


Abstract
Immune cell–mediated attack on the liver is a defining feature of autoimmune hepatitis and hepatic allograft rejection. Despite an assortment of diagnostic tools, invasive biopsies remain the only method for identifying immune cells in the liver. We evaluated whether PET imaging with radiotracers that quantify immune activation (18F-FDG and 18F-1-(2′-deoxy-2′-fluoro-arabinofuranosyl)cytosine [18F-FAC]) and hepatocyte biology (18F-2-deoxy-2-fluoroarabinose [18F-DFA]) can visualize and quantify liver-infiltrating immune cells and hepatocyte inflammation, respectively, in a preclinical model of autoimmune hepatitis. Methods: Mice treated with concanavalin A (ConA) to induce a model of autoimmune hepatitis or vehicle were imaged with 18F-FDG, 18F-FAC, and 18F-DFA PET. Immunohistochemistry, digital autoradiography, and ex vivo accumulation assays were used to localize areas of altered radiotracer accumulation in the liver. For comparison, mice treated with an adenovirus to induce a viral hepatitis were imaged with 18F-FDG, 18F-FAC, and 18F-DFA PET. 18F-FAC PET was performed on mice treated with ConA and vehicle or with ConA and dexamethasone. Biopsy samples of patients with autoimmune hepatitis were immunostained for deoxycytidine kinase. Results: Hepatic accumulation of 18F-FDG and 18F-FAC was 173% and 61% higher, respectively, and hepatic accumulation of 18F-DFA was 41% lower, in a mouse model of autoimmune hepatitis than in control mice. Increased hepatic 18F-FDG accumulation was localized to infiltrating leukocytes and inflamed sinusoidal endothelial cells, increased hepatic 18F-FAC accumulation was concentrated in infiltrating CD4 and CD8 cells, and decreased hepatic 18F-DFA accumulation was apparent in hepatocytes throughout the liver. In contrast, viral hepatitis increased hepatic 18F-FDG accumulation by 109% and decreased hepatic 18F-DFA accumulation by 20% but had no effect on hepatic 18F-FAC accumulation (nonsignificant 2% decrease). 18F-FAC PET provided a noninvasive biomarker of the efficacy of dexamethasone for treating the autoimmune hepatitis model. Infiltrating leukocytes in liver biopsy samples from patients with autoimmune hepatitis express high levels of deoxycytidine kinase, a rate-limiting enzyme in the accumulation of 18F-FAC. Conclusion: Our data suggest that PET can be used to noninvasively visualize activated leukocytes and inflamed hepatocytes in a mouse model of autoimmune hepatitis.
Immune cell infiltration into the liver and attack on liver cells are characteristic of various diseases, including autoimmune hepatitis and hepatic allograft rejection (1,2). Immune-mediated attack on the liver can lead to significant cellular damage, and hepatic allograft rejection is a leading cause of liver retransplantation (1–4). T lymphocytes make up a sizable fraction of the liver-infiltrating leukocytes in autoimmune hepatitis and hepatic allograft rejection, and various animal models suggest a critical role for T lymphocytes in these diseases (5–9).
Current clinical methods for evaluating hepatocyte inflammation and liver-infiltrating T lymphocytes in vivo are valuable but provide an incomplete picture of disease (10–17). Hepatocellular damage can be inferred from blood levels of enzymes and metabolites enriched in or processed by the liver, including alanine transaminase, aspartate transaminase, and bilirubin. However, these measures do not always accurately reflect the degree of liver-infiltrating T lymphocytes and are not predictive of disease reversibility (10,11). The liver can also be imaged by CT, MRI, and ultrasound, but these imaging modalities provide limited functional information (12). Blood CD4 T-cell intracellular adenosine triphosphate levels, measured ex vivo after stimulation, may provide information on immune activation in the setting of a liver transplant, but the sensitivity and specificity of this approach remain controversial, and this approach provides no direct information on lymphocyte activity specifically in the liver (13). Liver biopsies remain the gold standard for assessing liver-infiltrating lymphocytes but suffer significant sampling error, carry a risk of complications, and are often challenging to interpret in diseases with heterogeneous manifestations such as hepatic allograft rejection (14–17). A method to selectively quantify and visualize CD4 and CD8 T lymphocytes and inflamed hepatocytes in the liver within the context of autoimmune hepatitis and hepatic allograft rejection could improve our understanding of these diseases and function as a potential biomarker of therapy.
We recently developed a family of PET imaging radiotracers—18F-2-deoxy-2-fluoroarabinose (18F-DFA) and 18F-2-deoxy-2-fluororibose—that measure ribose salvage activity, a pathway highly upregulated in liver hepatocytes (18,19). We showed that 18F-DFA PET quantifies functional hepatocyte density in a mouse model of drug-induced acute liver failure and that hepatic 18F-DFA accumulation is decreased in mouse models of fatty liver disease (18–20). Alternatively, the PET radiotracers 18F-FDG and 18F-1-(2′-deoxy-2′-fluoro-arabinofuranosyl)cytosine (18F-FAC) measure glucose consumption and deoxynucleoside salvage, respectively, two processes upregulated during immune cell activation (21–23). 18F-FDG and 18F-FAC PET can monitor immune activation in mouse models of systemic autoimmune disease, colitis, and immune-mediated tumor rejection (22–26). Consistent with studies indicating that PET can be used to image autoimmune disease in other organ systems (27–29), previous studies suggest that hepatic 18F-FDG accumulation is increased in a mouse model of autoimmune hepatitis and a rat model of hepatic allograft rejection (30,31), although the precise cellular source of this increased accumulation remains unclear.
Here, we study 18F-FDG, 18F-FAC, and 18F-DFA in mouse models of autoimmune and viral hepatitis. 18F-DFA instead of 18F-2-deoxy-2-fluororibose was chosen here because 18F-DFA can be synthesized on an automated radiosynthesizer and its radiochemical precursor is commercially available (32). Our results suggest that PET imaging with radiotracers that measure ribose and deoxynucleoside salvage can be used to image and quantify hepatocyte inflammation and liver-infiltrating T lymphocytes during autoimmune hepatitis.
MATERIALS AND METHODS
Mice
Male BALB/c (8–10 wk old) and nonobese diabetic scid-γ (NSG; Jackson Laboratory) (8–10 wk old) mice were used for the concanavalin A (ConA) experiments. Male FVB (8–10 wk old) and NSG (8–10 wk old) mice were used for the adenovirus experiments. BALB/c and NSG mice were from an internal UCLA mouse breeding colony. FVB mice were from the Jackson Laboratory. All animal experiments were approved in advance by the UCLA Animal Research Committee.
Treatments
ConA
Mice were injected intravenously with saline vehicle or ConA (type IV, 10 mg/kg; Sigma-Aldrich).
Adenovirus
Mice were injected intravenously and intraperitoneally with a green fluorescent protein–expressing adenovirus (100 μL each; titer, 5 × 108 plaque-forming units/mL; Cyagen).
Dexamethasone
Mice were injected with 1× phosphate-buffered saline vehicle or dexamethasone-21-phosphate (300 mg/kg) intraperitoneally 18 and 1 h before the ConA injection.
Immunohistochemical Analyses
Immunohistochemistry was performed as previously described (20) with the following exceptions: antigen retrieval was performed in citrate antigen retrieval buffer (10 mM sodium citrate, pH 6.0) for deoxycytidine kinase (dCK), glucose transporter 1 (GLUT1), CD4, CD8, CD11b, and Ki-67 or a citrate-polysorbate 20 antigen retrieval buffer (10 mM sodium citrate, 0.05% polysorbate 20, pH 6.0) for B220. The slides were blocked in 2.5% horse serum (1 h at room temperature for GLUT1, CD8, B220, CD11b, and Ki-67 or overnight at 4°C for CD4) and incubated with primary antibody (overnight at 4°C for GLUT1, CD8, B220, CD11b, and Ki-67 or 1 h at room temperature for CD4; diluted in 2.5% horse serum). Antibodies and dilutions were as follows: dCK (clone 9D4; 1:1,000; developed by the Witte Lab), GLUT1 (clone 07-1401; 1:500; Millipore), CD4 (ab183685; 1:200; Abcam), CD8 (clone 4SM15; 1:200; eBiosciences), B220 (clone RA3-6B2; 1:100; Biolegend), CD11b (ab75476; 1:1,000; Abcam), and Ki-67 (ab16667; 1:1,000; Abcam). The slides were washed in 1× phosphate-buffered saline and incubated with horse antirabbit horseradish peroxidase (1 h at room temperature for GLUT1, CD4, CD11b, and Ki-67; Vector Laboratories) or treated with Elite ABC complex following the manufacturer’s protocol (PK-6100, for B220 and CD8; Vector Laboratories).
Liver sections from anonymized patients with autoimmune hepatitis were obtained from banked biopsy samples through the UCLA Translational Pathology Core Laboratory.
PET/CT Imaging and Quantification
PET imaging and quantification were performed as previously described (20). Briefly, anesthetized mice were injected with approximately 2.96 MBq (for the ConA experiments) or approximately 0.74 MBq (for the adenovirus experiments) and imaged 1 h later by PET/CT for 10 min on a G8 PET/CT system (Sofie Biosciences). 18F-FDG was from the UCLA Translational Imaging Division. 18F-FAC and 18F-DFA were synthesized as previously described (32). The mice were injected with tracer approximately 30 h after the ConA treatment and were imaged 1–3 d before and then again 2, 4, 8, and 12 wk after the adenovirus injections. Quantified PET images are reported as signal-to-background ratio, where signal refers to radiotracer accumulation in the liver and background refers to radiotracer accumulation in a background organ (18F-FDG: right forelimb triceps; 18F-FAC and 18F-DFA: brain).
Autoradiography
Autoradiography was performed as previously described (20). Frozen slides were fixed, stained with hematoxylin and eosin, and digitally imaged.
Ex Vivo Accumulation Assays
The mice were treated with ConA as described above. Liver-infiltrating leukocytes were isolated, sorted, and treated as previously described (24,33) except that the radiotracer 3H-deoxycytidine (Moravek, Inc.) was used.
Statistical Analysis
Data are plotted as mean, with error bars representing SEM. Statistical analyses were performed using 1-way and 2-way ANOVA with Tukey and Dunnett corrections, respectively, for multiple comparisons. Statistical analyses were performed by GraphPad Prism (version 7.01).
RESULTS
Hepatic 18F-FDG, 18F-FAC, and 18F-DFA Accumulation Is Affected in a Mouse Model of Autoimmune Hepatitis
The best-studied mouse model of autoimmune hepatitis is induced by treating mice with the plant lectin ConA (9). When injected into mice, ConA produces a CD4 T-cell– and tumor necrosis factor-α–dependent but CD8 T-cell–independent hepatitis (9,34). Mice injected with ConA exhibited a robust, periportal hepatitis, characterized by liver-infiltrating leukocytes containing a high proportion of CD4 T cells (64% ± 2%), with additional contributions from CD11b-positive innate immune cells, B220-positive B cells, and CD8 T cells (9% ± 2%, 18% ± 2%, and 13% ± 1%, respectively; Fig. 1A). A sizable fraction of the liver-infiltrating leukocytes expressed Ki-67 (67% ± 4%; Fig. 1A), a marker of cell proliferation. In agreement with the literature (30), mice treated with ConA had a significantly higher accumulation of hepatic 18F-FDG than did saline vehicle–treated mice (Figs. 1B and 1C; Supplemental Fig. 1). Mice treated with ConA also exhibited higher levels of hepatic 18F-FAC accumulation and lower levels of hepatic 18F-DFA accumulation than vehicle-treated mice (Figs. 1B and 1C; Supplemental Fig. 1).
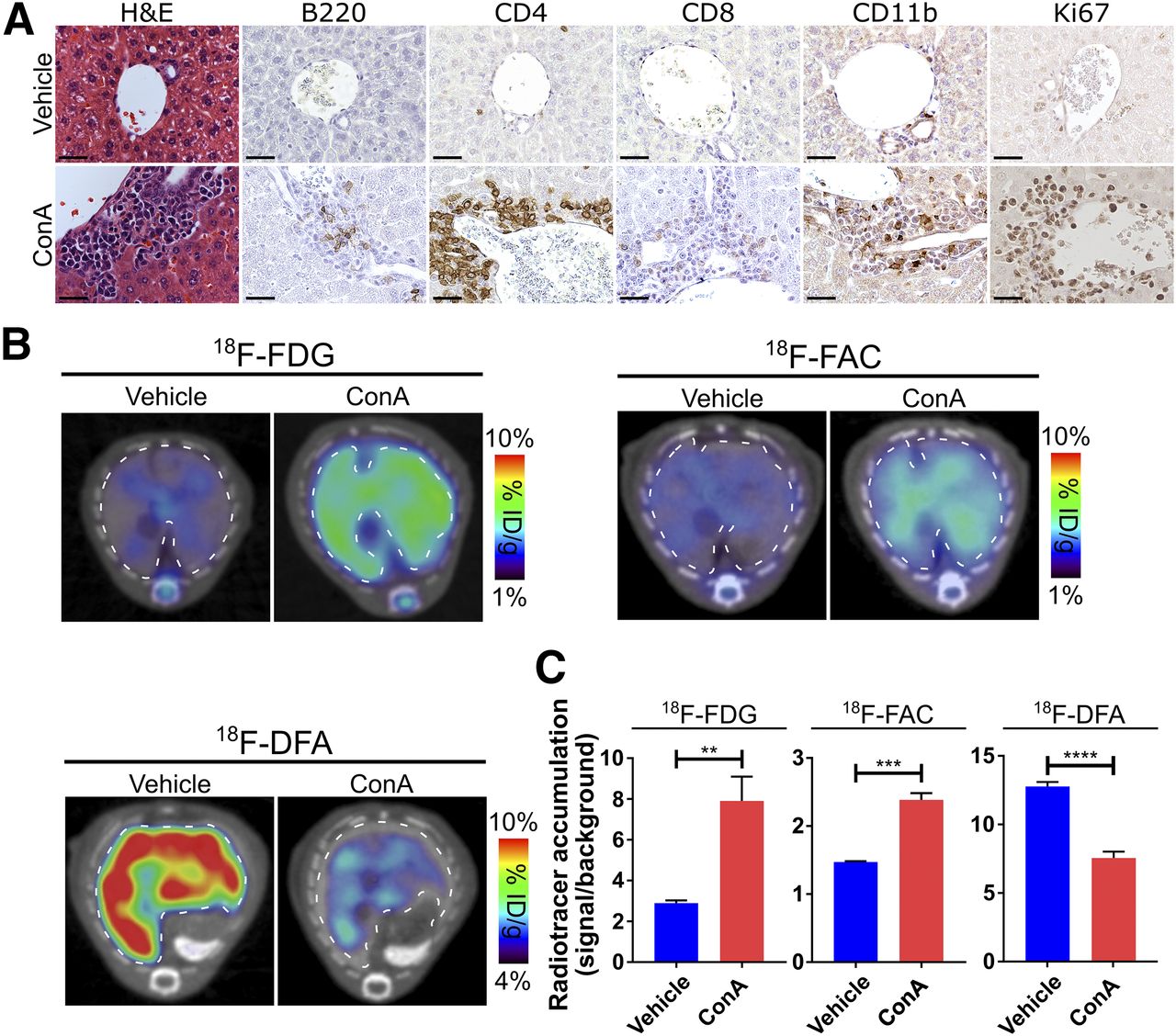
Hepatic 18F-FDG, 18F-FAC, and 18F-DFA accumulation is affected in mouse model of autoimmune hepatitis. (A) Histochemical and immunohistochemical analyses of liver sections from vehicle- and ConA-treated mice. Scale bars represent 50 μm. (B and C) Transverse PET/CT images (B) and quantification (C) of vehicle- and ConA-treated mice injected with 18F-FDG, 18F-FAC, and 18F-DFA. Livers are outlined. Quantification represents radiotracer accumulation in liver normalized to background organ. **P < 0.01. ***P < 0.001. ****P < 0.0001. H&E = hematoxylin and eosin; ID = injected dose.
Consistent with a functional role for the immune system in ConA-induced hepatitis (9), liver sections from immunocompromised NSG mice treated with ConA were indistinguishable histologically or immunohistochemically from liver sections of control mice (Supplemental Fig. 2A). No difference in hepatic 18F-FDG, 18F-FAC, or 18F-DFA accumulation was observed between vehicle- and ConA-treated immunocompromised mice (Supplemental Figs. 2B and 2C; Supplemental Fig. 1), indicating that the changes we observed in hepatic tracer accumulation in the immunocompetent mice were due to the ConA-induced hepatitis and were not unrelated effects of the ConA treatment.
Liver-Infiltrating CD4 T Cells Are a Major Source of Increased Hepatic 18F-FAC Accumulation in ConA-Induced Autoimmune Hepatitis
To better understand why hepatic 18F-FDG, 18F-FAC, and 18F-DFA accumulation is affected in ConA-induced autoimmune hepatitis, we studied the system at a cellular level. The cellular accumulation of 18F-FDG, 18F-FAC, and 18F-DFA is regulated by specific proteins (18,35,36), and changes in the levels of these proteins in the liver could in part explain changes in hepatic radiotracer accumulation. Expression and membrane localization of the facilitative glucose transporter GLUT1 is a major determinant of glucose consumption in activated lymphocytes, dCK regulates the cellular accumulation of 18F-FAC, and ribokinase is a key enzyme in the ribose salvage pathway that 18F-DFA measures (36–38).
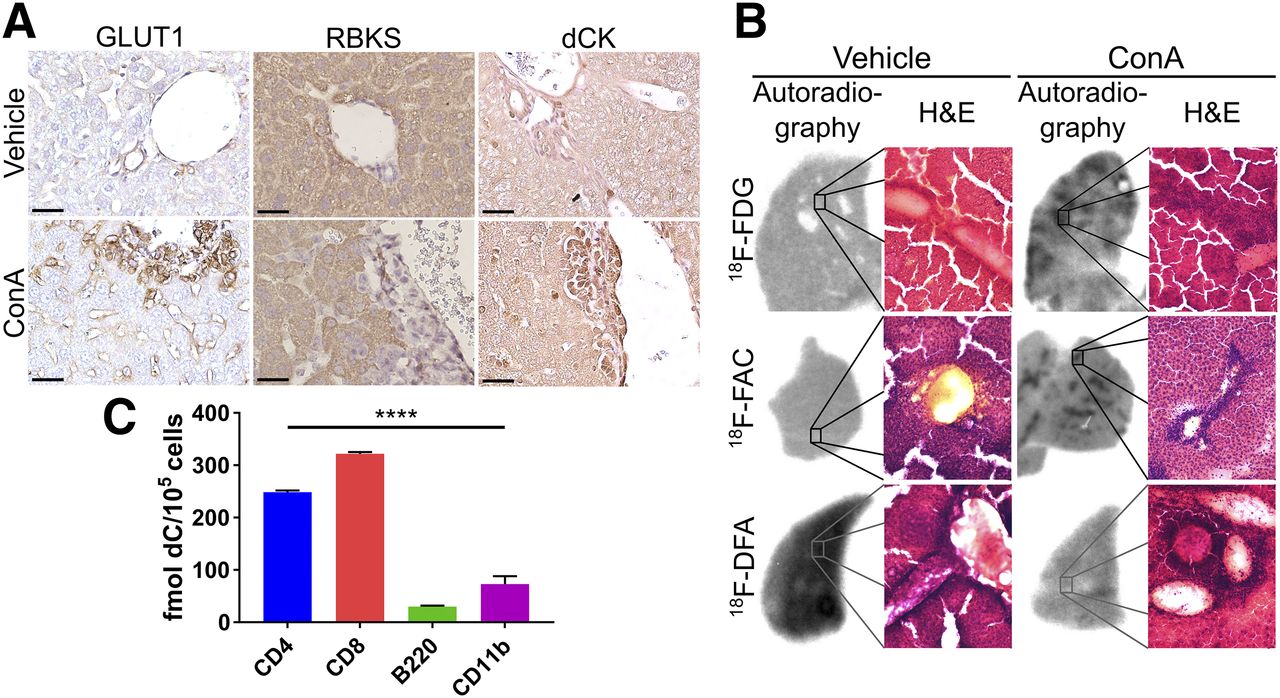
Liver sections from ConA-treated mice immunostained for GLUT1 showed high levels of membrane-localized GLUT1 in liver-infiltrating leukocytes, as well as increased GLUT1 membrane expression in leukocyte-proximal liver sinusoidal endothelial cells (Fig. 2A). Ribokinase protein levels were absent from liver-infiltrating leukocytes but present at high levels in liver hepatocytes (Fig. 2A). Liver-infiltrating leukocytes in ConA-treated mice expressed high dCK protein levels, and hepatocyte dCK levels were similar between vehicle and ConA-treated mouse liver sections (Fig. 2A). GLUT1, ribokinase, and dCK protein levels were unaffected in the livers of ConA-treated compared with vehicle-treated immunocompromised mice (Supplemental Fig. 3).

18F-FDG and 18F-FAC measure liver-infiltrating leukocytes, and 18F-DFA measures hepatocyte inflammation. (A) GLUT1, ribokinase, and dCK immunostained vehicle- and ConA-treated mouse liver sections. Scale bars represent 50 μm. (B) Autoradiographic and histologic analyses of liver sections from vehicle- and ConA-treated mice injected with 18F-FDG, 18F-FAC, or 18F-DFA. (C) Ex vivo accumulation of deoxycytidine in sorted leukocytes from livers of ConA-treated mice. ****P < 0.0001. H&E = hematoxylin and eosin; RBKS = ribokinase.
Autoradiography results of liver sections from vehicle- or ConA-treated mice injected with 18F-FDG, 18F-FAC, or 18F-DFA were coregistered with images of these same sections stained with hematoxylin and eosin. Areas of increased 18F-FDG and 18F-FAC accumulation, measured by autoradiography, were coincident with areas of significant leukocyte infiltration (Fig. 2B), suggesting that liver-infiltrating leukocytes are a major source of the increased hepatic 18F-FDG and 18F-FAC accumulation identified in ConA-treated mice. Regions of increased autoradiographic signal were 2.8-fold larger in the ConA-treated mice injected with 18F-FDG than in the ConA-treated mice injected with 18F-FAC (P < 0.0001). This finding may indicate that 18F-FAC is more specific than 18F-FDG for liver-infiltrating leukocytes, and is consistent with the immunohistochemistry results (Fig. 2A). 18F-DFA accumulation was decreased throughout the liver in ConA-treated mice and was not coincident with regions of significant leukocyte infiltration (Fig. 2B), consistent with decreased ribose salvage activity in hepatocytes throughout the liver. These data suggest that in ConA-treated mice, liver-infiltrating leukocytes consume high levels of 18F-FAC, that hepatic infiltration leukocytes and nearby liver sinusoidal endothelial cells consume high levels of 18F-FDG, and that inflamed hepatocytes decrease their 18F-DFA accumulation.
Given the potential specificity of 18F-FAC for liver-infiltrating leukocytes, the further selectivity of 18F-FAC for specific leukocyte populations was analyzed. A previous study demonstrated that during immune-mediated tumor rejection, 18F-FAC preferentially accumulated in adaptive immune cells (24). Our immunohistochemistry data suggest that CD4-, CD8-, B220-, and CD11b-positive leukocytes all infiltrate into the liver in the ConA-treated mice (Fig. 1A). CD4-, CD8-, B220-, and CD11b-positive cells were isolated from the liver of ConA-treated mice and incubated with 3H-deoxycytidine to measure deoxynucleoside salvage activity. Per cell, CD4 and CD8 T cells consumed approximately 4-fold more deoxycytidine than B220 or CD11b cells (Fig. 2C), and CD4 T cells were present in higher numbers than CD8 T cells in ConA-treated livers (Fig. 1A). Collectively, these data suggest that increased hepatic 18F-FAC accumulation predominantly measures infiltrating CD4 cells in the livers of ConA-treated mice.
Hepatic Accumulation of 18F-FDG and 18F-DFA But Not 18F-FAC Is Affected by Viral Hepatitis
Autoimmune hepatitis is one, but not the only, condition characterized by immune cell infiltration into the liver. Histologically, viral hepatitis closely resembles autoimmune hepatitis. We studied the behavior of 18F-FDG, 18F-FAC, and 18F-DFA in a model of viral hepatitis. Viral hepatitis was induced by treating mice with a replication-defective green fluorescent protein–expressing adenovirus. Immunocompromised NSG mice were used to control for non–immune-related effects of the adenoviral treatment on radiotracer accumulation. Mice treated with the adenovirus mount a robust immune attack on the liver within 2 wk of treatment that persists through 8 wk before clearing by 12 wk, leaving only histiocytes as evidence of the antecedent inflammation (Fig. 3A). Within the periportal leukocyte infiltrates, a significant proportion of the cells consisted of CD4 T cells and CD11b-positive innate immune cells whereas a smaller proportion of the cells consisted of B220-positive B cells and CD8-positive T cells (36% ± 2%, 47% ± 4%, 10% ± 2%, and 11% ± 2%, respectively). A smaller fraction of the infiltrating leukocytes was Ki-67–positive than in the autoimmune hepatitis model (37% ± 3% for this model vs. 67% ± 4% for the autoimmune hepatitis model; Figs. 1A and 3A). At the level of histology and immunohistochemistry, the livers of the immunocompromised mice were largely unaffected by the viral treatment (Supplemental Fig. 4A).
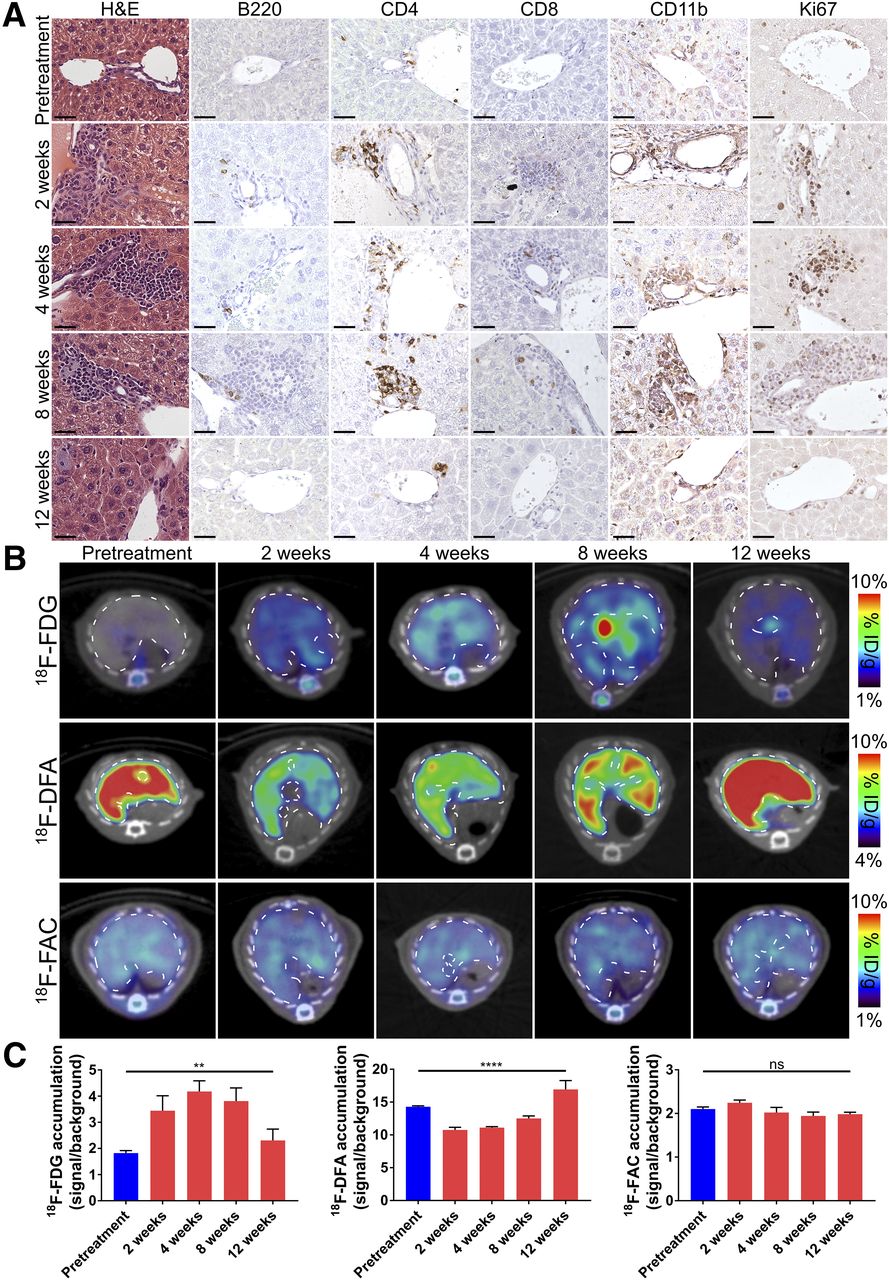
Hepatic accumulation of 18F-FDG and 18F-DFA but not 18F-FAC is affected in mouse model of viral hepatitis. (A) Histochemical and immunohistochemical analyses of liver sections from pretreatment and adenovirus-treated mice. Scale bars represent 50 μm. (B and C) Transverse PET/CT images (B) and quantification (C) of pretreatment and adenovirus-treated mice injected with 18F-FDG, 18F-DFA, and 18F-FAC. Quantification represents radiotracer accumulation in liver normalized to background organ. **P < 0.01. ****P < 0.0001. H&E = hematoxylin and eosin; ID = injected dose; ns = not significant.
Similar to results obtained in the autoimmune hepatitis model, viral hepatitis induced a significant increase in hepatic 18F-FDG accumulation that returned to pretreatment levels after the hepatitis resolved (Figs. 3B and 3C; Supplemental Fig. 5). Viral hepatitis also decreased hepatic 18F-DFA accumulation, which returned to higher than baseline levels at 12 wk after treatment (Figs. 3B and 3C; Supplemental Fig. 5). We speculate that hepatic 18F-DFA accumulation is elevated at 12 wk after treatment because of a compensatory response of the liver hepatocytes to the prior decrease in hepatic 18F-DFA accumulation. In contrast to the results from the autoimmune hepatitis model (Figs. 1B and 1C), hepatic 18F-FAC accumulation was unaffected at any time point throughout the induction and clearance of the viral hepatitis (Figs. 3B and 3C; Supplemental Fig. 5). No change in hepatic 18F-FDG, 18F-DFA, or 18F-FAC accumulation was identified in immunocompromised mice treated with adenovirus, suggesting that the above results required immune activation (Supplemental Figs. 4B, 4C, and 5).
Changes in Hepatic 18F-FAC Accumulation Can Be Used to Monitor Immunosuppressive Drug Treatments in a Mouse Model of Autoimmune Hepatitis
One potential application for a PET strategy that can quantify liver-infiltrating CD4 and CD8 T lymphocytes is monitoring the response of these cells to immunosuppressive therapies, similar to the role of PET in other fields, including oncology (39). Studies suggest, and we found, that pretreatment of mice with the immunosuppressive drug dexamethasone can block ConA-induced autoimmune hepatitis (Fig. 4A) (9). Mice treated with dexamethasone 18 and 1 h before treatment with ConA had significantly lower levels of hepatic 18F-FAC accumulation than mice treated with ConA and vehicle (Figs. 4B and 4C). This finding suggests that PET imaging with 18F-FAC can be used as a biomarker for immunomodulatory therapies developed to treat autoimmune hepatitis.

Changes in hepatic 18F-FAC accumulation can be used to monitor immunosuppressive drug treatments in mouse model of autoimmune hepatitis. (A) Representative hematoxylin- and eosin-stained liver sections of mice treated with vehicle, ConA, or ConA and dexamethasone. Scale bars represent 50 μm. (B) Transverse PET/CT images of hepatic 18F-FAC accumulation in mice treated with vehicle, ConA, or ConA and Dexa. (C) Quantification of hepatic 18F-FAC accumulation in mice treated with vehicle, ConA, or ConA and Dexa. Quantification represents radiotracer accumulation in liver normalized to background organ. ****P < 0.0001. Dexa = dexamethasone; H&E = hematoxylin and eosin; ID = injected dose; ns = not significant.
Liver-Infiltrating Leukocytes in Patients with Autoimmune Hepatitis Express dCK
Our data suggest that 18F-FAC may selectively image CD4 and CD8 T cells during autoimmune hepatitis. Whether this is true in humans remains to be determined and is the basis for future studies. To assess deoxynucleoside salvage activity in liver-infiltrating leukocytes in human patients with autoimmune hepatitis, we immunostained tissue sections from biopsy samples of patients with autoimmune hepatitis for dCK. Liver-infiltrating leukocytes in these liver sections stained strongly for dCK (Fig. 5), providing early evidence that PET tracers that measure dCK activity can potentially be used to quantify liver-infiltrating lymphocytes in humans with autoimmune hepatitis.

Liver-infiltrating leukocytes in patients with autoimmune hepatitis express dCK. Histochemical and immunohistochemical analyses of liver biopsies from patients with autoimmune hepatitis. Scale bars represent 50 μm. H&E = hematoxylin and eosin.
DISCUSSION
In our studies, 18F-FAC selectively accumulates in the autoimmune but not viral hepatitis model. Hepatic 18F-FDG and 18F-DFA accumulation was affected to a similar extent in the autoimmune and viral hepatitis models, suggesting that the difference in 18F-FAC accumulation between the two models is not due solely to differences in the magnitude of the immune response. Determining the precise mechanism for the selectivity of 18F-FAC for autoimmune over viral hepatitis is beyond the scope of this work. However, previous studies suggest a strong correlation between 18F-FAC accumulation and cell cycle in immune cells (24). The ConA-induced autoimmune hepatitis model has faster kinetics than the viral hepatitis model. ConA induces autoimmune hepatitis within 24 h of injection that begins to clear by 48 h. The viral hepatitis is induced within 2 wk and takes 12 wk to clear. Consistent with these kinetics, a higher percentage of liver-infiltrating leukocytes is positive for the cellular proliferation marker Ki-67 in the ConA-induced autoimmune hepatitis model than in the viral hepatitis model (67% ± 4% vs. 37% ± 3%, respectively; Figs. 1A and 3A). Additionally, we showed high deoxynucleoside salvage activity in CD4 T cells in the autoimmune hepatitis model, and this model has a higher percentage of CD4 T cells among the liver-infiltrating leukocytes than did the viral hepatitis model (64% ± 2% vs. 36% ± 2%, respectively). This finding may suggest that 18F-FAC accumulates in liver-infiltrating lymphocytes in the ConA-induced autoimmune hepatitis model because of a greater percentage of CD4 cells and greater cell proliferation among the infiltrating immune cells in this model.
We found that hepatic ribose salvage activity is decreased uniformly throughout liver hepatocytes in both the autoimmune and the viral hepatitis models and is not localized to areas of significant leukocyte infiltration. Little is known about the regulation of ribose salvage activity. However, the fact that ribose salvage is affected in hepatocytes adjacent to and distant from the infiltrating immune cells suggests a potential role for cytokines in this process. Previous studies reported that during ConA-induced autoimmune hepatitis, plasma protein and liver messenger RNA levels of cytokines, including tumor necrosis factor, IFN-γ, IL-2, IL-4, IL-6, IL-10, and IL-12, are all elevated (40). Whether cytokines regulate ribose salvage activity in this model remains to be determined.
Our studies suggest that deoxynucleoside salvage activity in the liver, measured by 18F-FAC accumulation, is higher in a mouse model of autoimmune hepatitis than in control mice. dCK is a rate-limiting enzyme for deoxynucleoside salvage (36). The precise role of deoxynucleoside salvage in T-cell activation is not well described, but genetic inactivation of dCK in mice blocks lymphocyte development and leads to significantly lower lymphocyte numbers (36). Recently, small-molecule inhibitors of dCK have been developed (41). Although further studies are required, our results may suggest new therapeutic opportunities for targeting dCK in the context of autoimmune hepatitis.
Implications for Human Studies
Diagnosing autoimmune hepatitis and hepatic allograft rejection and monitoring therapeutic responses in these diseases remain a challenge (42–44). In routine clinical practice, autoimmune hepatitis is diagnosed through descriptive criteria with consideration of features such as liver histology and serum biochemistries. A semiquantitative scoring system used in clinical trials that attributes positive or negative points to the presence or absence of alterations in liver histology, serum biochemistry, and serum autoantibodies improves on the descriptive criteria (1). Similarly, hepatic allograft rejection is diagnosed through a combination of liver biochemistry measurements and histologic features (2). All of these approaches depend on interpreting the results of a liver biopsy, which can be subject to significant sampling and reader error and is not without risk (14–17). We hypothesize that PET imaging with 18F-FAC (or 18F-FAC derivatives) can provide a tomographic, whole-organ, quantitative assessment of activated liver-infiltrating lymphocytes in these diseases and can complement biopsies. 18F-DFA PET might provide the same with respect to the presence of inflamed hepatocytes.
Autoimmune hepatitis and hepatic allograft rejection are treated with immunosuppressive drugs (1,2). These therapies are often effective but have narrow therapeutic windows, significant side effects, and high patient-to-patient variability in their pharmacokinetics and pharmacodynamics (45,46). Without a method for quantifying activated immune cells in the liver, identifying the correct dose of an immunosuppressive drug for an individual patient that blocks immune attack on the liver while limiting unwanted side effects remains a significant clinical challenge (47). The ability to directly and quantitatively measure liver-infiltrating lymphocytes—a desired target of these immunomodulatory drugs—could serve as a valuable biomarker for assessing efficacy. Here, we show in a mouse model that 18F-FAC PET can quantitatively assess liver-infiltrating lymphocytes and can be used to monitor the efficacy of an immunomodulatory drug.
The most prevalent hepatitis-inducing viruses in humans are hepatitis B and C. These diseases can occur as acute infections that resolve or can persist as chronic infections (48). Insofar as the adenovirus infection in mice resolves, it models an acute rather than a chronic infection. The lack of any effect on hepatic radiotracer accumulation in the adenovirus-treated immunocompromised mice suggests that the effects identified in the adenovirus-treated immunocompetent mice are solely due to immune attack on the liver and not to the adenovirus infection by itself. Chronic hepatitis infections are characterized by exhausted and impaired immune cell responses to the virus, an immunologic state associated with decreased glucose consumption (48,49). We would have to study a chronic hepatitis model to understand the behavior of the radiotracers during a chronic infection. However, we can speculate that the differences we identify here in hepatic 18F-FDG and 18F-DFA accumulation during the adenoviral infection would likely be muted during a chronic infection.
In a recent study, we showed that ribose salvage activity, the biochemical pathway measured by 18F-DFA, is similar between mouse and human hepatocytes in culture and when engrafted into a mouse liver (20). This finding suggests that the 18F-DFA imaging results presented here may be translatable into human studies.
18F-FAC images dCK activity in mice but is subject to deamination by the enzyme cytidine deaminase (50). Cytidine deaminase activity is higher in most human tissues than in mouse tissues, and 18F-FAC accumulation in lymphoid organs such as the spleen is lower in humans than in mice (24,51,52). 18F-FAC accumulation is also low in the human liver (52). The PET radiotracer 2-chloro-2′-deoxy-2′-18F-fluoro-9-d-arabinofuranosyl-adenine is a substrate for dCK and resistant to deamination by cytidine deaminase (53). This radiotracer accumulates at high levels in lymphoid organs in humans but also at high levels in the healthy human liver (54), and high accumulation in the healthy liver would disfavor its use for imaging lymphocytes in the human liver. However, whether activated human CD4 and CD8 T cells could accumulate sufficient 18F-FAC in the setting of high intracellular and peripheral cytidine deaminase activity to produce a PET image remains to be determined.
CONCLUSION
Our results suggest that PET imaging can potentially provide a quantitative, noninvasive method for monitoring hepatocytes and liver-infiltrating T cells during autoimmune hepatitis and in response to therapy. Previous studies suggest that 18F-FAC can be used to visualize a mouse model of systemic autoimmunity and colitis (23,26). The value with respect to treatment would be in directly assessing the targets of disease-modifying therapies. Collectively, these studies begin to suggest a potential role for 18F-FAC PET and PET tracers with organ-specific accumulation to visualize and quantify the development and treatment of autoimmune diseases.
DISCLOSURE
This work was supported by NIH grant R21AI119916 (to Peter Clark), Parker Institute for Cancer Immunotherapy (PICI) grant 20163828 (to Owen Witte), and the Broad Stem Cell Research Center at UCLA. Peter Clark and Owen Witte are inventors on a patent, held by the Regents of the University of California, that describes the 18F-DFA radiotracer. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank the UCLA Translational Pathology Core Laboratory, the Crump Preclinical Imaging Technology Center, and the Crump Cyclotron and Radiochemistry Technology Center for assisting in these studies. We thank Ralph and Marjorie Crump for their gift to the UCLA Crump Institute for Molecular Imaging.
Footnotes
Guest Editor: Todd Peterson, Vanderbilt University
Published online Apr. 26, 2018.
- © 2018 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication February 21, 2018.
- Accepted for publication April 23, 2018.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}