


Abstract
Current noninvasive imaging methods for monitoring immune response were largely developed for interrogation of the local reaction. This study developed the radiotracer 64Cu-labeled anti-CD11b (64Cu-αCD11b) for longitudinal assessment of local and systemic immune response involving mobilization of CD11b+ myeloid cells by small-animal PET/CT. Methods: Acute or chronic inflammation in the ears of BALB/c mice was induced by 12-o-tetradecanoylphorbol-13-acetate. Acute lung inflammation was induced by intratracheal lipopolysaccharide inoculation. αCD11b was conjugated with p-SCN-Bn-DOTA followed by labeling with 64Cu. PET/CT and biodistribution were evaluated at different times after intravenous injection of 64Cu-αCD11b. Cell populations from bone marrow (BM) and spleen were analyzed by flow cytometry. Results: 64Cu-αCD11b was primarily taken up by BM and spleen in control mice. In comparison, 64Cu-αCD11b uptake was significantly reduced in the BM and spleen of CD11b-knockout mice, indicating that 64Cu-αCD11b selectively homed to CD11b+ myeloid cells in vivo. In mice with ear inflammation, for the local inflammatory response, 64Cu-αCD11b PET/CT revealed significantly higher 64Cu-αCD11b uptake in the inflamed ears in the acute inflammation phase than the chronic phase, consistent with markedly increased infiltration of CD11b+ cells into the inflammatory lesions at the acute phase. Moreover, imaging of 64Cu-αCD11b also showed the difference in mouse systemic response for different inflammatory stages. Compared with uptake in control mice, BM 64Cu-αCD11b uptake in mice with ear inflammation was significantly lower in the acute phase and higher in the chronic phase, reflecting an initial mobilization of CD11b+ cells from the BM to the inflammatory foci followed by a compensatory regeneration of CD11b+ myeloid cells in the BM. Similarly, in mice with lung inflammation, 64Cu-αCD11b PET/CT readily detected acute lung inflammation and recruitment of CD11b+ myeloid cells from the BM. Immunohistochemistry staining and flow cytometry results confirmed the noninvasive imaging of PET/CT. Conclusion: 64Cu-αCD11b PET/CT successfully tracked ear and pulmonary inflammation in mice and differentiated acute from chronic inflammation at the local and systemic levels. 64Cu-αCD11b PET/CT is a robust quantitative method for imaging of local and systemic immune responses.
When local inflammation develops, myeloid cells expand in the bone marrow (BM), enter the blood circulation, and travel to the inflamed sites. Some myeloid cells transit through secondary lymphoid organs (i.e., the spleen). Local diseases can also elicit a systemic immune response secondary to release of cytokines and chemokines from the disease sites. Yet other diseases are marked by systemic inflammation to begin with, as in the case of systemic autoimmune diseases. Because different types of tissue injury at different stages of disease progression elicit different local and systemic inflammatory responses, the ability to longitudinally interrogate the depletion, migration, and expansion of immune cells throughout the whole body will greatly facilitate disease characterization and guide selection of appropriate treatment regimens.
Several imaging techniques have been used to monitor immune responses, including intravital optical imaging, MRI, and nuclear imaging, but all these techniques have limitations (1,2). Intravital optical imaging can directly visualize the dynamics of interactions among different types of cells; however, this technique suffers from limited penetration depth (3). MRI is useful for monitoring trafficking of infused immune cells labeled ex vivo but is not yet suitable for imaging biomarkers in specific immune cell populations labeled in vivo. In the area of nuclear imaging, several radiotracers have been reported to permit monitoring of inflammation on the basis of changes in metabolism (4–6) or ex vivo labeling of inflammatory cells or inflammatory cytokines (7,8), but it is limited by labeling efficiency and the labor-intensive cell labeling procedures (9). To date, few studies reported visualization of both the local and the systemic immune responses to inflammatory and neoplastic diseases (10).
Most innate inflammatory cells, including granulocytes, monocytes, and macrophages, express CD11b (also known as αM-integrin or Mac-1) on their cell surface (11,12). We hypothesized that CD11b could be used as an imageable biomarker for noninvasive assessment of local and systemic immune responses. In this work, we report the application of small-animal PET/CT with 64Cu-labeled anti-CD11b (64Cu-αCD11b) in monitoring the mobilization of CD11b+ myeloid cells from the BM to secondary lymphoid organs and to local inflammatory lesions.
MATERIALS AND METHODS
Reagents
Rat antimouse CD11b antibody (αCD11b, clone M1/70, catalog number 14-0112), rat antimouse CD11b phycoerythrin (clone M1/70), rat antimouse Gr-1 PerCP-cyanine5.5 (clone RB6-8C5), and rat antimouse Ly6C allophycocyanin (clone HK1.4) were purchased from eBioscience Inc. p-SCN-Bn-DOTA (S-2-(4-isothiocyanatobenzyl)-1,4,7,10-tetraazacyclododecane tetraacetic acid) was purchased from Macrocyclics, Inc. 12-o-tetradecanoylphorbol-13-acetate (TPA) was purchased from Sigma-Aldrich. Lipopolysaccharide (Escherichia coli 0111) was purchased from List Biologics. 64CuCl2 and 18F-FDG were obtained from the Cyclotron Radiochemistry Facility of the University of Texas M.D. Anderson Cancer Center.
DOTA Conjugation and Radiolabeling
p-SCN-Bn-DOTA was added to αCD11b or isotype-matched IgG at p-SCN-Bn-DOTA–to–antibody molar ratios of 50:1 in 0.1 M sodium bicarbonate buffer (pH 8.5). The resulting DOTA-αCD11b conjugate was purified using a PD-10 column, and the DOTA number for DOTA-αCD11b was measured (Supplemental Table 1; supplemental materials are available at http://jnm.snmjournals.org). For radiolabeling, 64CuCl2 was diluted with sodium acetate buffer (pH 6.0), and the pH of the solution was adjusted to 6.0 with 1N NaOH. Each DOTA-antibody conjugate was then added into 64CuCl2 solution (10 μg of DOTA-antibody per 37 MBq of 64Cu) and incubated for 1 h at 38°C with constant shaking. 64Cu-αCD11b or 64Cu-IgG was purified by passage through a PD-10 column.
Number of DOTA per αCD11b Antibody
The average number of DOTA chelators per αCD11b antibody was measured according to reported procedures (13–15). Briefly, nonradioactive CuCl2 (80-fold excess of DOTA-αCD11b) in 20 μL of 0.1N sodium acetate buffer (pH 5.5) was added to approximately 37 MBq of 64CuCl2 in 50 μL of 0.1N sodium acetate buffer; then, 20 μg of DOTA-αCD11b in 40 μL of 0.1N sodium acetate buffer were added to the above carrier-added 64CuCl2 solution. The reaction mixture was incubated with constant shaking at 40°C for 1 h. The 64Cu-αCD11b was purified by a PD-10 column with ×1 phosphate-buffered saline, and eluent (3.0–4.5 mL) was collected. The number of DOTA per αCD11b antibody was calculated as moles (Cu2+) × activity (3.0–4.5 mL)/moles (DOTA-αCD11b)/total activity (loaded for each labeling). The activities in the equation were all decay-corrected to the same time point. The results were expressed as mean ± SD (n = 3).
Competitive Cell-Binding Assay
Freshly isolated BM cells from C57BL/6 mice were suspended in phosphate-buffered saline containing 0.5% bovine serum albumin (1 × 105 cells per 200 μL). Cells were incubated with 64Cu-αCD11b (3.7 kBq/well, ∼4 × 10−10 M) in the absence and presence of increasing concentrations of nonradioactive αCD11b at room temperature for 2 h with gentle shaking. After removal of culture medium under vacuum, cells were washed 3 times with phosphate-buffered saline containing 0.1% bovine serum albumin. Radioactivity of the cells from each well was counted with a γ-counter. The 50% inhibitory concentration of nonradioactive αCD11b was calculated by fitting the data with nonlinear regression using GraphPad Prism (GraphPad Software).
Animal Models
All animal experiments were performed in compliance with the guidelines for care and use of research animals established by the Institutional Animal Care and Use Committee of the University of Texas M.D. Anderson Cancer Center (IACUC protocol 00001333). Female BALB/c mice and C57BL/6 mice (6–8 wk of age) were obtained from Taconic. CD11b−/− (CD11b knockout) C57BL/6 mice (B6.129S4-Itgamtm1Myd/J, stock number 003991) were purchased from the Jackson Laboratory.
Ear Inflammation Model
Twenty microliters of TPA solution (100 μg/mL in acetone) were topically applied to the ventral and dorsal sides of the right ear of BALB/c mice 1, 4, or 11 times (8,16).
Lung Inflammation Model
BALB/c mice were anesthetized with 2% isoflurane, and then 20 μg of lipopolysaccharide in 0.1 mL of saline were given intratracheally. Control animals were given 0.1 mL of saline intratracheally (17).
PET/CT and Biodistribution
PET/CT was performed using an Inveon or Albira scanner after intravenous injection of 64Cu-αCD11b or its corresponding isotype antibody control 64Cu-IgG at a dose of 3.7 MBq/mouse (1.22 ± 0.22 μg antibody/3.7 MBq/mouse). The mice were euthanized 24 or 48 h after injection. Blood, major organs, and inflamed tissue specimens were collected, wet-weighed, and counted with a γ-counter (Packard). The results were reported as mean %ID/g and SD (n = 3/group).
PET Imaging Quantification
For each PET scan, 3-dimensional regions of interest were drawn over the major organs or inflammatory lesions on decay-corrected whole-body coronal images. The average radioactivity concentration (accumulation) within an organ or inflammatory lesion was obtained from mean pixel values within the region-of-interest volume and then converted to percentage injected dose per gram of tissue (%ID/g) or percentage injected dose per cubic centimeter of tissue (%ID/cc). Imaging data acquired from the Inveon scanner (Siemens) were analyzed using Inveon software (Siemens) and expressed as %ID/g. Imaging data acquired from the Albira scanner (Bruker) were analyzed using PMOD software (PMOD Technologies, Ltd.) and expressed as %ID/cc.
Autoradiography
Immediately after the PET/CT imaging, the ears were harvested and then taped to a chilled autoradiography cassette containing a Super Resolution screen (Packard) and stored overnight at 4°C. Screens were scanned with a Fujifilm FLA-5100 imaging system.
Hematoxylin–Eosin and Immunohistochemistry Staining
For hematoxylin–eosin staining, organ samples were fixed in 10% (v/v) buffered formalin and embedded in paraffin. Five-micrometer-thick sections were cut and stained with hematoxylin–eosin. For immunohistochemistry staining, the formalin-fixed paraffin sections were deparaffinized, rehydrated, and incubated in 3% aqueous H2O2 to block the endogenous peroxidase activity. After blocking with 2% normal horse serum, the primary antibody rat antimouse Ly6G (eBioscience Inc.) at 1:50 dilution was added and incubated at 4°C overnight. Immunohistochemistry reaction was performed using Vectastain Elite ABC HRP kit (rat IgG) (Vector Laboratories).
Flow Cytometry
BM cells were obtained from mouse femur according to reported procedures (18). For flow cytometry, fluorescence probes were added to single-cell suspensions prepared from samples obtained from BM or spleen, and cells were incubated on ice for 1 h. After washing with phosphate-buffered saline 3 times, cells were examined on a FACSCalibur flow cytometer or LSRFortessa X-20 analyzer (Becton Dickinson) and analyzed with FlowJo software (Tree Star, Inc.). The following antibodies were used: rat antimouse Ly6G-FITC, rat antimouse CD11b-phycoerythrin, rat antimouse Gr-1-PerCP-cyanine5.5, and antimouse Ly6C-allophycocyanin.
Statistics
Statistical significance was analyzed with IBM SPSS statistics 24 software using 1-way ANOVA. P values of less than 0.05 were considered statistically significant.
RESULTS
64Cu-αCD11b Selectively Binds to CD11b+ Myeloid Cells
The binding affinity between 64Cu-αCD11b and mouse BM cells was determined by a competitive displacement assay. The 50% inhibitory concentration between 64Cu-αCD11b and nonradioactive αCD11b was 1.56 × 10−10 ± 5.14 × 10−11 mol/L (Supplemental Fig. 1).
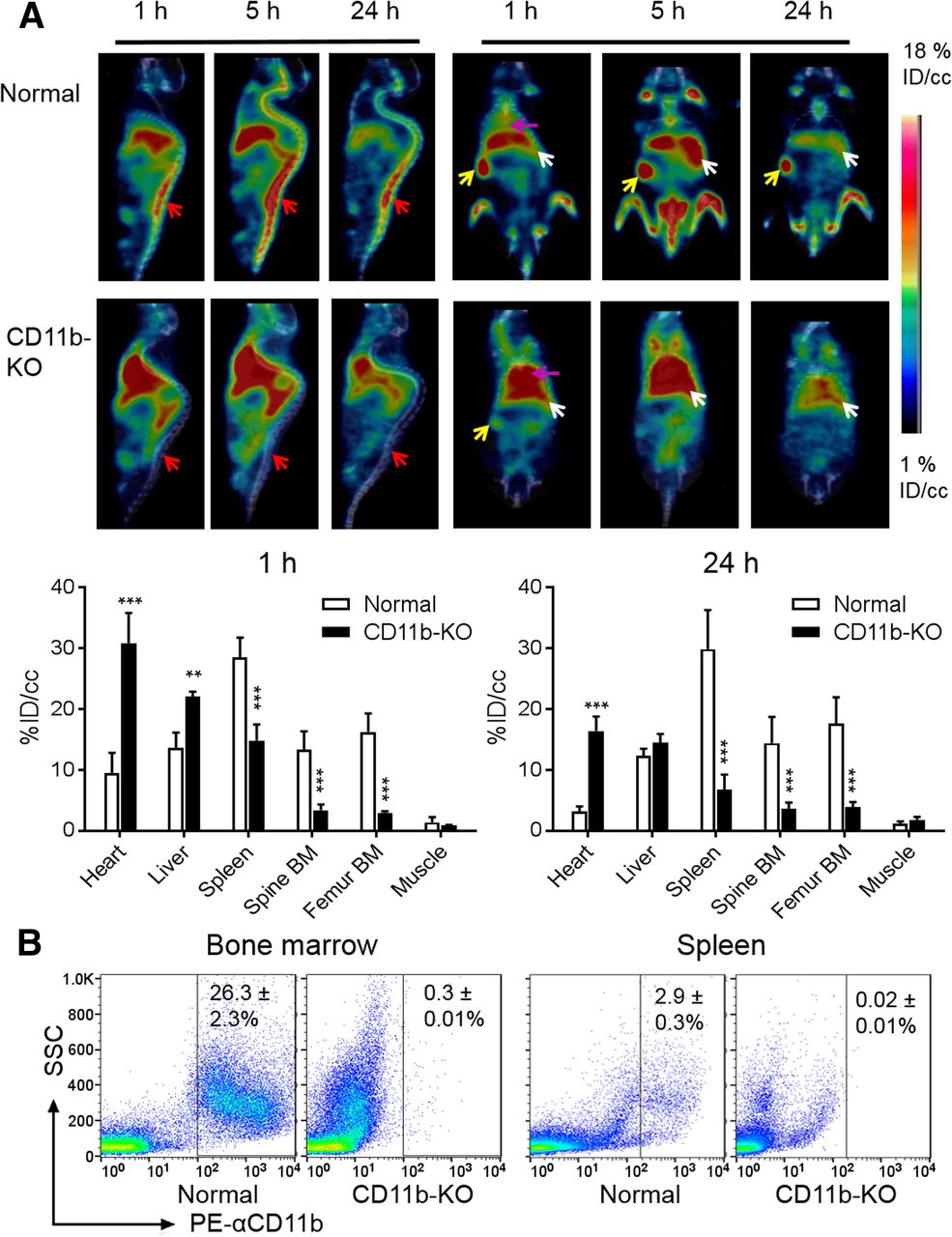
Figure 1A compares PET/CT images acquired at various times after intravenous injection of 64Cu-αCD11b in control (normal mice) (i.e., CD11b-sufficient) and CD11b knockout (CD11b−/−) C57BL/6 mice. In control mice, the radiotracer was distributed primarily to the BM, spleen, and liver. Strong signals from the spinal cord, hip bones, and joints could be visualized as early as 1 h after injection and peaked at 5 h after injection, suggesting robust uptake of 64Cu-αCD11b to the BM (which bears CD11b+ cells). The high uptake of 64Cu-αCD11b in the spleen is consistent with the fact that the mouse spleen harbors half the body’s monocytes within its red pulp (19). In contrast, in the CD11b knockout mice, the amount of 64Cu-αCD11b in the blood pool (heart) was significantly higher than that in control mice at all time points studied (P < 0.001). However, the uptake of 64Cu-αCD11b in the BM and spleen was significantly lower in the CD11b knockout mice than in the wild-type mice (Fig. 1A; Supplemental Figs. 2 and 3). Uptake of 64Cu-αCD11b in the liver was also greater in knockout mice, likely in part because of increased blood-pool activity. The observation that uptake in the heart and liver is nonspecific is also supported by the use of nonspecific IgG, as discussed below.

64Cu-αCD11b PET/CT in control (CD11b-sufficient) and CD11b-knockout C57BL/6 mice. (A) Representative PET/CT images of control mice (top) and CD11b-knockout (KO) mice (bottom). Red arrows: BM; yellow arrows: spleen; white arrows: liver; purple arrows: heart. PET quantification of 64Cu-αCD11b uptake is shown in major organs at 1 h and 24 h after intravenous injection of radiotracer. (B) Flow cytometry analysis of CD11b+ cells in BM and spleen of control and CD11b knockout (CD11b-KO) mice. All data are expressed as mean ± SD (n = 3/group). **P < 0.01. ***P < 0.001.
Flow cytometry analysis performed 48 h after radiotracer injection confirmed that the percentage of CD11b+ cells in the BM was 26.3% ± 3.0% in control mice versus 0.27% ± 0.01% in CD11b−/− mice (Fig. 1B; Supplemental Fig. 4). Likewise, the percentage of CD11b+ cells in the spleen was 2.9% ± 0.3% in control mice versus 0.02% ± 0.01% in CD11b−/− mice (Fig. 1B). These data indicate that 64Cu-αCD11b specifically bound to CD11b+ myeloid cells in vivo.
After peaking at 5 h, the uptake of 64Cu-αCD11b in the BM of control mice steadily and significantly decreased from 5 h to 24 h and 48 h after injection (P < 0.001), whereas uptake of 64Cu-αCD11b in the BM of knockout mice remained low at background levels, unchanged from 24 to 48 h (Supplemental Figs. 3C and 3D). This finding might be attributed to redistribution of 64Cu-αCD11b–bound myeloid cells from the BM to the peripheral circulation in control mice, clearance of radiotracer, or both.
In control BALB/C mice, 64Cu-αCD11b displayed a distribution pattern similar to that in control C57BL/6 mice: BM, spleen, and liver were the organs with major radiotracer uptake (Supplemental Figs. 5A and 5B). CD11b+ cells were the major cell population in the BM of BALB/c mice: 37.5% ± 1.92% of the BM cells expressed CD11b (Supplemental Fig. 5C).
64Cu-αCD11b PET Differentiates Different Stages of TPA-Induced Local Inflammation
We first investigated changes in the distribution pattern of CD11b+ myeloid cells at the local level in response to TPA-induced inflammation in the ears of BALB/c mice. PET/CT images acquired 24 h after intravenous injection of 64Cu-αCD11b showed significantly higher uptake in the inflamed ears 30 h after a single TPA challenge, during the acute inflammation stage (Fig. 2A). In comparison, when 4 or 11 TPA challenges were administered and imaging was done 8 or 22 d after the first challenge during the chronic inflammation stage, significantly less CD11b+ signal was seen within the inflamed ear. There was minimal radiotracer uptake in the contralateral, unchallenged ears or ears of control mice (Fig. 2A). These PET/CT data were confirmed by a classic biodistribution study (Fig. 2B).

64Cu-αCD11b PET/CT and biodistribution of 64Cu-αCD11b in BALB/c mice with TPA-induced acute and chronic ear inflammation. (A) Representative PET/CT images of control mouse, mouse with acute inflammatory focus after single TPA challenge in ear, and mice with chronic inflammatory foci after 4 and 11 TPA challenges. Images were acquired 24 h after intravenous injection of 64Cu-αCD11b. Purple arrows: inflamed ears; red arrows: BM; yellow arrows: spleen. (B) Biodistribution of 64Cu-αCD11b in control mice and mice with 1, 4, and 11 TPA challenges. Data were collected 48 h after intravenous injection of 64Cu-αCD11b. (C) Representative PET/CT images and corresponding quantitative analysis comparing 64Cu-IgG control antibody to 64Cu-αCD11b in mice with acute ear inflammation after single TPA challenge. Data are expressed as mean ± SD (n = 3/group). *P < 0.05. ***P < 0.001.
To exclude the possibility of nonspecific uptake of antibody in acute inflammation, we used 64Cu-IgG as a control. PET/CT showed low deposition of 64Cu-IgG in the BM and spleen (Fig. 2C), with most of the signal being detected in the heart and liver, a distribution pattern typical of nonspecific antibodies (20). Quantitative analysis showed that 64Cu-αCD11b uptake in the ears with acute inflammation was 3.8 times that of 64Cu-IgG (P < 0.001), further supporting that 64Cu-αCD11b selectively bound to CD11b+ myeloid cells in acute inflammatory lesions in vivo. Immunohistochemistry staining of Ly6G, a marker for myeloid-derived granulocytes and neutrophils (CD11b+Ly6G+) (21), showed infiltration of the inflamed ears by a large number of Ly6G+ neutrophils 30 h after a single TPA challenge (Fig. 3A). Neutrophils are among the first inflammatory cells to migrate toward a site of inflammation. In contrast, multiple applications of TPA produced a chronic inflammatory reaction characterized by increased ear thickness (Supplemental Fig. 6) and diminished Ly6G+ neutrophils (Fig. 3A).
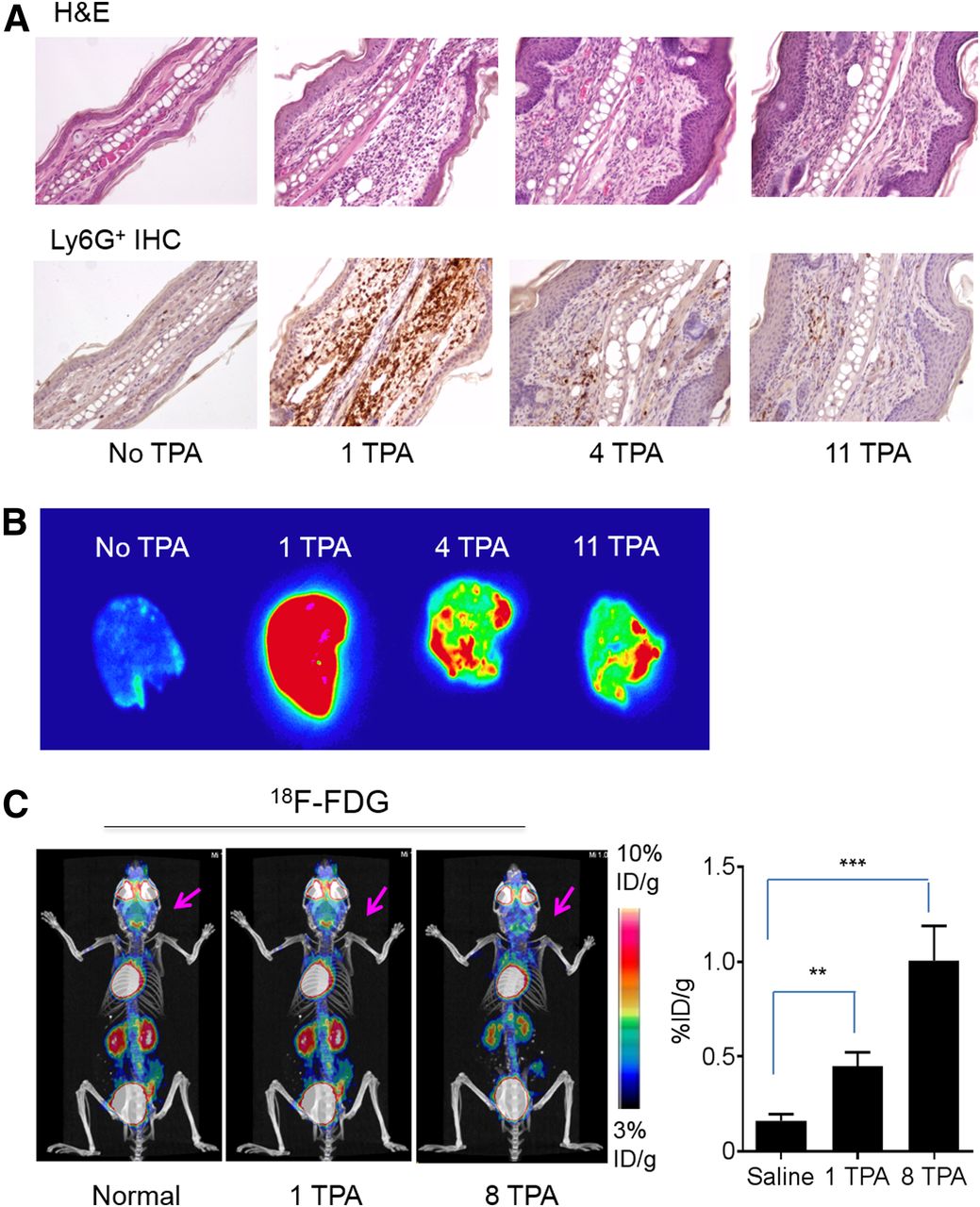
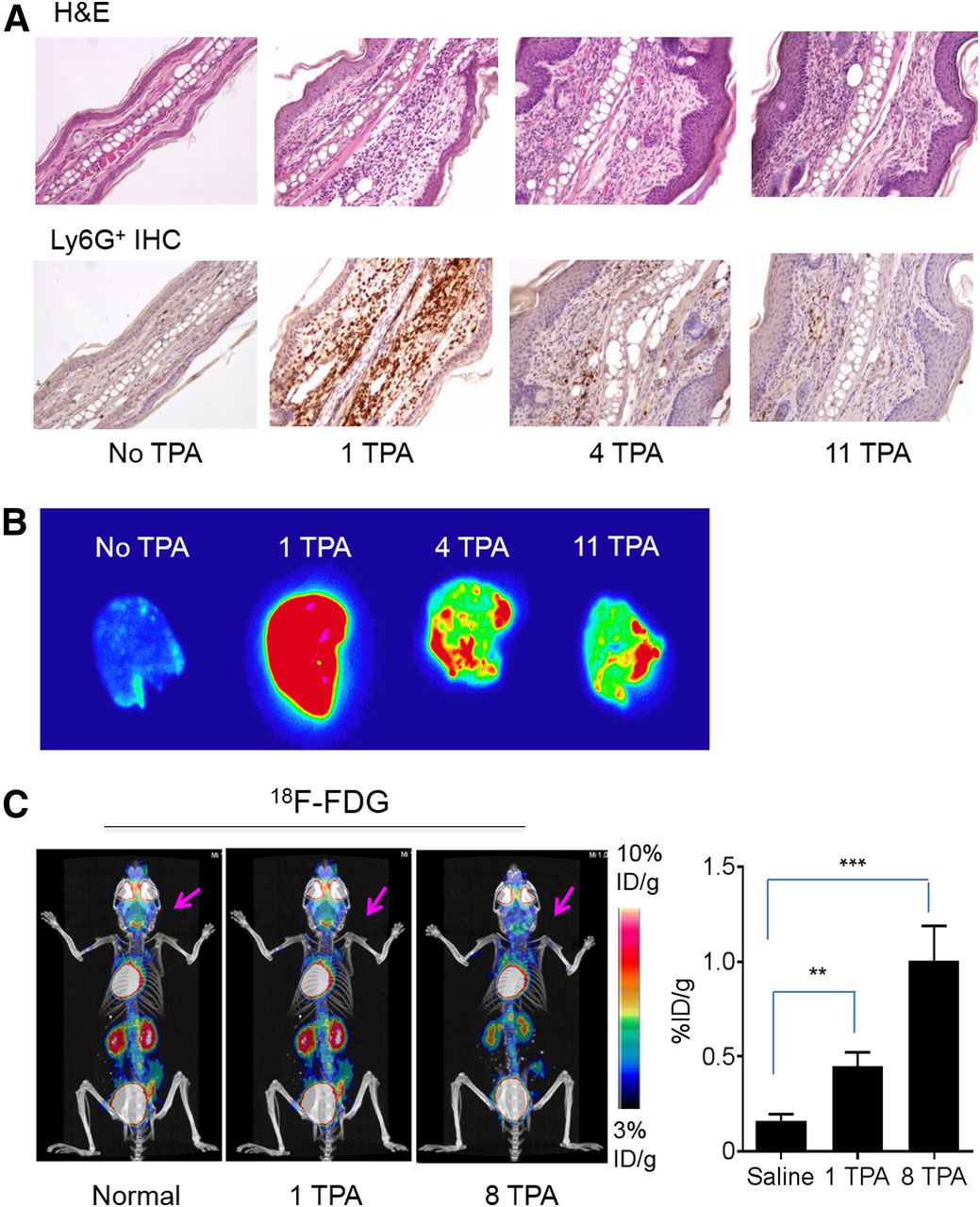
Histologic and immunohistochemistry confirmation of local inflammatory response in TPA-treated ears. (A) Representative images of sections of hematoxylin- and eosin-stained (H&E) and immunohistochemistry-stained (IHC) inflamed ears (×200). (B) Autoradiographs of ears from mice with different TPA treatments removed 48 h after intravenous injection of 64Cu-αCD11b. (C) Representative 18F-FDG PET/CT image of control mouse and mice with 1 and 8 TPA challenges in ears. Arrows: inflamed ears. corresponding quantitative analysis of 18F-FDG uptake in ears are shown. Data are expressed as mean ± SD (n = 3/group). **P < 0.01. ***P < 0.001.
Autoradiography 48 h after intravenous injection of 64Cu-αCD11b showed minimal radioactivity in normal ears (Fig. 3B), and intense radioactivity in ears after a single TPA challenge, consistent with extensive neutrophil infiltration, but decreased radioactivity in ears with 4 or 11 TPA challenges, consistent with decreased neutrophil infiltration in ears with chronic inflammation.
PET with 18F-FDG, which tracks elevated glucose metabolism, is a clinically established technique to assess the metabolic activity of inflammatory processes. In both acute and chronic inflammation in BALB/c mice, the inflamed ears were barely visible with 18F-FDG PET/CT (Fig. 3C; Supplemental Fig. 7). Nevertheless, the uptake of 18F-FDG in the ears after a single TPA challenge and after 8 TPA challenges was significantly higher than the uptake in normal ears, indicating low but increased metabolic activity in the inflamed ears. On comparison, the uptake of 18F-FDG in the inflamed ears (0.45–1.00 %ID/g) was much lower than the uptake of 64Cu-αCD11b in the ears with either acute inflammation (38.7 %ID/g) or chronic inflammation (3.6–6.9 %ID/g). Taken together, these data indicate that 64Cu-αCD11b PET/CT is more effective in differentiating acute from chronic inflammation than is tracking changes in metabolic activity with 18F-FDG PET/CT.
PET with 64Cu-αCD11b Detects TPA-Induced Inflammation at Systemic Level
In mice with ear inflammation, 64Cu-αCD11b PET/CT revealed a small decrease in the uptake of 64Cu-αCD11b in the BM after a single TPA challenge. Interestingly, BM uptake of 64Cu-αCD11b was significantly higher after 4 and 11 TPA challenges than after a single TPA injection (Fig. 2A). 64Cu-αCD11b uptake in the spleen increased after a single TPA challenge and further increased after additional TPA challenges (Fig. 2A; Supplemental Fig. 8). Uptake of radiotracer in the bone (including BM) of control mice and mice with 1, 4, and 11 TPA challenges was 13.1 ± 0.25, 10.6 ± 1.34, 15.9 ± 1.55, and 16.3 ± 0.69 %ID/g (P < 0.05 for all comparisons with control mice), respectively (Fig. 2B). These data likely underestimated the uptake values in the BM because the weight of the cortical bone was included in the calculation of ID%/g values. Thus, changes in CD11b+ myeloid cells in the BM and spleen could be readily visualized using 64Cu-αCD11b PET/CT.
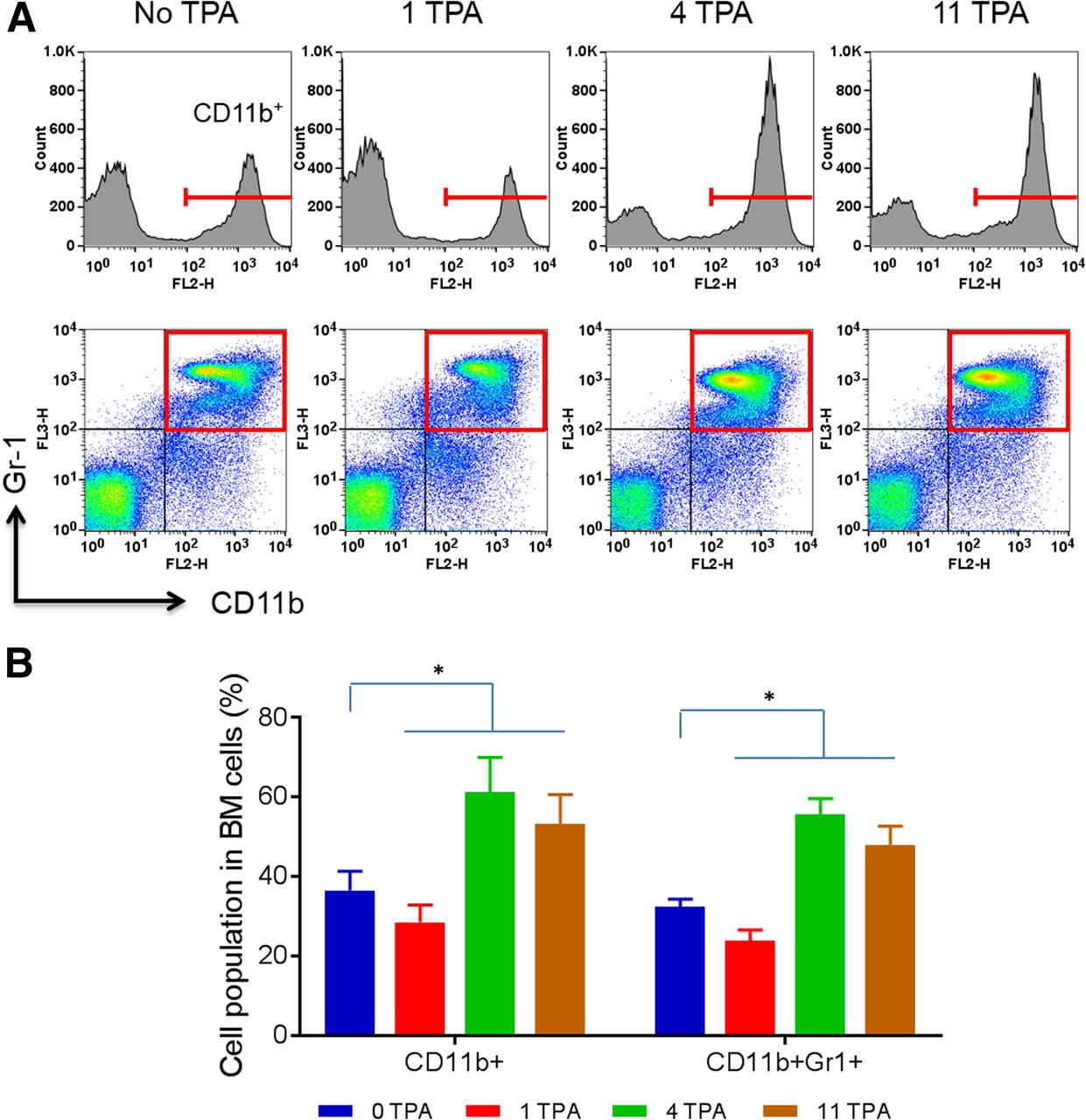
Flow cytometry analysis showed that CD11b+ cells in the BM cells of control mice and mice with 1, 4, and 11 TPA challenges were 36.5% ± 4.8%, 28.5% ± 4.4%, 61.2% ± 8.8%, and 53.2% ± 7.5%, respectively (Figs. 4A and 4B). Furthermore, the CD11b+Gr-1+ double-positive cells, consisting of myeloid progenitors and immature granulocytes, macrophages, and dendritic cells, in the BM cells of control mice and mice with 1, 4, and 11 TPA challenges were 32.3% ± 2.0%, 23.9% ± 2.7%, 55.5% ± 4.1%, and 47.9% ± 4.7%, respectively (Figs. 4A and 4B). These findings were consistent with the 64Cu-αCD11b PET/CT data that revealed an initial depletion of CD11b+ cells in acute inflammation followed by an expansion of CD11b+ cells in chronic inflammation in the BM (Fig. 2A).

Flow cytometry analysis of BM cells from mice with TPA-induced ear inflammation. (A) CD11b+ cells and CD11b+Gr-1+ double-positive cells from BM in mice with inflamed ears after 1, 4, and 11 TPA challenges. (B) Flow cytometric quantitation of percentages of CD11b+ cells and CD11b+Gr-1+ cells in whole BM cells in control mice (0 TPA) and mice with 1, 4, and 11 TPA challenges. All data are expressed as mean ± SD (n = 3/group). *P < 0.05.
PET with 64Cu-αCD11b Detects Endotoxin-Induced Lung Inflammation at Local and Systemic Levels
Lung inflammation induced by intratracheal lipopolysaccharide closely reflects human lung inflammation (22). PET/CT 24 h after intravenous injection of 64Cu-αCD11b (28 h after intratracheal injection of lipopolysaccharide) revealed higher radiotracer accumulation in the lungs, spleen, and liver and lower radiotracer signal in the BM of mice with lipopolysaccharide treatment (Fig. 5A). The lipopolysaccharide-treated lungs showed an increased thickness of the alveolar walls and increased neutrophil infiltration (Fig. 5B). There was significantly higher uptake of 64Cu-αCD11b in the lungs of lipopolysaccharide-treated mice (6.23 ± 1.57 %ID/cc) than in the lungs of control mice (2.03 ± 0.09 %ID/cc, P < 0.01) 24 h after radiotracer injection (Fig. 5C). Lipopolysaccharide-treated mice exhibited significantly lower radiotracer uptake in the spine (8.8 ± 1.2 %ID/cc vs 12.9 ± 1.2 %ID/cc, P < 0.05) and femoral BM (7.4 ± 0.7 %ID/cc vs 13.8 ± 2.1, P < 0.01) than did control mice. These lipopolysaccharide-treated mice also showed significantly higher radiotracer uptake in the spleen (24.3 ± 0.4 %ID/cc vs 16.4 ± 1.5 %ID/cc; P < 0.001), as well as the liver (19.2 ± 0.7 %ID/cc vs 10.4 ± 0.7 %ID/cc; P < 0.001). These CD11b+ cellular distribution patterns were similar to those observed in mice with TPA-induced acute ear inflammation.

PET/CT of 64Cu-αCD11b in BALB/c mice with lipopolysaccharide-induced acute lung inflammation. (A) Representative PET/CT images of control mouse with intratracheal instillation of saline and mouse with acute inflammation in lung induced by intratracheal instillation of lipopolysaccharide. Images were acquired 24 h after intravenous injection of 64Cu-αCD11b and 28 h after lipopolysaccharide instillation. Purple arrows: inflamed lung; red arrow: BM; yellow arrow: spleen. MIP = maximum intensity projections; 2D = 2-dimensional. (B) Representative hematoxylin- and eosin-stained lung tissues. Green arrows: granular leukocytes. (C) PET imaging quantification of 64Cu-αCD11b in control mice and mice with lung inflammation at 24 h. (D) Flow cytometry analysis of BM cells from control mice and mice with acute lung inflammation performed at 24 h. Data are expressed as mean ± SD (n = 3/group). *P < 0.05. **P < 0.01. ***P < 0.001.
Flow cytometry data revealed that the population of CD11b+ cells in the BM of lipopolysaccharide-treated mice decreased significantly compared with that of the control mice (Fig. 5D; Supplemental Fig. 9A), consistent with the 64Cu-αCD11b PET imaging data. CD11b+Gr-1+ cells consist of 2 major subsets: Ly6G+Ly6Clow granulocytic cells and Ly6G-Ly6C+ monocytic cells. Results showed a significant decrease in Ly6G+Ly6Clow granulocytic cells but not Ly6G-Ly6C+ monocytic cells in the BM of lipopolysaccharide-treated mice compared with values in control mice (Fig. 5D; Supplemental Fig. 9B). These results also demonstrated that 64Cu-αCD11b PET/CT permitted visualization of the dynamic innate immune response at both the local inflammation site and at the whole-body level, after a localized immune challenge.
DISCUSSION
In this study, we found that 64Cu-αCD11b specifically and robustly labeled CD11b+ myeloid cells in vitro and in vivo. 64Cu-αCD11b PET/CT could track and quantify changes in the number of CD11b+ cells both at the local level at the inflammatory sites and at the systemic level in the BM and the secondary lymphoid organ (spleen).
Several lines of evidence suggest that 64Cu-αCD11b specifically bound to CD11b+ cells in vivo. First, whereas the CD11b-sufficient C57BL/6 mice exhibited high radiotracer uptake in the BM and spleen, which are reservoir organs for CD11b+ cells, the CD11b-knockout mice had significantly lower uptake of the radiotracer in these organs and a significantly higher amount of 64Cu-αCD11b in the blood pool (particularly the heart). Second, deposition of nonspecific 64Cu-IgG in the BM and spleen of ear inflammation mice was significantly lower than the deposition of 64Cu-αCD11b in these organs.
Our study showed that 64Cu-αCD11b PET/CT could be used to differentiate acute from chronic local inflammation. 64Cu-αCD11b PET/CT clearly delineated inflamed ears in the acute inflammation phase and reduced radiotracer uptake in the chronic inflammation phase. Immunohistochemistry staining confirmed the presence of a large number of neutrophil granulocytes (CD11b+Ly6G+) within the inflammatory lesions in the acute phase but only a small number of CD11b+Ly6G+ granulocytes infiltrating the lesions in the chronic phase. It is known that during chronic inflammation, other types of immune cells responsible for tissue repair and remodeling, such as lymphocytes and fibroblasts that do not express CD11b, are the dominant inflammatory cells (23).
Importantly, our studies suggest that 64Cu-αCD11b PET/CT may be used to noninvasively interrogate mobilization of innate immune cells from the BM through the spleen and liver compartments to the diseased sites. In mouse inflammation models, 64Cu-αCD11b PET/CT demonstrated a decrease in BM radiotracer uptake in the acute inflammation phase and a significant increase in BM radiotracer uptake in the chronic inflammation phases. These imaging data were consistent with reduced CD11b+ and CD11b+Ly6G+ cells in the BM in the acute inflammation phase and increased CD11b+ and CD11b+Ly6G+ cells in the BM during chronic inflammation. Myeloid cells, in particular neutrophils, provide an important early defense mechanism against acute infection and inflammation. Under normal circumstances, most of the neutrophil pool is stored in the BM; only a small fraction of neutrophils enters the blood circulation at any given time (<2% of 65 million in the mouse) (24). Thus, the immediate need for a large number of neutrophils in the case of acute inflammation cannot be met by circulating neutrophils in the blood pool, leading to rapid mobilization of neutrophils from the BM. This dynamic but transient process of recruiting CD11b+ myeloid cells and CD11b+Ly6G+ granulocytes from the BM and their trapping in the spleen and liver were readily detected and quantified using 64Cu-αCD11b PET/CT. In the chronic inflammation phase, the system may be signaled to produce more myeloid cells, though this possibility needs verification using BrdU labeling studies. Likewise, extramedullary myelopoietic activity in the spleen may have contributed to the increased uptake of 64Cu-αCD11b in the spleen, though this hypothesis needs further verification. To sum, 64Cu-αCD11b PET/CT represents a powerful tool for studying the dynamics of the innate immune response locally and systemically in living animals.
Our finding that CD11b+ myeloid cells in the BM were readily accessible to 64Cu-αCD11b contrasts with the findings reported by Rashidian et al. (25), who used a much smaller variable domain of a camelid heavy-chain–only antibody targeting CD11b (∼15 kDa), 18F-VHHDC13. Although this 18F-labeled probe showed uptake in spleen and lymph nodes, there was no uptake in the BM. It may be that the αCD11b used in our study and VHHDC13 bind to different CD11b epitopes, which may be differentially expressed in BM-derived myeloid cells and in tumor-associated myeloid cells. It is also possible that the much larger αCD11b (∼150 kDa) has a longer blood half-life than that of VHHDC13 (∼20 min) and thus a greater likelihood of delivery to the cells in the BM.
Our study has a few limitations. First, CD11b is expressed on several different types of myeloid cells and on a few nonmyeloid cells as well. Additional imaging markers are needed to enhance cellular resolution. Second, the systemwide myeloid cell trafficking studies need to be expanded with accurate enumeration of CD11b cell numbers in various primary and secondary immune compartments, fortified with BrdU labeling studies. Third, whereas we have established that 64Cu-αCD11b could image CD11b in murine models of disease, the clinical significance of this approach in comparison to other imaging methods currently used to assess inflammation remains to be determined.
CONCLUSION
PET/CT imaging with 64Cu-αCD11b has the potential for tracking and quantifying changes in CD11b+ cell numbers longitudinally and noninvasively both locally and systemically.
DISCLOSURE
This work was supported by the John S. Dunn Foundation and by National Institutes of Health/National Cancer Institute Cancer Center Support Grant P30CA016672. No other potential conflict of interest relevant to this article was reported.
KEY POINTS
QUESTION: Can we use PET/CT for longitudinal assessment of local and systemic immune response involving mobilization of CD11b+ myeloid cells?
PERTINENT FINDINGS: PET/CT with 64Cu-labeled anti-CD11b antibody (64Cu-αCD11b) successfully tracked and differentiated acute and chronic inflammation at local and systemic levels. 64Cu-αCD11b small-animal PET/CT is a robust quantitative method for imaging systemic immune responses.
IMPLICATIONS FOR PATIENT CARE: 64Cu-αCD11b PET/CT may be useful for noninvasive assessment of a number of diseases involving abnormal activity of CD11b+ myeloid cells, including infectious disease, autoimmune disease, and toxicity and responses to antitumor therapies.
Acknowledgments
We thank Stephanie Deming for editing this article and Daniel Young for help with preparing samples for histologic examinations. We thank the Flow Cytometry and Cellular Imaging Core Facility and Small Animal Imaging Facility at M.D. Anderson Cancer Center for their support.
Footnotes
Published online Feb. 22, 2019.
- © 2019 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication September 12, 2018.
- Accepted for publication January 23, 2019.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}