


Abstract
Targeted α-therapy is a promising treatment option for various types of malignant tumors. Radiolabeled cancer-seeking agents, however, undergo degradation, resulting in a certain percentage of free radionuclide in the body. The radiohalogen 211At accumulates in various tissues, with specifically high uptake in the thyroid. When normal thyroid function is disturbed because of ionizing radiation (IR) exposure, deleterious effects can occur in tissues that depend on thyroid hormone (TH) regulation for normal physiologic function. However, knowledge of systemic effects is still rudimentary. We previously reported similarities in transcriptomic regulation between the thyroid and other tissues despite large differences in absorbed dose from 211At. Here, we present supportive evidence on systemic effects after 211At administration. Methods: Expression microarray data from the kidney cortex and medulla, liver, lungs, and spleen were used from previous studies in which mice were intravenously injected with 0.064–42 kBq of 211At and killed after 24 h or injected with 1.7 kBq of 211At and killed after 1, 6, or 168 h. Controls were mock-treated and killed after 24 h. Literature-based gene signatures were used to evaluate the relative impact from IR- or TH-induced regulation. Thyroid- and TH-associated upstream regulators as well as thyroid-related diseases and functions were generated using functional analysis software. Results: Responses in IR- or TH-associated gene signatures were tissue-specific and varied over time, and the relative impact of each gene signature differed between the investigated tissues. The liver showed a clear dominance of TH-responding genes. In the kidney cortex, kidney medulla, and lungs, the TH-associated signature was detected to at least an extent similar to the IR-associated signature. The spleen was the single tissue showing regulation of only IR-associated signature genes. Various thyroid-associated diseases and functions were inferred from the data: L-triiodothyronine, TH, TH receptor, and triiodothyronine (reverse) were inferred as upstream regulators with differences in incidence and strength of regulation depending on tissue type. Conclusion: These findings indicate that transcriptional regulation in various nonthyroid tissues was—in part—induced by thyroid (hormone)-dependent signaling. Consideration of the systemic context between tissues could contribute to normal tissue risk assessment and planning of remedial measures.
An organism is a complex network of interdependent functional units, which can be studied on the level of interacting molecules, communicating cells in a tissue, or organs within the body. Although understood in principle, interdependencies between cells, tissues, and organs can remain elusive in experimental settings in vivo and pose a challenge for estimation and limitation of long-term side effects in medical practice. In radionuclide therapy, cancer-targeting agents are labeled with radionuclides and administered intravenously or locally to specifically bind to and kill malignant cells—that is, to cause a strong and localized therapeutic effect in a systemic setting. In the 1960s and 1970s, several experiments were conducted studying systemic effects in an organism on (localized) ionizing radiation (IR) exposure (1–5). This venue, however, became less prominent in the following decades when research focused on in vitro studies and the molecular mechanisms underlying cellular responses. In a different fashion, the systemic aspect recurred with the description of the bystander phenomenon in the 1990s (6–9), showing that nonhit cells within an exposed population responded to (molecular) signals from hit cells mimicking IR-induced damage and response, although the former were not subjected to ionization events. In 2005, Mothersill et al. discussed the systemic aspect and multiorgan involvement in vivo, however in a high-dose exposure setting (10). Knowledge of systemic effects to low-dose exposure in vivo is still scarce but essential, in particular for nuclear medicine. The α-emitter 211At is proposed for targeted radiotherapy of various malignancies, in particular metastasized tumors (11–14). A certain percentage of the labeled compound, however, usually degrades and accumulates in the thyroid and to a lesser degree in various other tissues (15–20), resulting in differential low-dose IR exposure within the body.
In previous work, we studied transcriptomic regulation profiles and individual gene regulation in response to 211At, in particular after 24 h after injection of 0.064–42 kBq of 211At in BALB/c nude mice (21,22). Despite large differences in absorbed dose level between the thyroid and less accumulating tissues such as the kidneys, liver, lungs, and spleen, we observed distinct similarities in genome-wide transcriptional intensity between the thyroid and these tissues. We then hypothesized that observed responses were not exclusively induced by IR exposure in each tissue, but were in part influenced by, for example, thyroid hormones (TH) or systemic factors originating from the dominantly 211At-accumulating thyroid gland (22). Although the study design did not allow for direct evidence of thyroid-dependent effects in nonthyroid tissues, the comprehensive microarray data can be analyzed with dedicated statistical software that can predict changes in biologic functions indicative of a certain stimulus or outcome.
The aim of the present study was to determine whether the observed transcriptomic regulation in nonthyroid tissues indicated systemic effects originating from the thyroid based on previous microarray data. We performed functional analysis with regard to TH regulation and pathway regulation related to thyroid disorders. Furthermore, we used signature gene analysis to assess the relative impact that IR exposure or TH-induced regulation may have on transcriptomic responses in nonthyroid tissues.
MATERIALS AND METHODS
Experimental Design
This follow-up study was performed on microarray data obtained from 4 experimental studies discussed elsewhere (21–24). Briefly, 211At was produced via the 209Bi(α,2n)211At reaction at the Cyclotron and PET Unit at Rigshospitalet in Copenhagen, Denmark, and express-delivered to Gothenburg, Sweden. Free 211At was prepared via dry-distillation of the irradiated target on-site with isolation yields of 92% ± 3%, with distillation taking between 1 and 2 min as described by Lindegren et al. (25). Altogether, the procedure was performed within about 1–1.5 half-lives after irradiation of the target. The radioactivity purity was checked carefully after distillation, being close to 100% in the solutions used. The solutions used in animal experiments were prepared and administered quickly afterward, to ensure that 211At was in the state of astatide.
Six-month-old female BALB/c nude mice (Charles River) were intravenously injected with 211At prepared in saline solution at various activity concentrations or mock-treated as controls (n = 3/group). In the dose-range studies, responses were analyzed after 24 h after administration of 0.064, 0.64, 1.8, 14, or 42 kBq of 211At (21,22). In the short-time studies, an activity of 1.7 kBq was administered and responses studied after 1 h, 6 h, or 7 d (23,24). The control group was mock-treated with physiologic saline and killed after 24 h. The administered activities were chosen to deliver a desired absorbed dose over a certain period to the thyroid. In the dose-range study, the administered activities were chosen to cover a wide dose range from very low over moderate to very high absorbed dose to the thyroid over 1 d. In the time-range study, a low activity was chosen to result in (very) low absorbed dose even over 1 wk. BALB/c nude mice were chosen because the strain is routinely used in experimental research on human tumor xenografts. The experiments were designed in the context of studying normal tissue effects in tumor-bearing mice treated with radionuclides or radiolabeled agents. Animal handling and preparation of tissue samples have been described previously (22). Tissue-specific absorbed dose was adapted from previously published work (21–24); briefly, the absorbed dose was calculated using the MIRD formalism (26). All studies were approved by the Ethical Committee on Animal Experiments in Gothenburg, Sweden.
Transcriptional Analysis
Transcriptional analysis was performed on microarray data deposited in National Center for Biotechnology Information’s Gene Expression Omnibus with accessions GEO:GSE32306 (21), GEO:GSE40806 (22), GEO:GSE56894 (23), and GEO:GSE66089 (24). Briefly, genome-wide transcriptional regulation was analyzed using MouseRef-8 Whole-Genome Expression BeadChips (Illumina) and Nexus Expression 2.0 (BioDiscovery) as described elsewhere (22,27). Significantly regulated transcripts (hereafter referred to as regulated) were controlled for false-discovery rate according to the Benjamini–Hochberg method (28) with an adjusted P value cutoff of 0.01 and a fold-change threshold of at least 1.5. Thyroid-associated upstream regulators and diseases and functions were generated using Ingenuity Pathway Analysis (IPA; Ingenuity Systems, www.ingenuity.com) with the Fisher exact test (P < 0.05).
Transcript Expression Signatures
To discern between IR- and TH-induced responses, literature-based signature genes were compiled, and the microarray data (21–24) were analyzed with regard to significantly regulated transcripts within respective signatures. The IR-associated gene signature was composed of 56 genes adapted from Snyder and Morgan (29) and Chaudhry (30) as presented in Supplemental Table 1 (supplemental materials are available at http://jnm.snmjournals.org); the list also contains information on known RNA expression of human homologs according to the Human Protein Atlas (http://www.proteinatlas.org). For TH-associated responses, a list of 61 genes and gene groups (encoding multimeric protein) was based on the literature; Supplemental Table 2 provides a gene list, references, and known RNA expression of human homologs. Various studies on TH-induced responses have been performed on the protein level without investigating the level of transcriptional regulation. In these cases, gene regulation was inferred.
RESULTS
Transcriptional Regulation and Absorbed Dose
The dose–response studies (21,22) showed a distinctly similar pattern in total transcriptional regulation after 24 h for all investigated tissues (Fig. 1). A similar substantial decrease in transcript regulation was observed in all tissues between 0.64 and 1.8 kBq of 211At, although thyroid and nonthyroid tissues were subjected to distinctly different absorbed dose levels by a factor of approximately 102. Distinct dissimilarities between absorbed dose and number of regulated transcripts were observed between tissues. For instance, the spleen showed between 80% and 94% of the total number of transcripts regulated in the thyroid on administration of 0.064 and 0.64 kBq of 211At, respectively, while receiving only around 1% of absorbed dose to thyroid. Consecutive studies (23,24) on transcriptional regulation over time in response to medium-activity (1.7–1.8 kBq) 211At administration further indicated that the extent of transcriptional regulation was tissue-specific (Fig. 2). The liver and cortical and medullary kidney tissues received a similar absorbed dose level but showed pronounced differences in the total number of significantly regulated transcripts; moreover, the ratio of regulated transcripts between the tissues varied over time. Interestingly, these tissues received the lowest absorbed dose levels but showed stronger overall transcriptional responses (with the exception of the kidney medulla at 7 d) than the lungs and spleen. Among nonthyroid tissues, the lungs received the highest absorbed dose levels but showed a response intensity similar to the spleen across all time points. Further dissimilarities between absorbed dose and response intensity were observed in relation to time of analysis. For instance, both kidney tissues received similar absorbed dose, but after 7 d, around 100 transcripts were regulated in the kidney cortex whereas regulation in the kidney medulla was only marginal. In contrast, liver tissue received only around 1% of absorbed dose to thyroid over 1 h, but the tissue still showed 71% of thyroid response intensity at this early time point.

Absorbed dose and total transcript regulation after 24 h over 211At activity range. Tissue-specific absorbed dose (mGy) from intravenously administered 211At (0.064–42 kBq) was adapted from previously presented studies on kidney cortex and medulla, liver, lungs, and spleen (22) and thyroid (21). Number of significantly regulated transcripts includes both up- and downregulated transcripts at a given exposure.
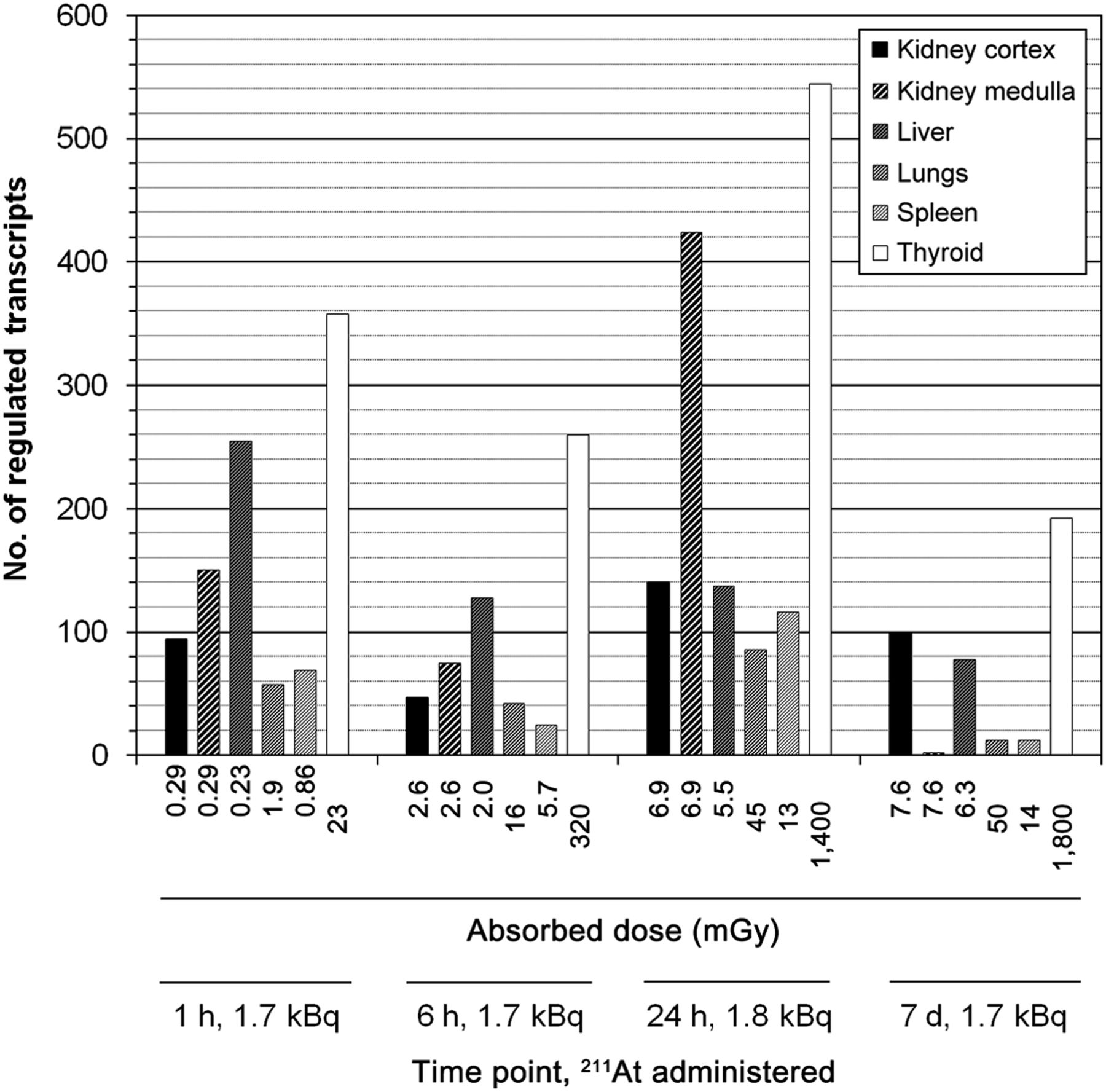
Absorbed dose and total transcript regulation over time to low 211At activity. Tissue-specific absorbed dose (mGy) from intravenously administered 211At (1.7–1.8 kBq) was adapted from previous studies on kidney cortex and medulla, liver, lungs, and spleen (23) and thyroid (24). Number of significantly regulated transcripts includes both up- and downregulated transcripts at a given exposure and time point.
Signature Gene Regulation
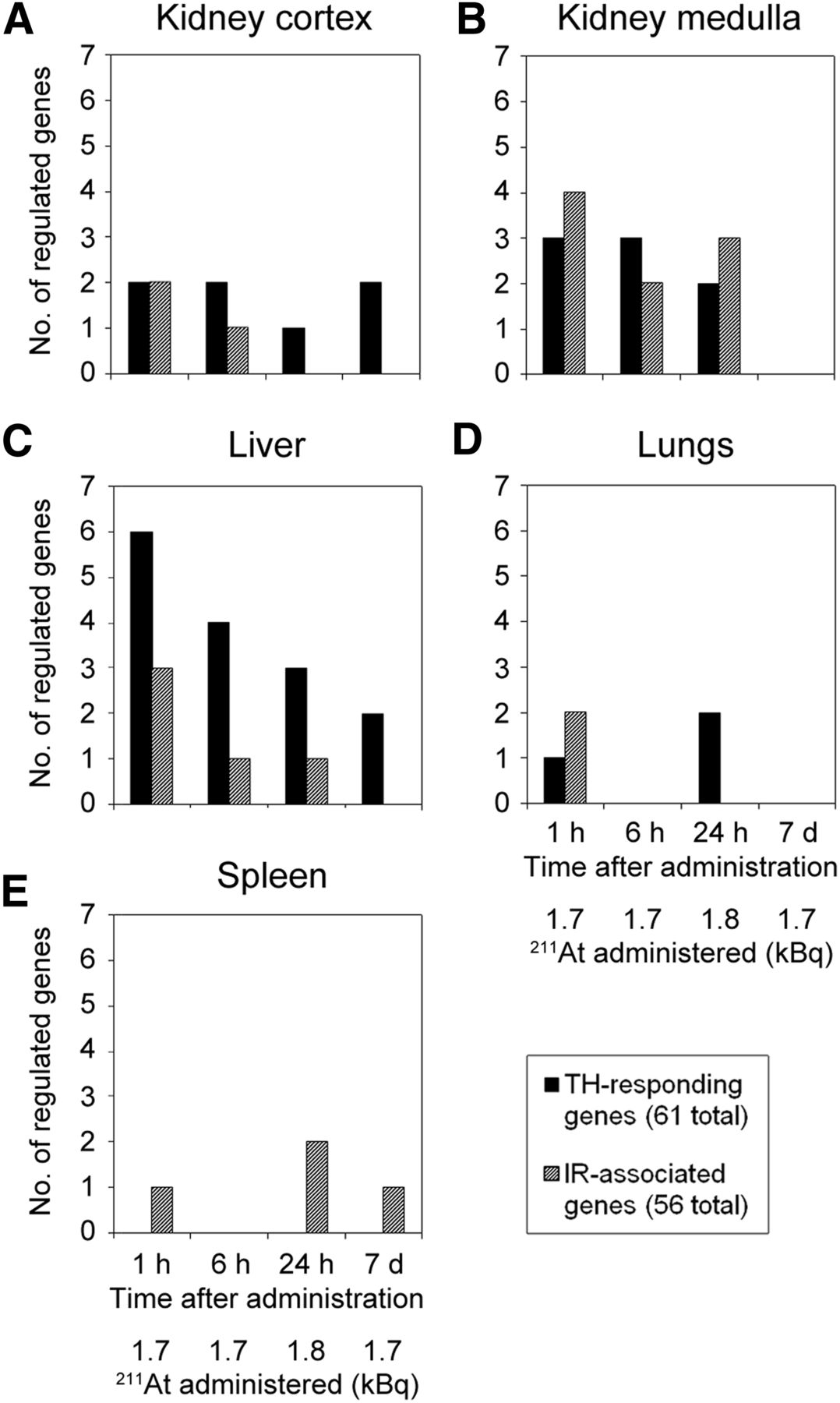
The microarray datasets were analyzed with regard to transcript up- or downregulation of genes associated with IR or responding to TH regulation (Fig. 3). The transcriptional response for either gene signature showed tissue specificity, varied over time, and supported the dissimilarity between response intensity and absorbed dose level. Detailed information on regulated gene, probe ID, and fold change for respective signatures is shown in Supplemental Tables 3 and 4. The number of regulated genes for either signature ranged between 0 and 6. In the kidney cortex (Fig. 3A), transcriptional responses to TH were observed at all time points, whereas responses to IR-associated genes were seen only after 1 and 6 h. The number of regulated TH-responding genes was higher than IR-associated genes across all time points except at 1 h, where both signatures showed regulation of 2 genes. The kidney medulla (Fig. 3B) showed a slightly increased number of regulated genes in both signatures compared with cortical tissue, however, no responses were detected for either signature after 7 d. The liver (Fig. 3C) showed a distinctly stronger response in genes associated with TH-induced regulation, which dominated over IR-associated regulation consistently across all time points. For both signatures, the regulatory incidences decreased with time and no regulation was detected in IR-associated genes after 7 d. In lung tissue (Fig. 3D), TH-responding genes were regulated after 1 h (1 gene) and after 24 h (2 genes), whereas IR-associated genes were regulated only after 1 h (2 genes). The spleen (Fig. 3E) showed no response in TH-responding genes at any investigated time point and exhibited regulatory responses only to IR-associated genes—that is, after 1 h (1 gene), 24 h (2 genes), and 7 d (1 gene). Interestingly, lung tissue showed the lowest incidence in IR-associated genes, although it received highest absorbed dose among nonthyroid tissues.

Comparison of signature gene regulation in kidney cortex and medulla, liver, lungs, and spleen. Number (no.) of significantly regulated genes at various time points after low-activity 211At administration is shown for genes associated with regulation induced by IR (hashed) and TH (black). Responses are shown for kidney cortex (A), kidney medulla (B), liver (C), lungs (D), and spleen (E). Analyses were performed on microarray data GEO:GSE40806 and GEO:GSE56894 first reported in Langen et al. (22), and Langen et al. (23), respectively. Supplemental Tables 3 and 4 show regulated genes, transcript identification number, and fold change values of IR-associated and TH-responding signature genes, respectively.
Thyroid- and TH-Associated Pathways and Regulators
Pathway analysis predicted regulation of several TH-associated upstream regulators in all tissues, but the incidence varied between tissues and time points (Table 1). The liver and kidney cortex showed the highest incidence with 3–4 and 1–3 significantly detected upstream regulators per time point, respectively. In the kidney medulla, lungs, and spleen, upstream regulators were identified only at intermittent time points. L-triiodothyronine exhibited the highest overall response in the cohort, showing a trend toward activation at early time points and a trend toward inhibition after 7 d in the kidney cortex and liver. L-triiodothyronine was further detected in the lungs after 24 h and in spleen after 1 h, but not in the kidney medulla. Further identified upstream regulators were TH, TH receptor, and triiodothyronine (reverse). Most predicted TH-associated biologic functions and diseases were related to endocrine system disorders and thyroid cancer (carcinoma) (Tables 2 and 3). The incidence of TH-associated diseases and functions generally showed different trends in time response compared with upstream regulators. In the kidney cortex and liver, upstream regulators were continuously detected, whereas responses in disorders and functions were identified only after 6 h, or after 24 h and 7 d, respectively. The liver showed by far the highest incidence with 15 detected disorders or functions (Table 3), which was in agreement with showing the highest incidence of upstream regulators. Interestingly, detection incidence was distinctly higher in the kidney medulla and spleen compared with upstream regulators, whereas the lungs did not show any responses regarding diseases and functions.
TH-Associated Upstream Regulators in Nonthyroid Tissues
Thyroid-Associated Diseases and Functions in Kidneys, Lungs, and Spleen
Thyroid-Associated Diseases and Functions in Liver
DISCUSSION
Regulation of gene expression in response to a stressor is a multistage process. Aside from epigenetic factors and transcriptional regulation, transcript expression levels represent the first stage of gene expression, which in turn regulates cellular responses. High-throughput technologies such as expression microarrays allow analysis of genome-wide transcriptional regulation without restricting the analytic frame to a limited set of genes or pathways. This approach is particularly beneficial for an in vivo setting with differential dose exposure, in which the analytic frame must cover a multifactorial and tissue-specific context. In the case of free 211At, the regulatory thyroid gland is dominantly affected by IR exposure due to specifically high uptake. Gene regulation in the thyroid gland in response to 211At has been analyzed in detail in related studies (21,24). Other tissues that underlie thyroid-dependent metabolic regulation also exhibit 211At uptake, but at a much lower level. Additional information on 211At characterization and considerations for biodistribution and absorbed dose are provided in the supplemental materials.
Interorgan signaling from the thyroid gland to target tissues introduces systemically induced gene regulation in addition to IR-induced gene regulation in each tissue. In theory, systemic effects originating from the thyroid gland could be caused by changes in synthesis or secretion of physiologic signaling factors, that is, TH or calcitonin, or by IR-induced long-range nontargeted signaling that would be mediated by other factors. Because knowledge of specific target genes for bystander effects is still scarce, especially in the in vivo context, the type or mechanism of long-range bystander signaling could not be discerned in this setup. Instead, the aim of the study was to demonstrate the regulation of TH-responding genes and thyroid-related pathway regulation in nonthyroid tissues. The systemic physiologic context, however, does not exclude the possible contribution of strictly IR-associated long-range nontargeted effects in this setting. The analysis of signature gene responses in nonthyroid tissues supported the hypothesis (22) that transcriptional regulation in these tissues was not solely due to IR exposure, but in part due to thyroid-dependent signaling. TH-related responses in nonthyroid tissues were also demonstrated using pathway analysis. The relative contribution to the overall transcriptional regulation, however, could not be inferred from this type of data. It should further be noted that in the literature, Ccnd1, Cdkn1a, Fos, Mapk1, and Mdm2 are associated with both IR- and TH-induced gene regulation and were used for both signatures, accordingly. It is difficult to discern if these genes were regulated in response to one or both inducers in this setting.
Irrespective of the underlying mechanisms, systemic effects appear to have a strong impact on normal tissue gene regulation, which ultimately can influence tissue health status. The available data suggest that the quality and extent of systemic effects is dependent on various physical and biologic factors concerning not only the type of exposure, but also what tissue is irradiated. Clinical practice, however, does not consider systemic effects in normal tissue risk assessment. There is a need for basic research and clinical investigations to elucidate the molecular mechanisms that mediate these effects and to understand the parameters that lead to detrimental, beneficial, or insignificant systemic effects. Once these mechanisms are understood on the physical and biologic levels, clinicians will be able to estimate the extent and quality of systemic effects in diagnostic and treatment planning and can give tissue-specific prophylactic countermeasures, if needed.
In general, tissue specificity was seen regarding continuous versus intermittent regulation, strength of regulation, and number of incidence. Although trends in activation or inhibition were observed, respective z scores did not exceed the threshold of ±2.0 for statistical significance. The liver showed the highest incidence of response regarding both upstream regulators and identified diseases and functions, which—taken together with its strong response in signature gene analysis—indicated that it may be a major target tissue of thyroid-dependent regulation among the investigated tissues. In the kidney cortex and liver, a general time-dependent trend was observed for L-triiodothyronine (T3) with activation at early time points after 211At administration and inhibition after 1 wk. In contrast, the TH receptor as an upstream regulator was inferred only at 1 h in either tissue. TH receptors are nuclear receptors that function as transcription factors after binding to TH; hence, the functional outcome regarding transcript expression is dependent on up- or downregulation of both hormone and receptor. Interestingly, several diseases and functions were identified in all tissues (except lungs) that were not only related to TH, but also to thyroid tissue–specific disorders and processes. These identified pathways could comprise target genes of normal TH-dependent regulation that are associated with thyroid-specific diseases in the database but are not biologically related to disorders in nonthyroid tissues. It is also possible that these responses indicate nontargeted effects that are reminiscent of IR-induced effects in thyroid tissue.
Another unknown factor is tissue-specific response over time, because tissues are categorized as early or late responding with regard to IR-induced responses (31). As such, responses in a tissue may be best related to the tissue-specific time of onset and not to a certain time point after administration. This might also be valid for TH-induced responses, but to the best of our knowledge, the temporal context of this systemic regulation has not been established on the transcriptional level in the target tissues of interest.
CONCLUSION
This consecutive study on transcriptomic regulation after intravenous 211At administration supported the previously postulated hypothesis that transcriptional responses in the liver, lungs, spleen, and kidney tissues are influenced—in part—by thyroid-dependent effects. Although it was not possible to demonstrate direct causality in the given setup, TH-associated signature genes were regulated more frequently than IR-associated signature genes. The strength of response for either signature appeared to be tissue-specific. Furthermore, upstream regulators related to TH and diseases and functions related to thyroid (hormones) were detected in nonthyroid tissues using Ingenuity Pathway Analysis; moreover, tissue-specific variation in response intensity as well as time of response were also observed. These findings demonstrate the complexity of in vivo responses and the need to consider organs in a systemic context when studying normal tissue responses to systemically administered radionuclides.
DISCLOSURE
We thank the following organizations for financial support of this study: the European Commission FP7 Collaborative Project TARCC HEALTH-F2-2007-201962, the Swedish Research Council (grant no. 21073), the Swedish Cancer Society (grant no. 3427), BioCARE—a National Strategic Research Program at the University of Gothenburg, the Swedish Radiation Safety Authority, the King Gustav V Jubilee Clinic Cancer Research Foundation, the Sahlgrenska University Hospital Research Funds, the Assar Gabrielsson Cancer Research Foundation, the Adlerbertska Research Fund, and the Wilhelm och Martina Lundgren science trust fund. No other potential conflict of interest relevant to this article was reported.
Footnotes
Published online Oct. 20, 2016.
- © 2017 by the Society of Nuclear Medicine and Molecular Imaging.
REFERENCES
- Received for publication April 15, 2016.
- Accepted for publication September 8, 2016.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.