


Abstract
The 2006 National Academy of Sciences Biologic Effects of Ionizing Radiation (BEIR) VII report is a well-recognized and frequently cited source on the legitimacy of the linear no-threshold (LNT) model—a model entailing a linear and causal relationship between ionizing radiation and human cancer risk. Linearity means that all radiation causes cancer and explicitly excludes a threshold below which radiogenic cancer risk disappears. However, the BEIR VII committee has erred in the interpretation of its selected literature; specifically, the in vitro data quoted fail to support LNT. Moreover, in vitro data cannot be considered as definitive proof of cancer development in intact organisms. This review is presented to stimulate a critical reevaluation by a BEIR VIII committee to reassess the validity, and use, of LNT and its derived policies.
See an invited perspective on this article on page 1014.
The National Academy of Sciences Biologic Effects of Ionizing Radiation (BEIR) VII report (1) concludes that the available human epidemiologic studies and animal experiments lack statistical power to distinguish a linear no-threshold (LNT) model from the absence of an effect at lower doses. Although none of the numerous publications cited in BEIR VII have demonstrated increased cancer risk from low-dose radiation exposure and thus none provide evidence for LNT, BEIR VII nevertheless asserts that the current scientific evidence is consistent with a linearly proportional relationship between cancer risk and low-dose ionizing radiation and that there is no threshold below which an increased cancer risk does not occur.
On page 245 of BEIR VII, the report asserts,It is abundantly clear that direct epidemiologic and animal approaches to low-dose cancer risk are intrinsically limited in their capacity to define possible curvilinearity or dose thresholds for risk in the range 0–100 mSv. For this reason the present report has placed much emphasis on the mechanistic data that can underpin such judgments.
BEIR VII’s mechanistic view subscribes to a model of radiation carcinogenesis consisting of a series of events in intact organisms—from radiation exposure to different types of DNA damage followed by chromosomal damage and (incomplete) repair to clinical cancer—leading inevitably to clinical cancer. Since BEIR VII notes that epidemiologic data linking low-dose radiation to clinical cancer are “intrinsically limited,” much emphasis is placed on a single study concerning one “mechanistic” feature of cancer initiation, that of radiogenic chromosomal damage. That study (2), performed in vitro, was selected to better define the shape of the dose–response curve in the low-dose range of 0–100 mSv and is the focus of the current review.
Also on page 245 the report says,…given the intimate relationship established between DNA damage response, gene or chromosomal mutations, and cancer development, the form of the dose-response for mutation induction in single cells should be broadly informative for cancer initiation. Data from a large-scale study noted in Chapter 2 suggest a linear relationship between low-[linear energy transfer] dose and chromosomal mutation down to around 20 mGy.
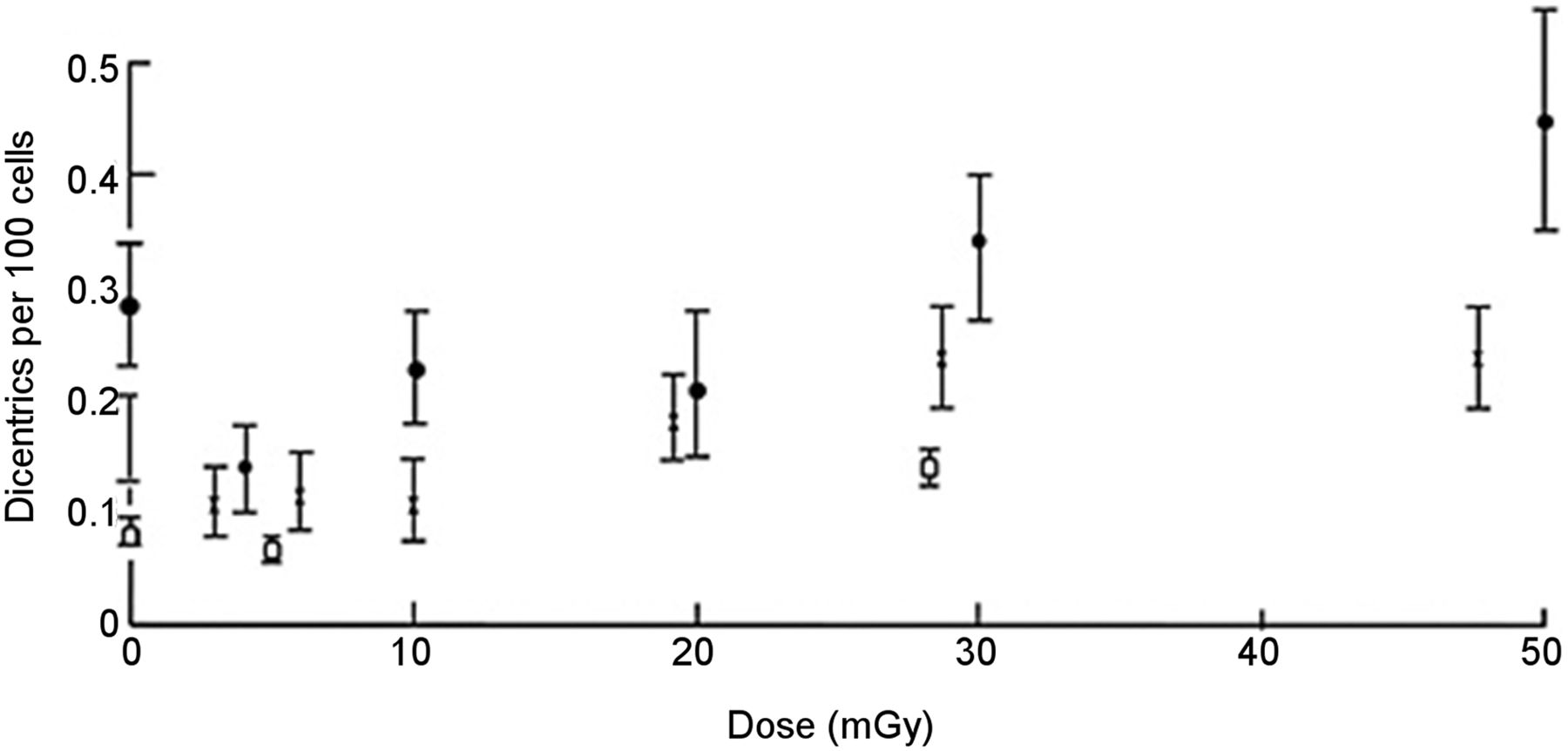
The large-scale study is by Lloyd et al. (2) and is the basis of BEIR VII’s assertion:Considerable effort has been expended to test this proposition [i.e., that the dose-response for all forms of mutations will be linear at low doses and low dose rates], and in a very large multicenter study using assays of dicentric aberrations [one biomarker of radiation damage] in human lymphocytes, the linearity of the response was evident down to at least 20 mGy of low-[linear energy transfer] radiation (Lloyd and others 1992), which is illustrated in Figure 2-5 [reproduced as Fig. 1

Lloyd’s study is a collaboration among 6 laboratories that counted chromosomal aberrations in human lymphocytes induced in vitro by x-rays over a 0- to 300-mGy dose range. Figure 1 is an exact replication of BEIR VII’s Figure 2-5, which reproduces a figure from the Lloyd article (2). Figure 1 represents 3 datasets from separate experiments, two by Lloyd and one by Pohl-Rüling et al. (3). For better clarity, we have redrawn Figure 1 as a bar graph, with a nonlinear x-axis, in Figure 2, showing only mean values of dicentrics at each nominal dose level and Lloyd’s zero-dose data point of 0.17 for experiment 1, which was omitted from BEIR’s Figure 2-5.
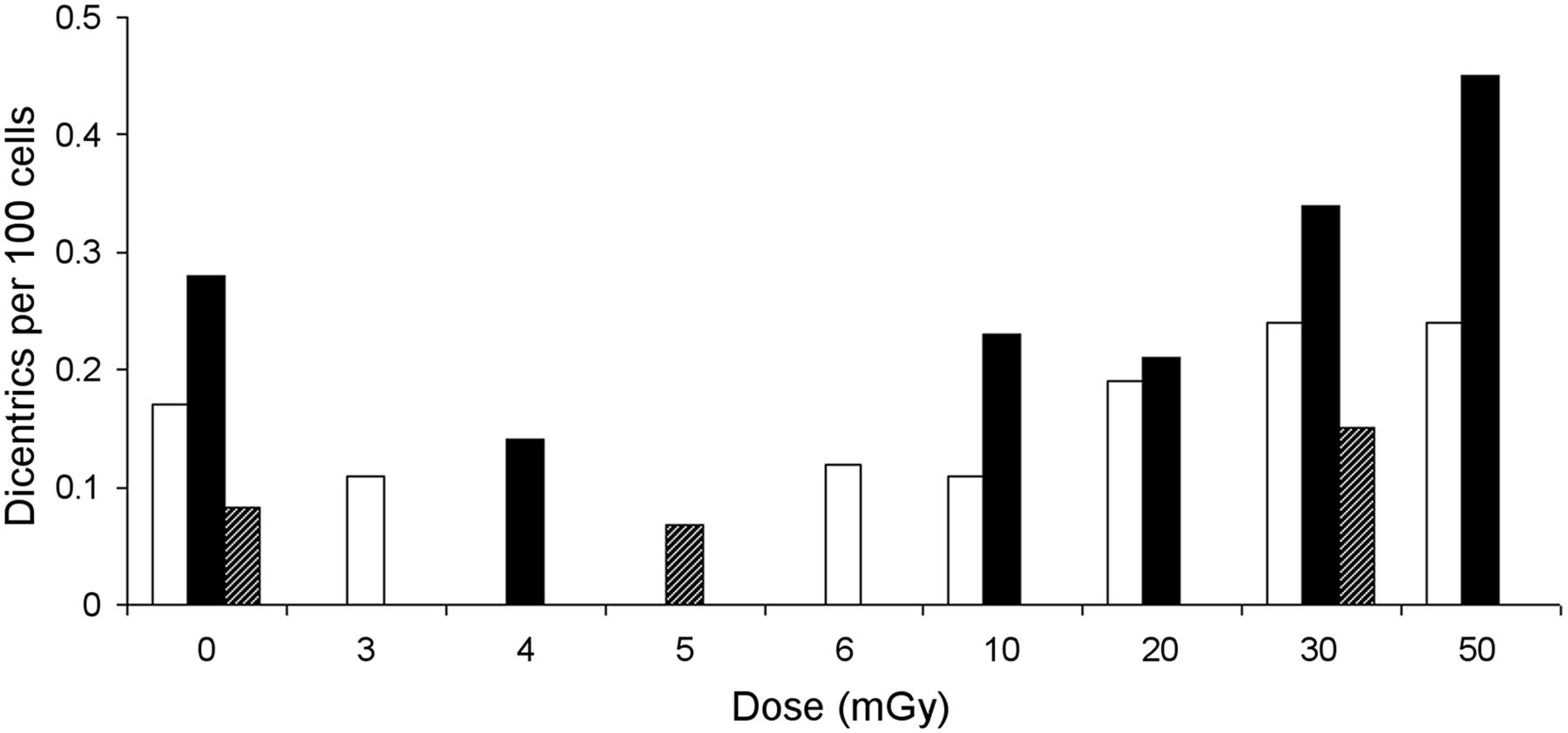
CRITIQUE OF USE OF IN VITRO MECHANISTIC DATA
Specifically, in experiment 1, Lloyd et al. found the numbers of dicentrics per 100 cells to be 0.17, 0.11, 0.12, 0.11, 0.19, 0.24, 0.24, and 1.28, for nominal doses of 0, 3, 6, 10, 20, 30, 50, and 300 mGy, respectively (Figs. 1 and 2). Relative to the control (zero-dose) value, Lloyd found actual reductions in dicentrics at the nominal doses of 3, 6, and 10 mGy, followed by small increases at the higher doses of 20, 30, 50, and a significant increase only at the much higher dose of 300 mGy. In Lloyd’s experiment 2, fewer doses were used, but the data support this initial reduction in dicentrics from the control value. Nevertheless, they dismiss the values below 20 mGy as subject to too much uncertainty based on the small numbers of damaged dicentrics, saying that the apparent reduction at the 3 lowest nonzero doses most likely, “is not of biological origin but may be an artefact of the higher-than-expected frequency of spontaneously arising [zero-dose] aberrations.”
Lloyd et al. rejected the “higher-than-expected” zero-dose value as valid, interpreting it as statistical noise, and because of statistical variations below 20 mGy, concluded that, “the question of whether linearity extends down to zero dose, or whether there is a plateau (threshold) indicative of inducible repair remains open.” This served as justification for their assertion that, “there is nothing in the present work which is incompatible with the prudent radiological protection assumption of a linear initial slope.”
In fact, the data are not compatible with a “linear initial slope.” There is no initial linear rise; rather, there is no measurable increased dose response from 3 to 10 mGy and all these initial values are no greater than the zero-dose value. Above 10 mGy, there is a less-than-proportional (i.e., nonlinear) rise and possibly a second plateau from 20 to 50 mGy (Figs. 1 and 2), in the range where Lloyd regards the data as less uncertain and more reliable. These data, in contrast, appear more supportive of a threshold. Lloyd also cites a previous, similar in vitro study by Pohl-Rüling et al. (3), who found a similar initial negative slope but, rather than dismissing the finding as noise, offered a biologic explanation (see below). Pohl-Rüling’s data are also represented in BEIR VII’s Figure 2-5, albeit without mention in the report.
With respect to the 20- to 300-mGy data points in experiment 1, Lloyd concludes that, “This work has shown conclusively that linearity of biological effect can be demonstrated with this system down to doses of about 20 mGy.” However, this is not borne out by their data. The inclusion of the non–low-dose (300 mGy) value dominates the increase in dicentrics above 20 mGy, and the departure from linearity at 30 and 50 mGy was implicitly regarded as measurement error, although Lloyd downplays error in this range. Not only are the 20- to 50-mGy values relatively flat, but they hardly exceed the empiric control value (0.17) at 0 mGy.
Furthermore, Lloyd’s data are consistent with a more recent observation from the Life Span Study atomic bomb survivor population (4):Linearity at low doses does not exist [in this case 20–50 mGy and likely beyond, somewhere between 50 and 300 mGy]; rather, it is forced by the high-dose extrapolation of the LNT model.
In contrast to Lloyd, Pohl-Rüling does not discount the initial reduction in the dicentrics below the control value (at doses of 4, 10, and even up to 20 mGy; Figs. 1 and 2). In agreement with many other experimenters who have found the same feature (5,6), Pohl-Rüling acknowledges the reductions as being real and offers the following biologic explanation:The deviations can be explained by an altered kinetics of aberration production at very low doses probably due to DNA repair mechanisms operating in these cells.
DISCUSSION
BEIR VII, first, claims “a linear relationship between low-linear energy transfer dose and chromosomal mutation down to around 20 mGy”—an assessment not established by Lloyd’s data—and, second, ignores the fact that Lloyd’s data were obtained only in vitro. Such studies can only suggest mechanisms of cancer initiation; they cannot by themselves provide evidence of clinical cancer development in whole organisms. In fact, on page 133 of Chapter 5, the BEIR report asserts that “at relatively low doses, there is still uncertainty as to whether there is an association between radiation and disease, and if there is an association, there is uncertainty about whether it is causal or not.”
Thus, BEIR’s assertion that the link between initiation and clinical cancer has been “established” is unsupportable. The continual creation from normal mitochondrial metabolism, in both intact organisms and cells in culture, of DNA-damaging reactive oxygen species results in frequent production of genetic mutations, but clinical cancer development in intact organisms is usually prevented by evolved adaptive protective responses, from cellular (described by Pohl-Rüling) to tissue to organismal levels. These mechanisms include antioxidant production, bystander signaling among damaged cells and their neighbors, and immune system cleanup of unrepaired cells (5–7).
Although DNA damage (i.e., genetic mutations) may be a necessary prequel, it is far from sufficient to produce clinical cancer (8,9). The outdated “one mutation = one cancer” model has been replaced by immune-system failure (4). Subsequent repair or removal of DNA double-strand breaks back to background levels have been demonstrated in patients within 24 h after a CT scan (10).
BEIR VII’s assertion—that “the presence of a true dose threshold demands totally error-free” repair in response to radiogenic damage—ignores removal responses. And their own selected data from Lloyd (and incidentally the data of Pohl-Rüling) negate the essential conclusion of their report that “current scientific evidence is consistent with the hypothesis that there is a linear, no-threshold dose–response relationship between exposure to ionizing radiation and the development of radiation-induced solid cancers in humans.” Specifically, the Lloyd study negates LNT on both counts; it fails to show either linearity from zero-dose or absence of a threshold. This serves to confirm the significant evidence that our bodies do not respond similarly to high- and low-dose exposure (4–7).
BEIR VII is probably the most frequently cited support for LNT, but there are serious concerns that undermine its validity as a guide to either radiation science or regulatory policies. The resulting LNT-based risks currently associated with low radiation doses appear to be overestimated and fail to reflect the actual risk–benefit balance of medical imaging.
CONCLUSION
Nuclear medicine physicians, radiologists, and radiation protection professionals aim to convey accurate radiation-related information to patients and the public, but their reliance on BEIR VII is unfortunately based on flawed interpretation of the BEIR VII Committee’s selected literature. The consequences of misdiagnoses due to imaging avoidance are potentially more immediate and harmful than any future LNT-predicted cancers avoided by stringent dose-reduction strategies. For these reasons, we call for the establishment of a BEIR VIII committee to critically reassess the validity, and use, of LNT and its derived policies.
DISCLOSURE
The opinions expressed in this article are the personal opinions of the authors. No other potential conflict of interest relevant to this article was reported.
Footnotes
Published online Feb. 23, 2018.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication December 18, 2017.
- Accepted for publication February 12, 2018.




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.