


Abstract
18F-FDG PET imaging is routinely used to investigate brown adipose tissue (BAT) thermogenesis, which requires mitochondrial uncoupling protein 1 (UCP1). It remains uncertain, however, whether BAT 18F-FDG uptake is a reliable surrogate measure of UCP1-mediated heat production. Methods: UCP1 knockout (KO) and wild-type (WT) mice housed at thermoneutrality were treated with the selective β3 adrenergic receptor agonist CL 316, 243 and underwent metabolic cage, infrared thermal imaging and 18F-FDG PET/MRI experiments. Primary brown adipocytes were additionally examined for their bioenergetics by extracellular flux analysis as well as their uptake of 2-deoxy-3H-glucose. Results: In response to CL 316, 243 treatments, oxygen consumption, and BAT thermogenesis were diminished in UCP1 KO mice, but BAT 18F-FDG uptake was fully retained. Isolated UCP1 KO brown adipocytes exhibited defective induction of uncoupled respiration whereas their glycolytic flux and 2-deoxy-3H-glucose uptake rates were largely unaffected. Conclusion: Adrenergic stimulation can increase BAT 18F-FDG uptake independently of UCP1 thermogenic function.
- animal imaging
- neuroendocrine
- PET/MRI
- brown adipose tissue
- glucose metabolism
- thermogenesis
- uncoupling protein 1
In human PET imaging studies, brown adipose tissue (BAT) 18F-FDG uptake has been shown to be markedly increased by controlled cold exposure (1,2) and agonism of β3 adrenergic receptors (3), correlating positively with changes in energy expenditure (1,3). Similar findings have been made in small-animal PET imaging studies performed on mice (4,5).
Mitochondrial uncoupling protein 1 (UCP1) endows both human and mouse brown adipocytes with thermogenic capacity (6). It is widely assumed that BAT 18F-FDG uptake provides an indirect quantitative measure of thermogenesis. We therefore sought to formally test this basic assumption by performing 18F-FDG PET/MRI experiments on UCP1-deficient mice that exhibit defective BAT thermogenesis and uncoupled respiration in response to the selective β3 adrenergic receptor agonist CL 316, 243 (7,8).
MATERIALS AND METHODS
Animals
Experiments were performed on female wild-type (WT) C57BL/6J mice and UCP1 knockout (KO) littermates (Jackson Laboratory) aged 3–9 mo and were all approved by the Institutional Animal Care and Use Committee at the University of Leipzig (TVV 63/13 and TVV 04/12). Animals were housed in an incubator (Memmert GmbH & Co. KG) set at thermoneutrality (30°C) and 60% humidity on a 12-h light–dark cycle and had free access to a high-fat high-sugar diet (Sniff GmbH) to potentiate thermogenic responses to adrenergic stimulation (8) unless otherwise indicated. In line with the findings of Feldmann et al. (8), UCP1 KO mice housed under these conditions consistently showed higher body weights than WT littermates (Table 1).
Weight of UCP1 KO Mice Compared with Their WT Littermates at Time of Experiments
Indirect Calorimetry
Animals were transferred to metabolic cages (TSE Systems) immediately after intraperitoneal injection of the selective β3 adrenergic receptor agonist CL 316, 243 (1 mg/kg) (Tocris) (7), and oxygen consumption was measured for 3 h by indirect calorimetry.
Core Body Temperature Measurements
Core body temperature was measured using a rectal probe (Thermalert) at baseline and at 15-min intervals for 1 h after intraperitoneal injection of CL 316, 243 (1 mg/kg) (7).
Infrared Thermal Imaging
Infrared thermal imaging (9) was performed using a VarioCAM thermal camera (InfraTec GmbH). Each mouse was placed on a cage top at a fixed distance away from the camera lens. Serial 1-s images (10 Hz) were taken in triplicate at baseline and at 15-min intervals for 1 h after intraperitoneal injection of CL 316, 243 (1 mg/kg). For analysis, a constantly sized circular region of interest was drawn over BAT, and the average temperature was recorded.
PET/MRI
Small-animal PET/MRI (Mediso Medical Imaging Systems) was performed as previously described (10). Overnight fasted mice received intraperitoneal injections of either CL 316, 243 (1 mg/kg) (4) or 0.9% saline along with an intraperitoneal injection of 18.8 ± 0.4 MBq of 18F-FDG (Department of Nuclear Medicine, University Hospital Leipzig). Animals were then returned to their home cages for 45 min and subsequently anesthetized with isoflurane (1.8%, 0.35 L/min) delivered in a 60% oxygen–40% air mixture (MCQ Instruments) and transferred to the PET/MRI scanner. A 15-min static PET scan was initiated, during which animals were maintained at 37°C under isoflurane anesthesia. For analysis, mean SUVs of 18F-FDG by BAT were calculated.
Cellular Bioenergetics and 2-Deoxy-3H-Glucose Assay
Stromal vascular cells were isolated from interscapular BAT and differentiated as previously described (11). On day 7 of differentiation, mature brown adipocytes were washed twice with assay medium (XF Dulbecco modified Eagle medium supplemented with 25 mM glucose, 2 mM pyruvate, and 4% bovine serum albumin [w/v]), followed by incubation at 37°C without CO2 for 20 min. Cells were transferred to an XF96 Extracellular Flux Analyzer (Seahorse Bioscience), and after 5 basal values oligomycin (5 μg/mL) was injected to induce uncoupled respiration followed by an injection of 100 μM 2-,4-dinitrophenol. To correct for nonmitochondrial respiration and nonglycolytic sources of changes in extracurricular acidification rate, a cocktail of 2.5 μM rotenone, 2.5 μM antimycin A, and 2.5 μM 2-deoxyglucose was injected. For analysis, basal values were averaged as were the 3 values immediately after oligomycin treatment and the 3 values at 1 h after oligomycin treatment.
For the 2-deoxy-3H-glucose uptake assay, mature brown adipocytes were cultivated overnight in serum-free medium and were stimulated with 1 μM CL 316, 243 (Sigma-Aldrich GmbH) (12) or distilled water followed by incubation at 37°C in 5% CO2 for 110 min. Cells were washed once with prewarmed phosphate-buffered saline, then 2-deoxy-3H-glucose (0.5 kBq/mL) and 2-deoxy-glucose (100 μM) were added, followed by incubation at 37°C in 5% CO2 for 10 min. After being washed with ice-cold phosphate-buffered saline, cells were lysed with radioimmunoprecipitation assay buffer and transferred to a liquid scintillation analyzer (PerkinElmer) for 3 h. Uptake rates of 2-deoxy-3H-glucose were normalized to protein concentration.
Statistics
Two-way ANOVA with the Sidak post hoc test was used to assess differences between groups (GraphPad Software Inc.). P values of less than 0.05 were considered significant.
RESULTS
The increase in whole-body oxygen consumption caused by CL 316, 243 treatment was blunted in UCP1 KO compared with WT mice (F1, 90 = 554.8, P < 0.0001) (Fig. 1A) as was the rise in core body temperature (F1, 35 = 10.77, P = 0.0023) (Fig. 1B).

(A) Diminished oxygen consumption (VO2) and (B) core body temperature (Tb) changes in response to adrenergic stimulation in UCP1 KO mice. Data points represent mean ± SEM of 3–5 animals per group. *P < 0.05. **P < 0.01. ****P < 0.0001.
Representative thermal images are presented in Figure 2A. Unlike UCP1 KO mice, WT mice showed an increase in interscapular BAT temperature in response to CL 316, 243 treatment (F1, 30 = 18.53, P = 0.0002) (Fig. 2B).
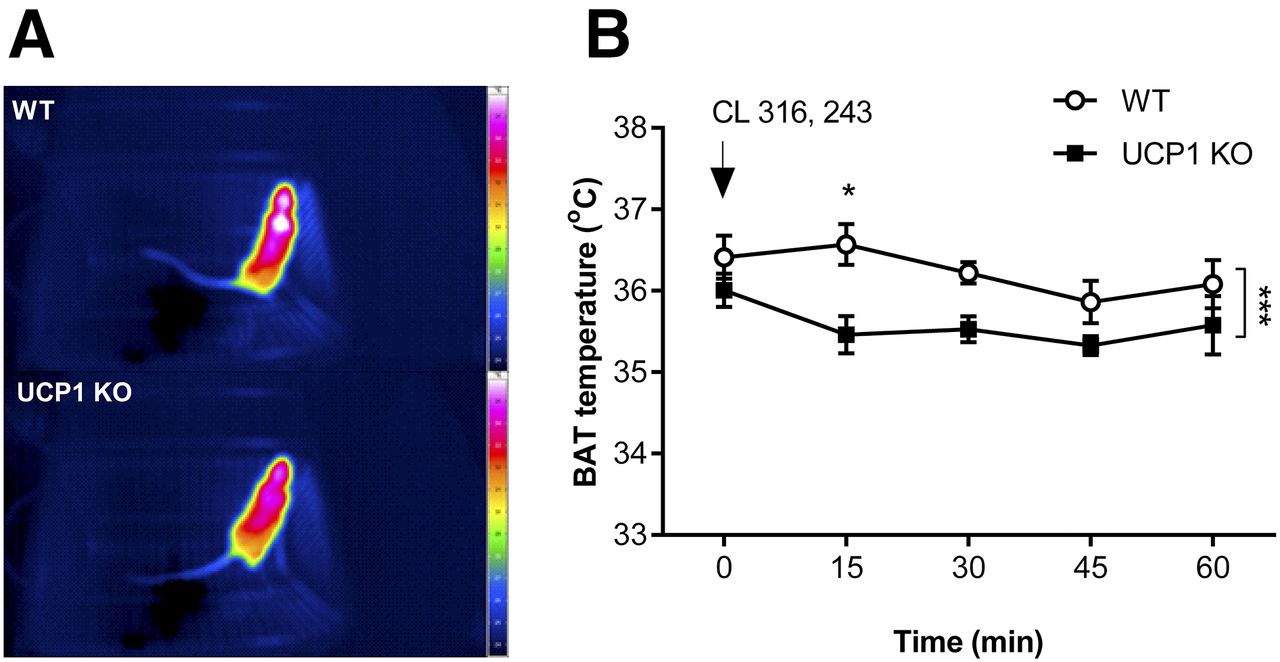
(A) Representative thermal images of WT (top) and UCP1 KO (bottom) mice taken 15 min after CL 316, 243 injections. (B) Defective BAT thermogenesis in response to adrenergic stimulation in UCP1 KO mice. Data points represent mean ± SEM of 4 animals per group. *P < 0.05. ***P < 0.001.
Representative 18F-FDG PET/MRI data are presented in Figure 3A. Both WT and UCP1 KO mice showed an increased mean SUV for18F-FDG in BAT in response to CL 316, 243 treatment relative to vehicle treatment (P < 0.0001) (Fig. 3B), with no significant effect of genotype found (F1, 14 = 0.17, P = 0.68).

(A) Representative PET/MRI data of WT and UCP1 KO mice taken 45–60 min after CL 316, 243/saline and 18F-FDG injections. White arrows in each image point to interscapular BAT. (B) Retained BAT 18F-FDG uptake in response to adrenergic stimulation in UCP1 KO mice. Bars represent mean ± SEM of 4–5 animals per group. ****P < 0.0001.
Extracellular flux analysis of cultured primary brown adipocytes revealed that 1 h after oligomycin treatment, reporting induction of uncoupled respiration (13), oxygen consumption rates were significantly lower for UCP1 KO than WT cells (P < 0.001) (Fig. 4A). In contrast, extracellular acidification rates, reporting glycolytic activity (13), tended to be similar between UCP1 KO and WT cells at this time point after oligomycin treatment (P = 0.07) (Fig. 4B). Both WT and UCP1 KO brown adipocytes showed an increase in 2-deoxy-3H-glucose uptake in response to CL 316, 243 treatment compared with vehicle (P = 0.01 and P < 0.0001, respectively) (Fig. 4C), with no significant effect of genotype found (F1, 12 = 0.00045, P = 0.97).
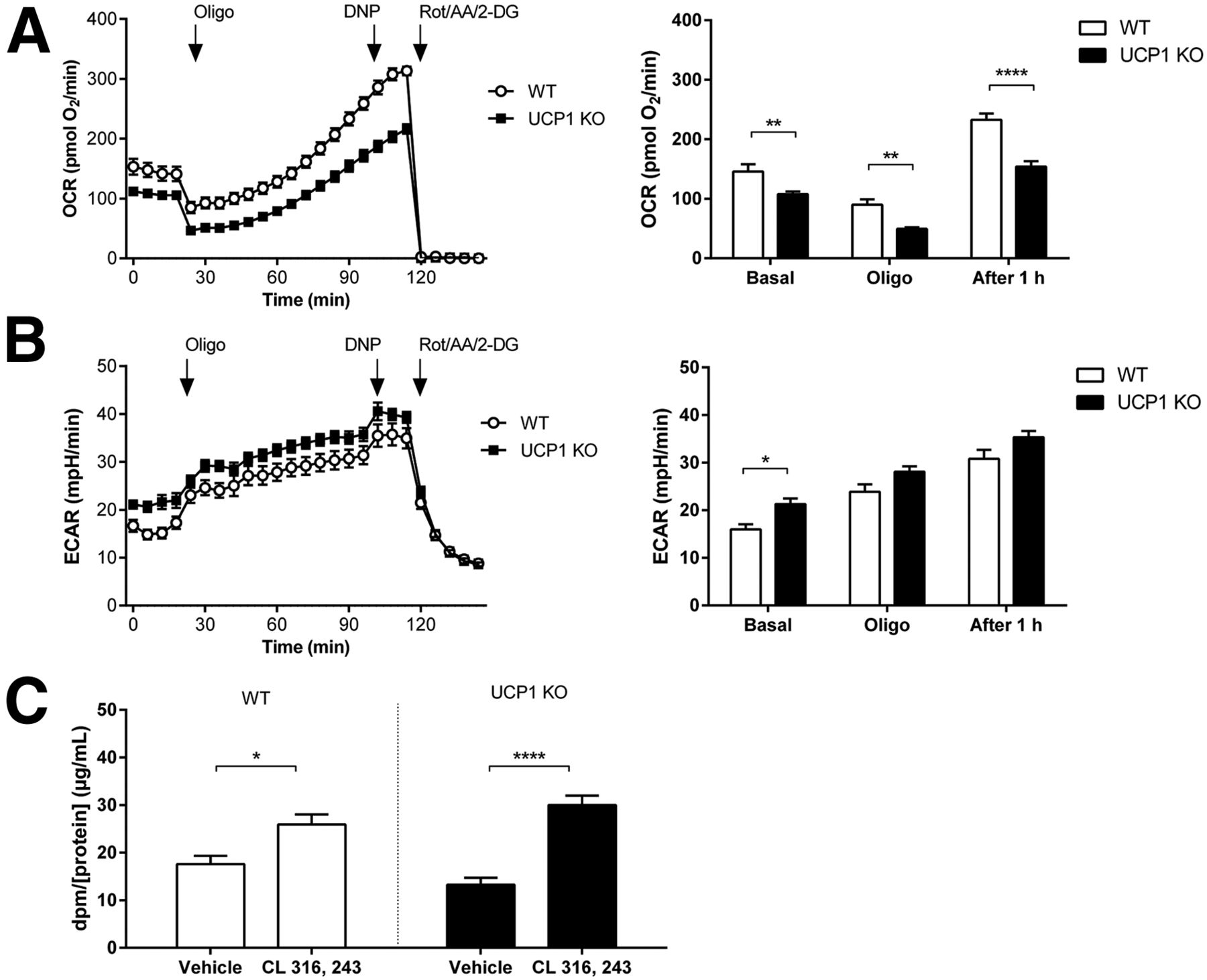
(A) Impaired induction of uncoupled respiration but normal (B) glycolysis and (C) 2-deoxy-3H-glucose uptake in UCP1 KO mouse primary brown adipocytes. Data points represent mean ± SEM of 4–6 cultures per group. 2-DG = 2-deoxyglucose; AA = antimycin A; DNP = dinitrophenol; dpm = dose per minute; ECAR = extracellular acidification rate; Rot = rotenone. *P < 0.05. **P < 0.01. ****P < 0.0001.
DISCUSSION
The findings from the present study demonstrate that glucose metabolism can increase in brown adipocytes in the absence of UCP1-mediated uncoupled respiration and heat production.
In mouse primary brown adipocytes treated with CL 316,243, glucose is transported into the cell and through the action of diacylglycerol acyltransferase 2 feeds into specialized lipid droplet pools, which are simultaneously hydrolyzed (14). The released free fatty acids are then either rapidly oxidized or activate UCP1 in mitochondria (14). Our findings of similar β3 adrenergic receptor–stimulated 18F-FDG/2-deoxy-3H-glucose uptake between WT and UCP1 KO brown adipocytes suggest that UCP1 (thermogenic) function does not provide feedback, on the initial step at least, of this concerted process.
Noradrenaline treatment was previously found not to stimulate BAT 2-deoxy-3H-glucose uptake in male UCP1 KO mice (15). The discordance with the findings from the present study could be explained by our use of female UCP1 KO mice. Indeed, cold-stimulated BAT 18F-FDG uptake is diminished in male UCP1 KO mice but not in female counterparts (16).
It is unlikely that the low BAT 18F-FDG uptake in the present study is diet related (17). This is because we found in preliminary experiments performed on high-fat high-sugar fed mice a much higher amount of BAT 18F-FDG uptake when animals were housed and scanned at room temperature (22°C). This suggests that experimental conditions even only mildly below thermoneutrality (30°C for mice) (8) can significantly stimulate BAT. Therefore, studies under thermoneutral conditions are optimal because the low BAT activity constitutes a true basal state allowing the effects of novel stimulators of BAT to be examined in isolation.
CONCLUSION
BAT 18F-FDG uptake and UCP1-mediated heat production can be dissociated. Additional techniques that measure BAT thermogenesis apart from 18F-FDG PET imaging and blood flow measurements (18) are required in model systems in which UCP1 is expected not to be functional. Because UCP1 deficiency is not the standard physiologic scenario in humans, caution is, however, warranted when translating our findings.
DISCLOSURE
The work in this manuscript was funded by the German Federal Ministry of Education and Research, the German Research Foundation, and the German Centre for Diabetes Research. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank Dr. Kerstin Krause for assistance with the experiments.
Footnotes
↵* Contributed equally to this work.
Published online Jan. 12, 2017.
- © 2017 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication November 2, 2016.
- Accepted for publication December 16, 2016.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- High-fructose feeding suppresses cold-stimulated brown adipose tissue glucose uptake in young men independently of changes in thermogenesis and the gut microbiome
- Brown adipose tissue lipoprotein and glucose disposal is not determined by thermogenesis in uncoupling protein 1-deficient mice
- Blocking mitochondrial pyruvate import causes energy wasting via futile lipid cycling in brown fat
- Substrate fluxes in brown adipocytes upon adrenergic stimulation and uncoupling protein 1 ablation
- CPEB2-dependent translation of long 3'-UTR Ucp1 mRNA promotes thermogenesis in brown adipose tissue
- Brown adipocyte glucose metabolism: a heated subject
- Accurate quantification of brown adipose tissue mass by xenon-enhanced computed tomography