 5 × 105 cells in 500 μL at 4oC and 37°C as described (14).
RESULTS
Induction of Wild-Type fHFE Expression Increases TfR Levels
To confirm that fHFE gene expression could be switched on or off in the cells used in our studies, cells were analyzed for the presence of fHFE protein by Western blot analysis after a 96-h incubation with or without doxycycline. As shown in Figure 1A, HFE expression was induced in the absence of doxycycline but was turned off in the presence of 1 μg doxycycline. Since prior studies have demonstrated an important role for the TfR in gallium uptake, cells were also analyzed for TfR expression by Western blotting. As shown in Figure 1B, lanes 1–3, the induction of fHFE expression resulted in an increase in cellular TfRs.
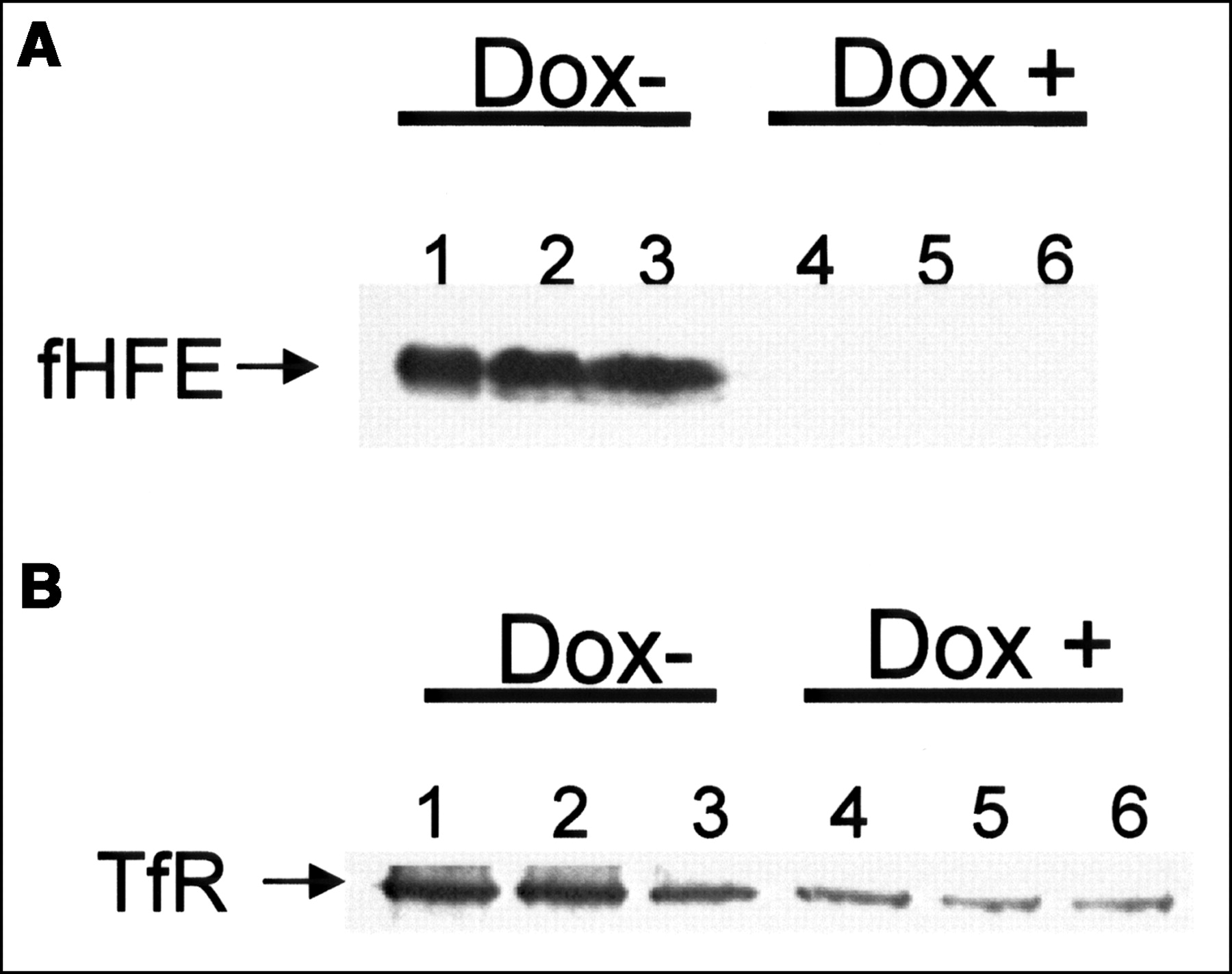
Effect of wild-type fHFE expression on TfR expression. (A) Western blot shows inducible expression of fHFE protein. Cells were incubated for 96 h in medium in absence (Dox -) or presence (Dox +) of 1 μg/mL doxycycline. Total cellular protein was loaded on gel as follows: 30 μg in lanes 1 and 4; 60 μg in lanes 2 and 5; and 120 μg in lanes 3 and 6. (B) Western blot shows that induction of fHFE expression results in increase in cellular TfR. Dox - (lanes 1–3) represents cells grown in absence of doxycycline; Dox + (lanes 4–6) represents cells grown in presence of doxycycline. All lanes were loaded with 35 μg protein from cell lysates prepared from separate incubations.
Effect of fHFE Expression on 67Ga Uptake
67Ga uptake by cells with or without fHFE switched on was measured using a range of 67Ga concentrations and exogenously added Tf. As shown in Figure 2A–2C, after 24 h of incubation, cells that expressed fHFE incorporated a significantly lower amount of 67Ga than cells in which fHFE was not expressed. In the initial experiment shown in Figure 2A, 67Ga uptake was measured in cells incubated in serum-supplemented medium that contained bovine Tf (approximately 0.2 μg/mL). However, bovine Tf is known to have a much lower affinity for the human TfR (present on HeLa cells) than human Tf (15). Therefore, subsequent 67Ga uptake experiments (Fig. 2B and 2C) were conducted in the presence of exogenously added human Tf. As shown in these figures, the differences in 67Ga uptake between fHFE-expressing and fHFE-nonexpressing cells was more pronounced in the presence of 1 and 10 μg of human Tf, (Figs. 2B and 2C) than in the absence of exogenously added Tf (Fig. 2A).
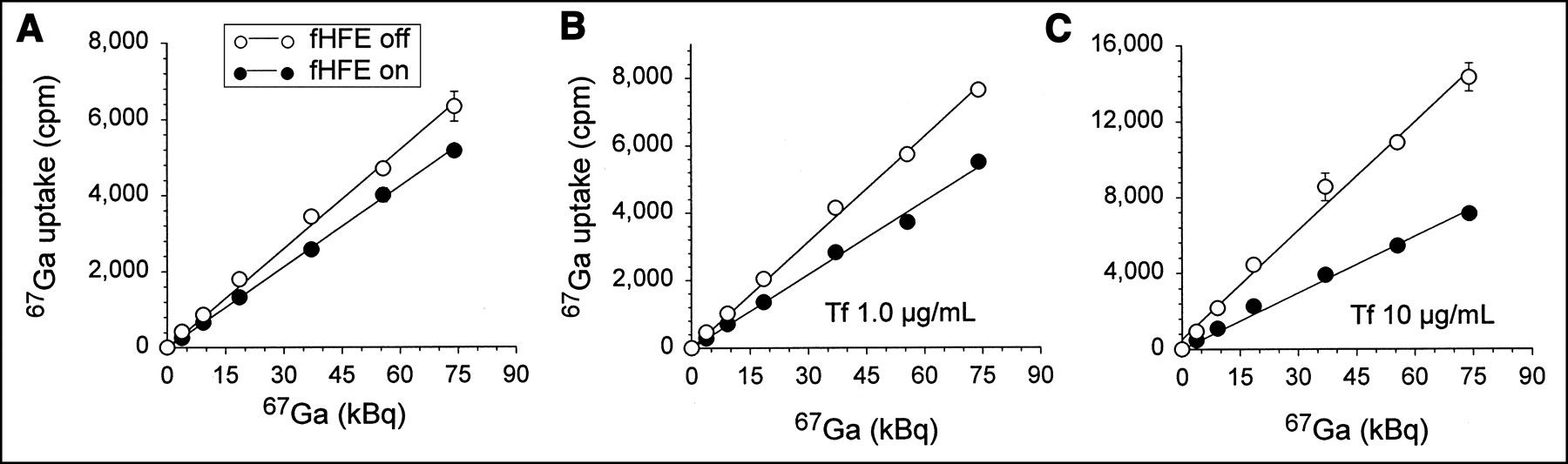
Effect of fHFE expression on 67Ga uptake by cells. Cells were incubated for 96 h in absence of doxycycline to induce fHFE gene expression (fHFE on) or in presence of 1 μg/mL doxycycline to repress fHFE expression (fHFE off). Cells were then replated in medium containing increasing concentrations of 67Ga, and amount of radioactivity taken up by cells was determined after 24-h incubation. (A) 67Ga uptake in complete medium. (B and C) 67Ga uptake in complete medium containing 1 μg/mL (B) or 10 μg/mL (C) of exogenously added human Tf. Values shown represent mean ± SE of representative experiment performed in triplicate. Differences in 67Ga uptake between cells with fHFE turned off and on are significant (P < 0.01, by t test) for all data points above 15 kBq 67Ga.
Effect of fHFE Expression on Tf Binding Sites and Iron Uptake
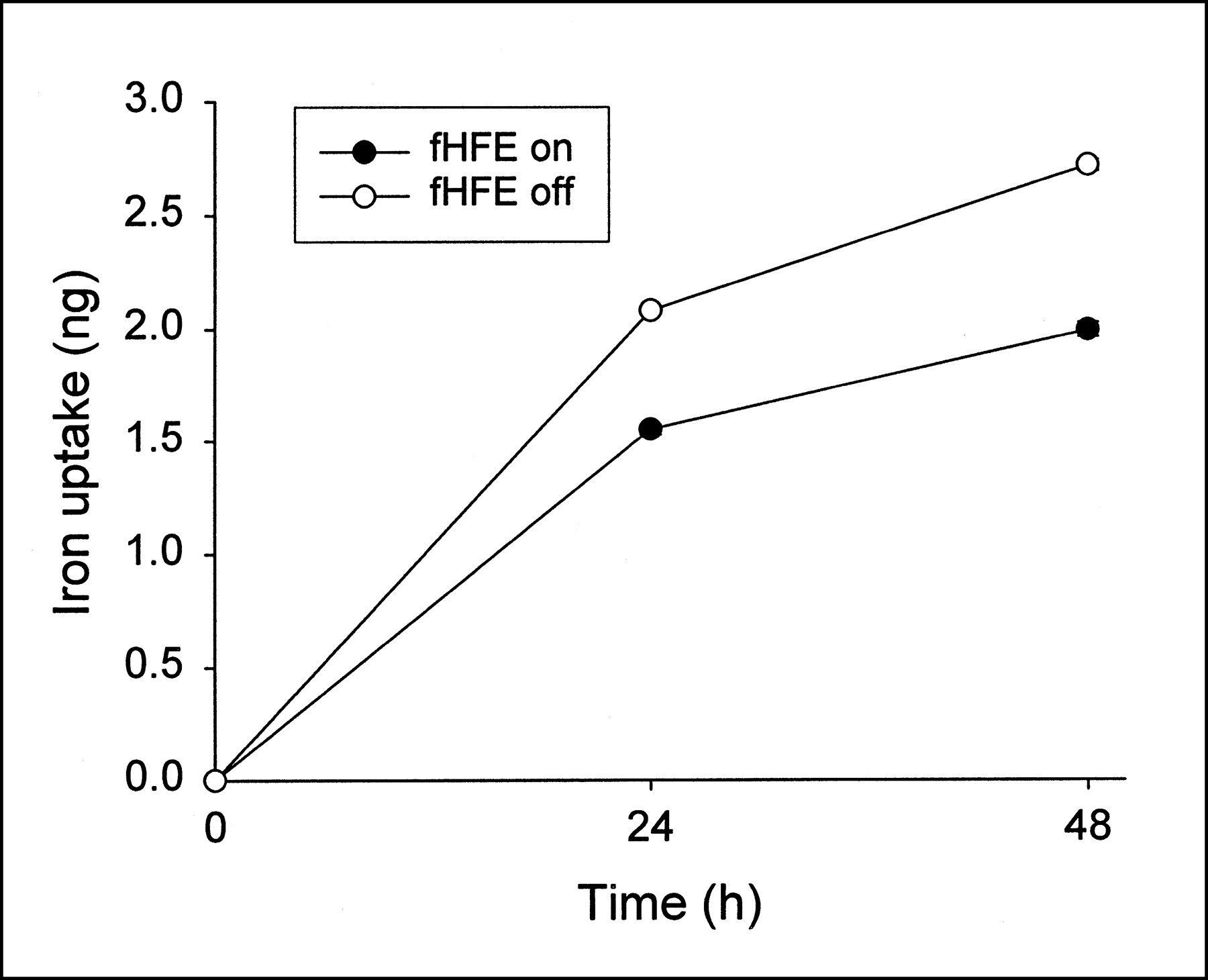
In contrast to the increase in TfR protein seen on immunoblotting with the induction of fHFE, 125I-Tf binding assay revealed that cells expressing fHFE had a lower number of total cellular and cell surface Tf binding sites (measured at 37°C and 4°C, respectively) when compared with cells in which fHFE was not induced (Fig. 3). To confirm that the decrease in Tf binding sites associated with fHFE expression had physiologic significance, iron uptake by cells was measured. As shown in Figure 4, the induction of fHFE expression resulted in a significant reduction in 59Fe accumulation in cells over time.
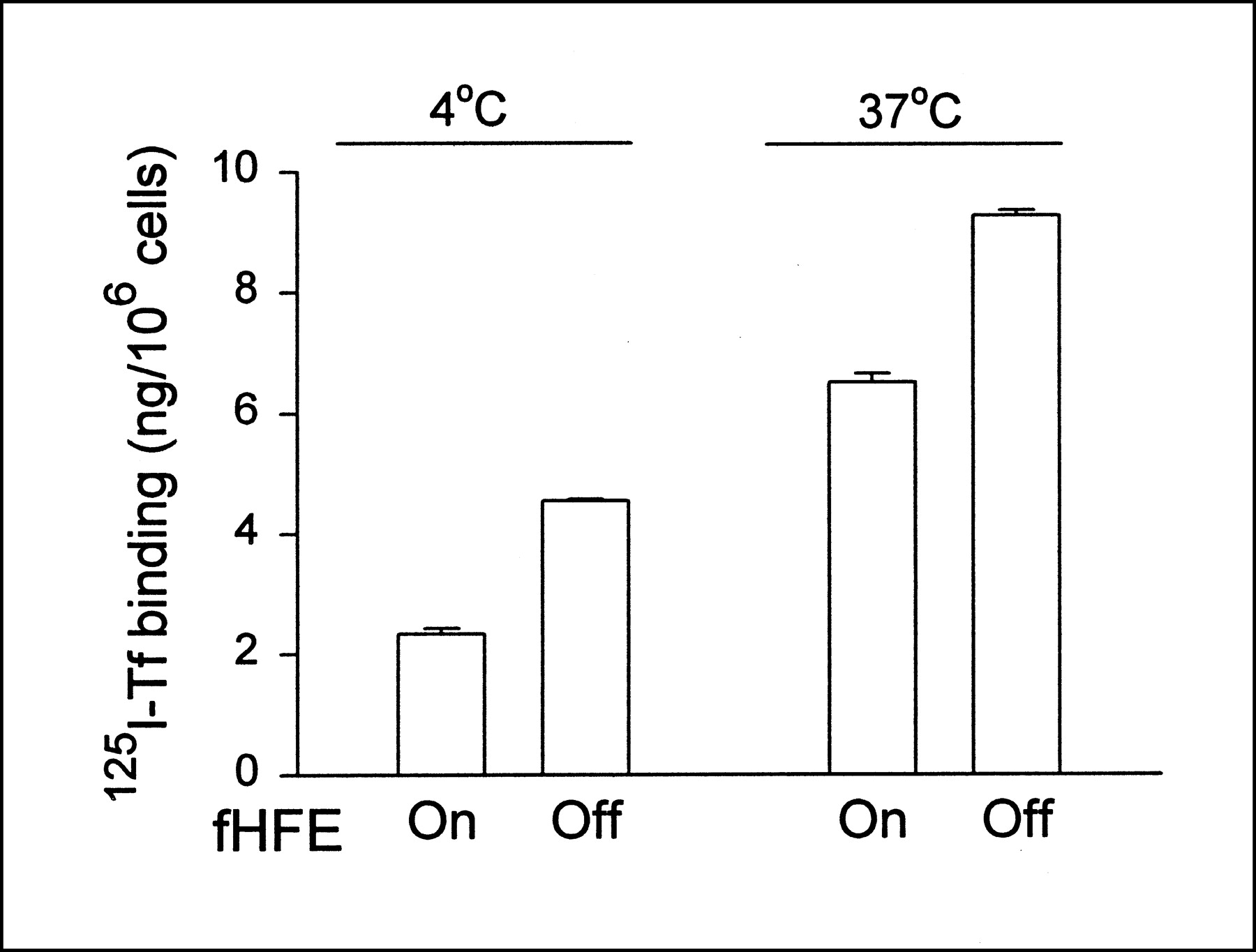
fHFE expression produces decrease in cellular Tf binding sites. Cells grown in presence (fHFE off) or absence (fHFE on) of doxycycline were analyzed by 125I-Tf binding assay at 4oC and 37°C as described. P < 0.003 for differences in Tf binding between cells with fHFE turned on and off.

Effect of fHFE expression on iron uptake by cells. Cells grown in presence of (fHFE off) or absence (fHFE on) of doxycycline were replated in fresh medium and analyzed for 59Fe uptake as described. Cells were incubated with 59Fe at 8 ng/mL (as 59Fe-Tf), and 59Fe uptake was measured after 24 and 48 h of incubation. Values shown represent means and range of representative experiment performed in duplicate. P < 0.00001 for differences in iron uptake between cells with fHFE turned on and off.
DISCUSSION
In this study, we show that an increase in the expression of the HFE gene significantly decreases the uptake of 67Ga by cells. Furthermore, our results suggest that the effect of HFE on 67Ga uptake is secondary to a reduction in the binding of Tf with its receptor rather than a reduction in the number of cellular TfRs. Analysis of TfR expression in fHFE-expressing cells by Western blotting revealed an increase in cellular immunoreactive TfR; however, an increase in TfRs per se would be expected to increase 67Ga-Tf uptake rather than decrease it. In contrast, the decrease in Tf binding sites, as determined by the 125I-Tf binding assay, would be expected to result in a decrease in the Tf-mediated uptake of 67Ga. Therefore, a key finding in our studies that explains the HFE-induced decrease in 67Ga uptake is that the expression of fHFE leads to a decrease in the number of Tf binding sites on the TfR in cells, which, in turn, leads to a decrease in the cellular uptake of 67Ga-Tf complexes. The HFE-induced reduction in available Tf binding sites is consistent with recent reports that show that HFE protein competes with Tf for binding to the TfR (16). Our studies also confirm earlier findings demonstrating similarities between 67Ga and iron uptake by cells via the Tf-TfR pathway.
Our study provides information that extends our current understanding of the mechanisms involved in 67Ga uptake and its use as a scanning agent for tumor imaging. First, whereas prior investigations have established a role for the TfR in 67Ga uptake, our results indicate that it is the availability of Tf binding sites rather than the level of the TfR protein per se that is a key factor in determining 67Ga uptake by cells. Hence, studies attempting to correlate TfR with 67Ga uptake may yield equivocal data if TfR levels are measured by immunologic methods alone using anti-TfR antibodies rather than by functional ligand binding assays. Second, the uptake of 67Ga is likely to be decreased in malignant cells that express significant levels of wild-type HFE protein relative to cells that do not express HFE. Although our studies were performed using the HeLa cell line, our results are likely to be applicable to the clinical situation where 67Ga scanning is used. Recently, we showed that immortalized peripheral blood B-lymphocyte cell lines derived from individuals with wild-type HFE or the HFE C282Y mutation express significant levels of wild-type or mutated HFE protein, respectively (17). It is likely, therefore, that HFE may be expressed in certain lymphoid malignancies and that this may impact on the results of 67Ga scanning frequently performed in these disorders.
Our results also support the idea that differences in 67Ga uptake by cells may exist between individuals with wild-type HFE and individuals with the HFE C282Y mutation. Whereas wild-type HFE protein binds to the TfR and interferes with Tf binding, the HFE C282Y protein does not bind to the TfR and thus does not interfere with the binding of Tf to its receptor (13). Accordingly, tumor cells with the HFE C282Y mutation would be expected to have greater 67Ga uptake than those expressing wild-type HFE. Although we did not specifically examine cells with the HFE C282Y mutation, cells in which fHFE gene expression was turned off could be considered to be analogous to cells expressing HFE C282Y since an HFE-TfR complex is not formed in either situation.
CONCLUSION
The expression of HFE and its interaction with the TfR may play a role in modifying the uptake of 67Ga by cells. Further studies are planned to examine the expression of HFE in lymphoma cells since 67Ga scanning is used for tumor imaging in this malignancy. Given the high frequency of the HFE C282Y mutation in the general population, such investigations appear warranted.
Acknowledgments
The authors are grateful to Caroline A. Enns, PhD, for generously providing the fwtHFE/tTA HeLa cell line for these studies. This work was supported by a grant from the Cancer Center of the Medical College of Wisconsin.
Footnotes
Received Jul. 25, 2002; revision accepted Jan. 21, 2003.
For correspondence or reprints contact: Christopher R. Chitambar, MD, Division of Neoplastic Diseases, Medical College of Wisconsin, 9200 W. Wisconsin Ave., Milwaukee, WI 53226.
E-mail: chitambr{at}mcw.edu
REFERENCES
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Gallium-induced cell death in lymphoma: role of transferrin receptor cycling, involvement of Bax and the mitochondria, and effects of proteasome inhibition
- Resistance to the Antineoplastic Agent Gallium Nitrate Results in Marked Alterations in Intracellular Iron and Gallium Trafficking: Identification of Novel Intermediates