


Abstract
We recently developed a novel PET probe, 6-11C-methyl-m-tyrosine (11C-6MemTyr), for quantitative imaging of presynaptic dopamine synthesis in the living brain. In the present study, 11C-6MemTyr was compared with β-11C-l-DOPA and 6-18F-fluoro-l-dopa (18F-FDOPA) in the brains of normal and Parkinson disease (PD) model monkeys (Macaca fascicularis). Methods: PD model monkeys were prepared by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) administration, and 11C-β-CFT was applied to assess neuronal damage as dopamine transporter (DAT) availability. 11C-6MemTyr, β-11C-l-DOPA, or 18F-FDOPA was injected with and without carbidopa, a specific inhibitor of peripheral aromatic L-amino acid decarboxylase. In normal and PD monkeys, the dopamine synthesis rates calculated using PET probes were analyzed by the correlation plot with DAT availability in the striatum. Results: In normal monkeys, whole-brain uptake of β-11C-l-DOPA and 18F-FDOPA were significantly increased by carbidopa at the clinical dose of 5 mg/kg by mouth. In contrast, 11C-6MemTyr was not affected by carbidopa at this dose, and the metabolic constant value of 11C-6MemTyr in the striatum was significantly higher than those of the other 2 PET probes. Significant reduction of the presynaptic DAT availability in the striatum was detected in MPTP monkeys, and correlation analyses demonstrated that 11C-6MemTyr could detect dopaminergic damage in the striatum with much more sensitivity than the other PET probes. Conclusion: 11C-6MemTyr is a potential PET probe for quantitative imaging of presynaptic dopamine activity in the living brain with PET.
Parkinson disease (PD) provides progressive degradation of nigrostriatal pathways with selective loss of the dopaminergic neurons in the substantia nigra pars compacta (SNc), resulting in movement disorders such as resting tremor, akinesia, bradykinesia, rigidity, and postural instability. These clinical symptoms of PD were induced after 40%–50% loss of the dopaminergic neurons in the SNc and the reduction of dopamine in the striatum to about 20% of normal levels (1). Although the direct cause of this selective neurodegeneration in PD is not clearly understood, mitochondrial dysfunction has emerged as a common aspect of the pathogenesis (2,3).
Brain abnormality of PD patients has been imaged with PET as impaired presynaptic dopamine parameters such as dopamine synthesis, dopamine transporter (DAT), and vesicular monoamine transporter availabilities, as well as tentatively upregulated postsynaptic dopamine D2 receptor binding at the initial stage (4). Dopamine synthesis capacity is regulated by DOPA concentration. PD syndromes are ameliorated by l-DOPA administration, suggesting that lowered dopamine neuronal activity can be overcome by a higher level of l-DOPA, Our previous study using l-β-11C-DOPA (5) demonstrated that 6R-L-erythro-5,6,7,8-tetrahydrobiopterin, a cofactor for tyrosine hydroxylase (TH), could increase the striatal dopamine synthesis in normal monkey brain (6). 6-18F-fluoro-l-dopa (18F-FDOPA) has widely been applied for clinical diagnosis of the presynaptic dopamine function using PET. It has been assumed that 18F-FDOPA follows the same transport, synthetic, and metabolic pathways of dopamine (7). 18F-FDOPA uptake reflects the activity of aromatic L-amino acid decarboxylase (AADC) in the nerve terminals of the nigrostriatal dopaminergic pathway, and the metabolized 18F-fruoro-dopamine (18F-FDA) is stored in the vesicles through vesicular monoamine transporter. We have applied β-11C-l-DOPA (5) to monitor dopamine synthesis rate facilitated by pharmacologic manipulations in normal monkeys (6,8,9) as well as impaired (10) in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)–treated PD model monkeys (11). Because these research studies required multiparametric PET assessments of presynaptic dopamine synthesis, DAT, and postsynaptic dopamine receptors in the same subject in a day, the short half-life of 11C is very useful for the repeated PET measurements. In contrast, the labeling by multienzymatic reactions limits the availability of β-11C-l-DOPA only in the limited PET facilities (5).
Despite the usefulness of 18F-FDOPA PET for the diagnosis of PD patients (4,12), 18F-FDOPA is not always an ideal PET probe. One reason is that because 18F-FDA is taken into the vesicles in presynaptic dopaminergic terminals, released, and metabolized further, its signal is influenced by both vesicular uptake and release processes (13). Another reason is caused by the peripheral decarboxylation by AADC and the methylation at the 3-hydroxyl group by catechol O-methyl transferase (COMT). The resulting 18F-3-O-Me-FDOPA crosses the blood–brain barrier by the large neutral amino acid transporter (14), thereby worsening the signal-to-noise ratio and complicating kinetic modeling (15,16). A further limitation is due to the radiolabeling production of 18F-FDOPA with electrophilic reactions using molecular 18F-F2 or 18F-acetylhypofuorite (17) and nucleophilic reactions with no-carrier-added 18F-fluoride (18,19). The former reactions provide the low radioactivity yields with low specific radioactivity, and the latter ones are concerned with the complex multistep reactions and enantiomeric purity.
6-18F-fluoro-L-m-tyrosine (18F-FmT) has been proposed for presynaptic dopamine imaging (20–23). 18F-FmT is decarboxylated to 6-18F-fluoro-m-tyramine (18F-FmTA) by AADC with a 10-fold-greater affinity than 18F-FDOPA (21). 18F-FmTA has a 3-fold-greater affinity for monoamine oxidase-A than 18F-FDA, and 6-18F-fluoro-3-hydroxyphenylacetic acid (18F-FHPAA) is trapped in the striatum (21). 18F-FmTA and 18F-FHPAA were also produced by peripheral AADC and monoamine oxidase-A, respectively, in plasma; however, these metabolites of 18F-FmT cannot cross the blood–brain barrier (24). Methylated metabolites of 18F-FmT are not formed because 18F-FmT does not have the catechol structure for a substrate of COMT. These metabolic properties can make the signal of 18F-FmT fully representing the extent of AADC activity with better quality than l-DOPA–based PET probes. However, the long half-life of 18F prevents repeated PET measurements of pre- and postdrug treatments in the same subject in a day.
We developed a 11C-labeled PET probe, 6-11C-methyl-m-tyrosine (11C-6MemTyr), for quantification of presynaptic dopamine activity (25). In the present study, 11C-6MemTyr was compared with β-11C-l-DOPA and 18F-FDOPA in the monkey brain (Macaca fascicularis). The effects of carbidopa, a peripheral AADC inhibitor, on the brain uptake of these PET probes were assessed in normal animals. Furthermore, in PD animals, dopaminergic neuronal damage induced by MPTP was determined as DAT availability measured using 11C-β-CFT (26), and sensitivity of each PET probe to dopaminergic neuronal damage was evaluated by correlation plot analyses with DAT availability.
MATERIALS AND METHODS
Animals and Drugs
Animals were maintained and handled in accordance with the recommendations of the U.S. National Institutes of Health and the guidelines of the Central Research Laboratory, Hamamatsu Photonics. This experiment was approved by the Ethical Committee of the Central Research Laboratory, Hamamatsu Photonics.
MR images of the monkeys were obtained with a 3.0-T MR imager (Signa Excite HDxt 3.0T; GE Healthcare) using a 3-dimensional spoiled gradient echo sequence (176 slices with a 256 × 256 image matrix; slice thickness/spacing, 1.4/0.7 mm; echo time, 3.4–3.6 ms; repetition time, 7.7–8.0 ms; inversion time, 400 ms; and flip angle, 15°) under pentobarbital anesthesia.
Isoflurane and pancronium were purchased from Sumitomo Dainippon Pharma Co. Ltd. and Daiichi Sankyo Co. Ltd., respectively. Kryptofix222 (K[2,2,2]), N-(diphenylmethylene)glycine tert-butyl ester, and MPTP were from Sigma-Aldrich Japan. The enzymes for β-11C-l-DOPA synthesis, alanine racemase (EC 5.1.1.1.), D-amino acid oxidase (EC1.4.3.3.), and β-tyrosinase (EC 4.1.99.2.) were purchased from lkeda Food Research. 6-nitoroveratraldehyde and (R,R)-3,4,5-trifluorophenyl-NAS bromide were obtained from Tokyo Chemical Industry Co. Ltd. and Wako Pure Chemical Industries Ltd., respectively. Fluoro-dopamine (FDA) was from ABX GmbH. Precursors of 11C-6MemTyr, 18F-FDOPA, and 11C-β-CFT; their corresponding standard compounds; and 6-Me-m-tyramine (6MemTA), 6-Me-m-hydroxyphenylacetic acid (6MHPAA), and 3-O-Me-FDOPA were purchased from NARD Institute Ltd.
PET Ligand Syntheses
Positron-emitting 11C and 18F were produced by 14N(p,α)11C and 18O(p, n)18F nuclear reactions, respectively, using the cyclotron (HM-18; Sumitomo Heavy Industry, Ltd.) at the Hamamatsu Photonics PET center.
11C-6MemTyr was labeled using a rapid Pd(0)-mediated cross-coupling reaction of 11C-methyl iodide and corresponding alkenyl boronate precursor (25). The synthesis time was 35 min from an end of bombardment. The radiochemical purity was better than 99%, and the specific radioactivity was 40.0 ± 10.1 GBq/μmol.
β-11C-l-DOPA was synthesized using a combination of organic synthesis and multienzymatic procedures (5). The radiochemical purity was better than 98%, and the specific radioactivity was 25.4 ± 8.5 GBq/μmol.
18F-FDOPA was labeled by a highly enantioselective synthesis method using chiral phase-transfer alkylation (19). The radiochemical purity was greater than 99%, and the specific radioactivity was 47.0 ± 13.9 GBq/μmol.
11C-β-CFT for DAT was labeled by 11C-N-methylation of its nor-compound with 11C-methyl iodide (26). The radiochemical purity was greater than 98%, and the specific radioactivity was 42.0 ± 7.9 GBq/μmol.
PD Monkey Model
Cynomolgus monkeys weighing 2.0–3.5 kg were used for PD model preparation (10,11). MPTP at doses ranging from 0.2 to 0.4 mg/kg of free base in phosphate-buffered saline was injected intravenously once per week over a 4-mo period until a stable parkinsonian syndrome was observed. The total dose of MPTP administered was 1.5 and 3.0 mg/kg.
PET Measurements
Four normal and 4 MPTP monkeys underwent PET scans using 11C-6MemTyr, β-11C-L-DOPA, and 18F-FDOPA, all of which were obtained with and without carbidopa (5 mg/kg, by mouth, 1 h before probe injection), and 11C-β-CFT under isoflurane anesthetic state. For more comparison of carbidopa effects, 4 normal monkeys were analyzed using 11C-6MemTyr and 18F-FDOPA after carbidopa administrations at 0.15 and 5 mg/kg, intravenously, 1 h before probe injection. After overnight fasting, monkeys were tracheostomized, immobilized with 0.05 mg/kg intramuscularly of pancronium bromide every 2 h, and artificially ventilated (Cato; Dräger). Anesthesia was continued with 0.8 volume % isoflurane in a N2O/O2/N2 (1:1:1) gas mixture during the entire experiment. A venous cannula for PET ligand injection was inserted. The mean arterial blood pressure, heart rate, rectal temperature, arterial PO2 and PCO2, and pH were continuously or regularly monitored. The animal’s body temperature was maintained within normal limits with a heating blanket. The animal was placed supine with a head holder in the PET gantry with stereotactic coordinates aligned parallel to the OM plane. After a transmission scan for 30 min, emission scans were obtained for 91 min using an animal PET scanner (SHR-7700; Hamamatsu Photonics) (27). The PET data were reconstructed by the filtered backprojection method with a Hanning filter of 4.5 mm in full width at half maximum and attenuation correction using the transmission scan data.
Volumes of interest (VOIs) in brain regions were drawn manually on the MR image according to regional information from BrainMaps.org (28), and VOIs of MR images were superimposed on the coregistered PET images to measure the time–activity curves of each PET probe for kinetic analyses.
PET Data Analysis
The quantitative analyses of 11C-6MemTyr, β-11C-l-DOPA, and 18F-FDOPA were performed by a multitime graphical analysis (MTGA) (29–31) using PMOD software (PMOD Technologies Ltd.) applying the time–activity curve in the cerebellum as an input function.
The analysis of 11C-β-CFT was conducted by nondisplaceable binding potential (BPND) analysis (32) using PMOD software. As an input function, the time–activity curve in the cerebellum was applied.
Metabolic Analyses
A radio–thin-layer chromatography method was used to evaluate the effects of carbidopa on the metabolic profiles of 11C-6MemTyr and 18F-FDOPA in plasma. Arterial blood samples were obtained from monkeys at 5, 15, 30, 60, and 90 min after the injection of each PET probe and centrifuged at 12,000 rpm for 3 min to separate plasma. Ethanol was added to the plasma (plasma/ethanol = 1:2 (v/v)) followed by centrifugation. The obtained supernatants were developed with CHCl3/n-BuOH/CH3COOH/H2O (2:3:1:1) for 11C-6MemTyr or n-BuOH/CH3COOH/H2O (4:1:1) for 18F-FDOPA, respectively, and exposed to a phosphoimaging plate. The plate was measured with FLA-7000 (Fujifilm Corp.).
Immunohistochemical Assessment
Monkeys were perfused with 4% paraformaldehyde through the ascending aorta under anesthesia with an overdose sodium pentobarbital. The brains were removed and cut into blocks 5 mm thick. These blocks were postfixed, left for 3 d in phosphate-buffered saline containing 30% sucrose, and cut into coronal sections 30 μm thick. Sections were treated with 0.3% H2O2 for 15 min to inhibit endogenous peroxidase. Sections were incubated at 48°C for 2 d in phosphate-buffered saline containing 0.3% Triton X-100 and primary antibodies against mouse monoclonal anti-TH antibody (1:8,000). Sections were incubated with biotinylated antimouse IgG (1:1,000) for 1 h at room temperature and in avidin–biotin–peroxidase complex (1:50) for 30 min at room temperature. Peroxidase activity was revealed in 50 mM Tris-HCl buffer (pH 7.6) containing 0.0004% H2O2 and 0.01% diaminobenzidine.
Statistical Analysis
Results are expressed as mean ± SD. Comparisons between conditions were performed using the paired, 2-tailed Student t test. A probability level of less than 5% (P < 0.05) was considered to indicate statistical significance.
RESULTS
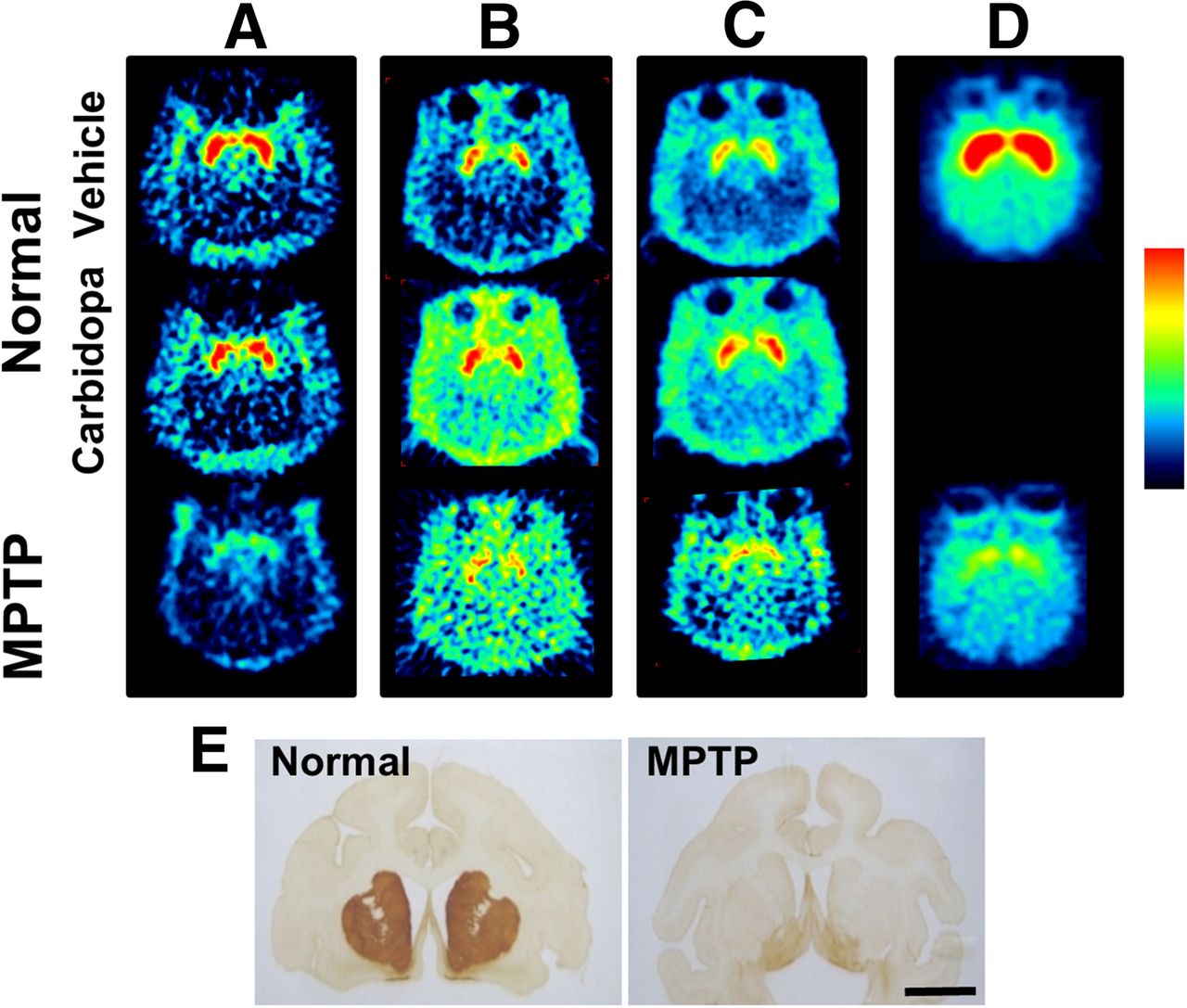
As shown in the uppermost panels of Figures 1A–1C, each SUV image of 11C-6MemTyr (Fig. 1A), β-11C-l-DOPA (Fig. 1B), and 18F-FDOPA (Fig. 1C) clearly showed the presynaptic dopamine synthesis activity in the striatum of normal monkey brain, and PET images of 11C-6MemTyr provided high SUV in the striatum with best contrast between thestriatum and cortical regions, even without carbidopa administration. After the preadministration of carbidopa at 5 mg/kg by mouth shown in the middle panels of Figures 1A–1C, the uptake of β-11C-l-DOPA (Fig. 1B) and 18F-FDOPA (Fig. 1C) was increased not only in the striatal and cortical regions but also outside the skull. In contrast, no uptake changes in 11C-6MemTyr were observed in the cortical regions by carbidopa (Fig. 1A).

Typical PET images of 11C-6MemTyr (A), β-11C-l-DOPA (B), 18F-FDOPA (C), and 11C-β-CFT (D) in brains of normal and MPTP-treated monkeys. For PET imaging from A to D, vehicle or carbidopa at clinical dose of 5 mg/kg by mouth was administered. PET scans were conducted for 91 min with each PET probe, and SUV images from 62 to 91 min after injection were reconstructed from A to E. Immunohistologic analyses were performed using anti-TH antibody and brain slices of normal and MPTP-treated monkeys (E).
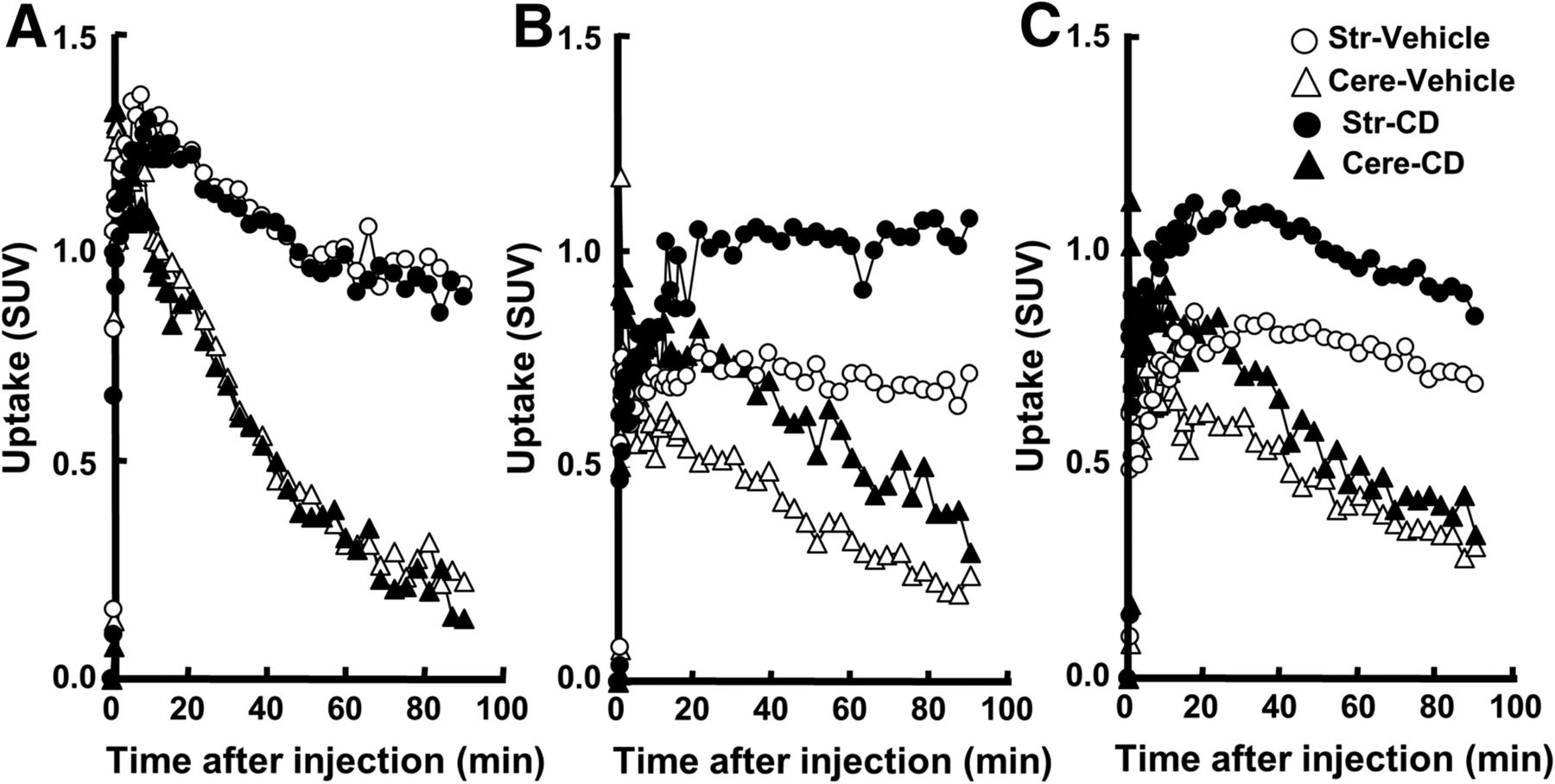
Figure 2 shows time–activity curves of 11C-6MemTyr (Fig. 2A), β-11C-l-DOPA (Fig. 2B), and 18F-FDOPA (Fig. 2C) in the striatal and cerebellar regions of normal monkey brain with and without carbidopa. Without carbidopa, time–activity curves of 11C-6MemTyr reached a peak within 10 min in the striatum and within 5 min in the cerebellum after the injection, followed by the gradual clearance from the striatum and rapid from the cerebellum (Fig. 2A). Time–activity curves of β-11C-l-DOPA (Fig. 2B) and 18F-FDOPA (Fig. 2C) showed much lower peak values with much slower kinetics in the monkey brain than those of 11C-6MemTyr (Fig. 2A). The preadministration of carbidopa at 5 mg/kg by mouth resulted in the higher peak values as well as slower kinetics of β-11C-l-DOPA (Fig. 2B) and 18F-FDOPA (Fig. 2C) than those in control. In contrast, the kinetics of 11C-6MemTyr in the striatum and cerebellum were not affected by carbidopa (Fig. 2A).

Effects of carbidopa (CD) at 5 mg/kg by mouth on time–activity curves of 11C-6MemTyr (A), β-11C-l-DOPA (B), and 18F-FDOPA (C) in normal monkey brains. PET scans were conducted for 91 min with each PET probe. SUV images from 62 to 91 min after injection were reconstructed from A to C, and VOIs of striatum and cerebellum were set on PET images aided by MRI to obtain time–activity curves of each PET probe. Uptake values in each VOI were converted to SUVs.
As shown in Figure 3, MTGA demonstrated that 11C-6MemTyr (Fig. 3A), β-11C-l-DOPA (Fig. 3B), and 18F-FDOPA (Fig. 3C) provided linear regression curves in the striatum. The slopes were highest for 11C-6MemTyr, middle for β-11C-L-DOPA, and lowest for 18F-FDOPA without carbidopa, and those of β-11C-L-DOPA and 18F-FDOPA were shifted by carbidopa.
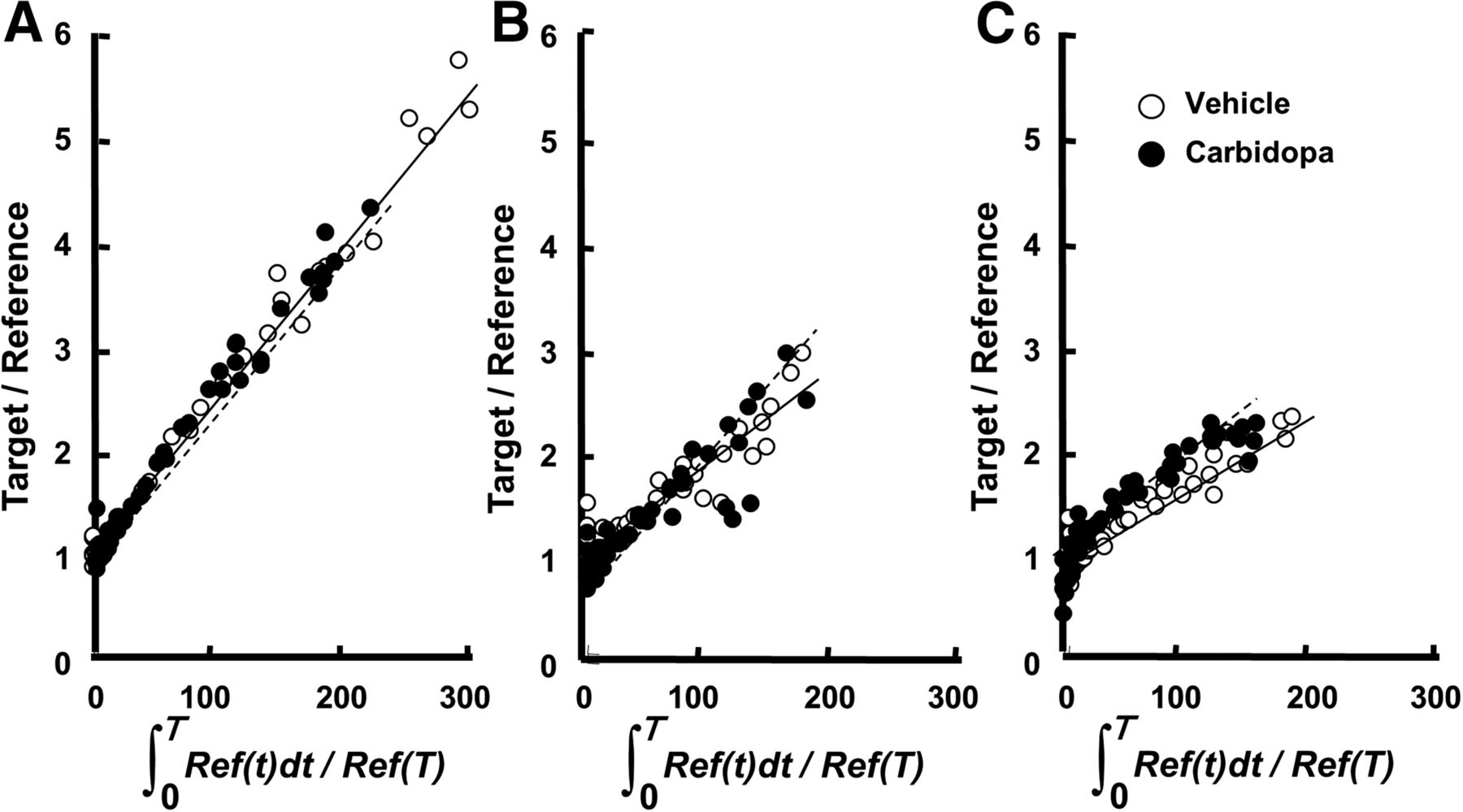
MTGA of 11C-6MemTyr (A), β-11C-l-DOPA (B), and 18F-FDOPA (C) in normal monkey brains. PET scans were conducted for 91 min with each PET probe, and time–activity curves of SUVs in striatum and cerebellum were obtained as done in Figure 3.
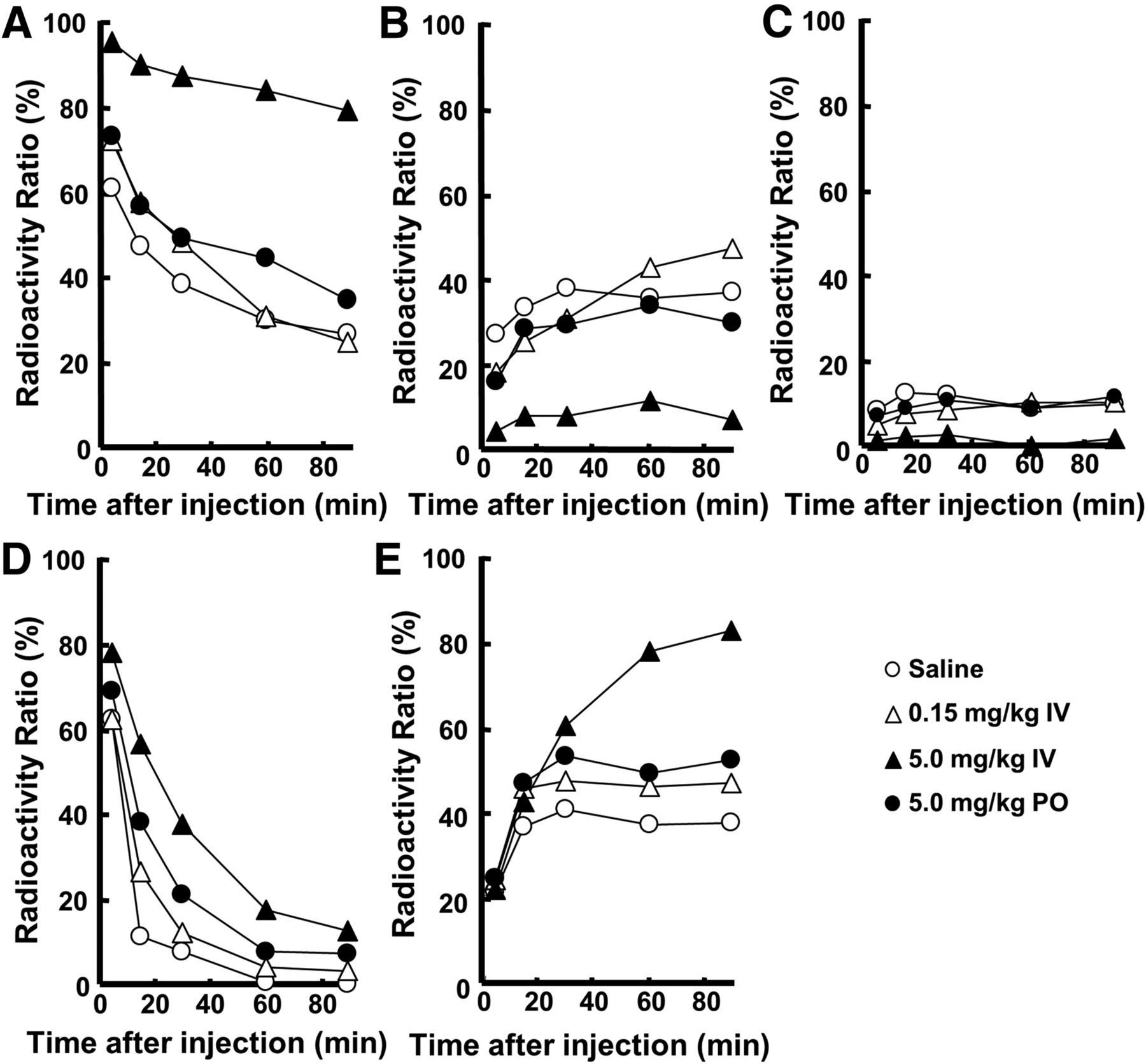
As shown in Figure 4, metabolic analyses of 11C-6MemTyr and 18F-FDOPA in plasma without carbidopa indicated that the proportion of unmetabolized 18F-FDOPA to total radioactivity (Fig. 4D) was markedly lower than that of 11C-6MemTyr (Fig. 4A) 15 min and thereafter. The preadministration of carbidopa increased this proportion of unmetabolized 18F-FDOPA (Fig. 4D) and also increased that of metabolized 18F-3-O-Me-FDOPA (Fig. 4E) by not only intravenous but also mouth administrations. The metabolic profile of 11C-6MemTyr was affected by the intravenous administration of carbidopa, whereas the alteration degrees with by-mouth administration in the ratios of unmetabolized 11C-6MemTyr (Fig. 4A), metabolized 11C-6MemTA (Fig. 4B), and 11C-6MHPAA (Fig. 4C) were much lower than those of 18F-FDOPA.
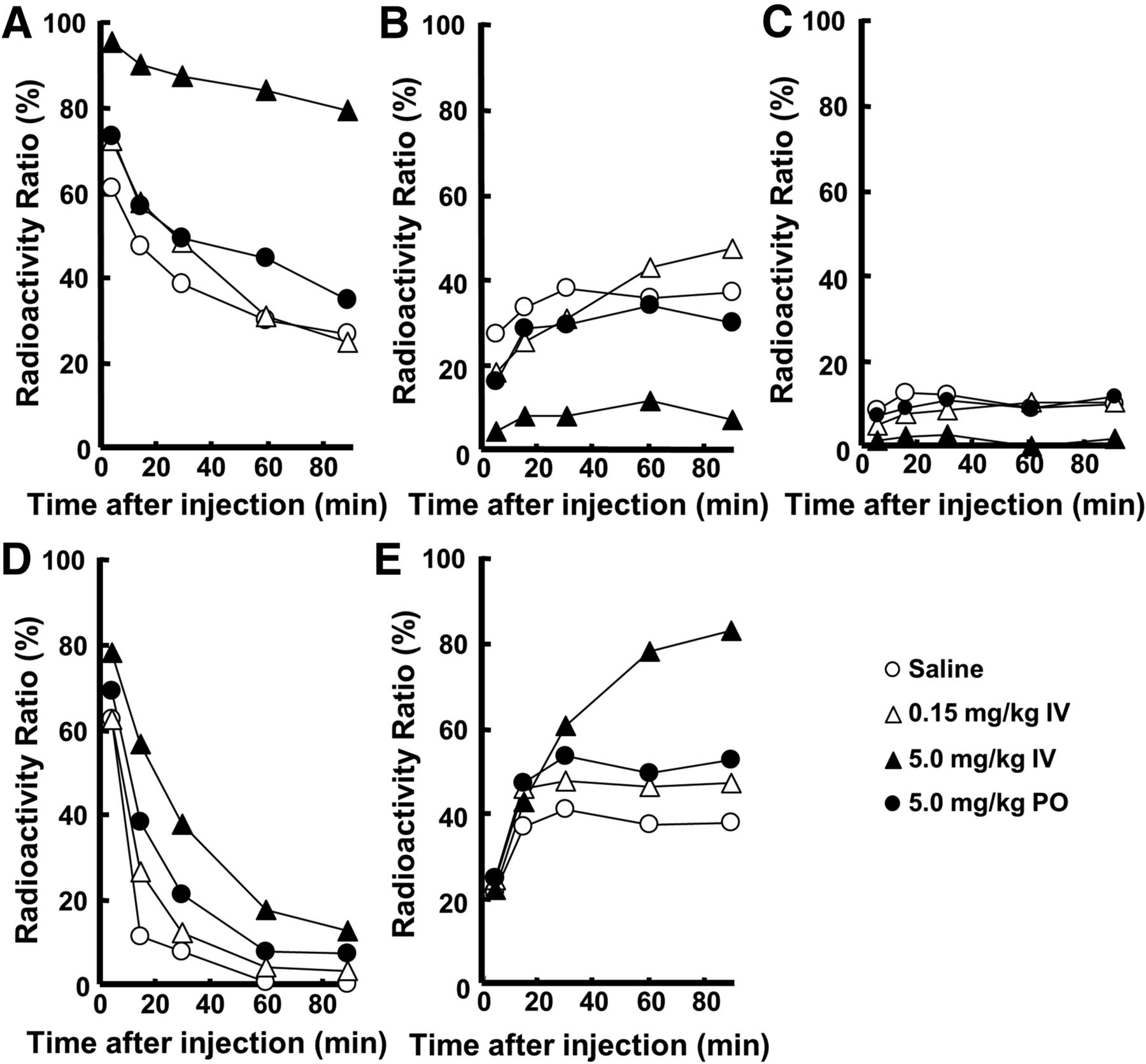
Effects of intravenous administration of carbidopa on metabolic profiles of 11C-6MemTyr (A–C) and 18F-FDOPA (D and E), in normal monkey plasma. Arterial blood samples were obtained from monkeys, and obtained plasma supernatants were applied on radio–thin-layer chromatography to analyze metabolic profiles of 11C-6MemTyr (A), 11C-6MemTA (B), 11C-6MHPAA (C), 18F-FDOPA (D), and 18F-3-O-Me-FDOPA (E). IV = intravenously; PO = by mouth.
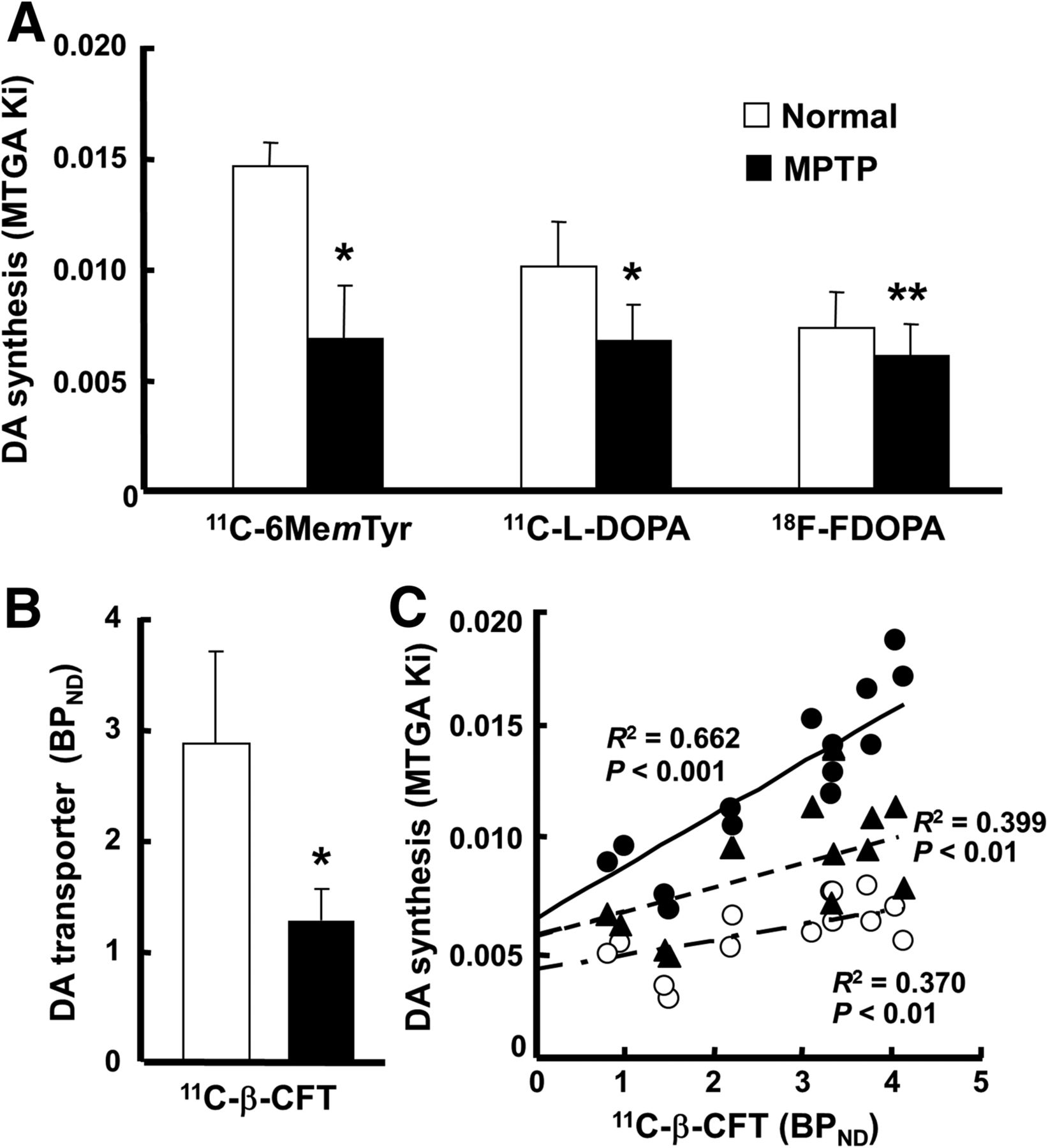
In terms of MTGA metabolic constant (Ki) values of the striatum calculated using a time–activity curve in the cerebellum as an input function, 11C-6MemTyr was the highest, β-11C-l-DOPA was middle, and 18F-FDOPA was the lowest in normal animals (Fig. 5A). The SUV images (Fig. 1) and MTGA Ki values (Fig. 5A) revealed significant reduction of a presynaptic parameter of dopamine synthesis in the striatum of MPTP monkeys using 11C-6MemTyr (Ki = 0.015 min−1 for normal and 0.007 min−1 for MPTP), β-11C-l-DOPA (0.010 and 0.007 min−1, respectively), and 18F-FDOPA (0.007 and 0.006 min−1, respectively). The most marked reduction by MPTP-induced dopamine damage was detected by 11C-6MemTyr, namely, 46.6% of that of normal animals (Fig. 5A). The levels of Ki values of MPTP-treated monkeys were 58.2%, 66.7%, and 83.1% of their corresponding normal animals, for β-11C-l-DOPA, and for 18F-FDOPA, respectively (Fig. 5A).
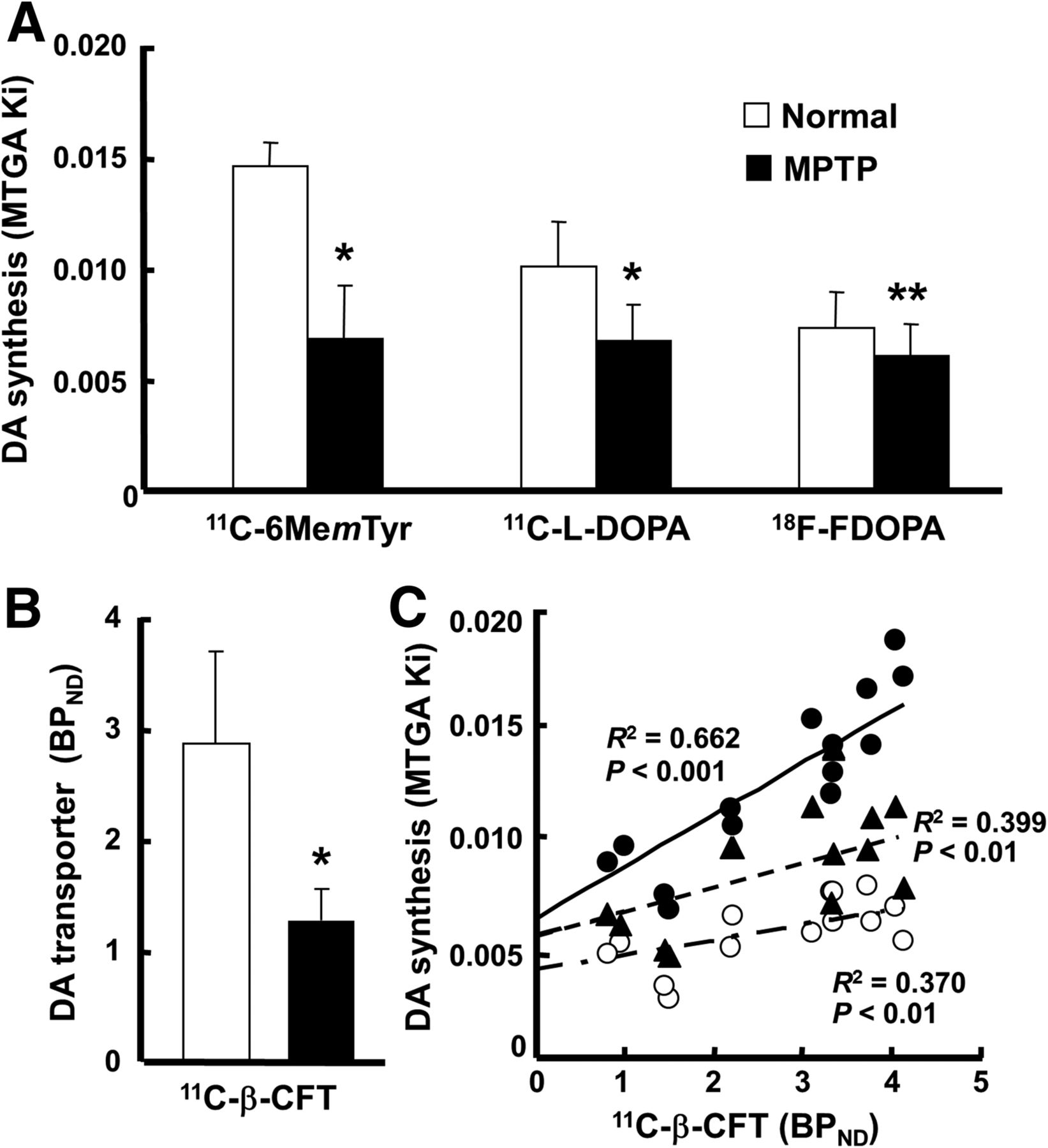
Effects of MPTP treatment on MTGA Ki values of 11C-6MemTyr, β-11C-l-DOPA, and 18F-FDOPA (A); BPND values of 11C-β-CFT (B); and correlations of MTGA Ki values of 11C-6MemTyr, β-11C-l-DOPA, or 18F-FDOPA against BPND of 11C-β-CFT (C) in monkey brains. BPND and MTGA Ki values were calculated using each time–activity curve in cerebellum as input function. ● = 11C-6MemTyr; ▲ = β-11C-L-DOPA; ○ = 18F-FDOPA. *P < 0.01 vs. each normal condition. **P < 0.05 vs. each normal condition.
When a presynaptic parameter of DAT availability was assessed using BPND of 11C-β-CFT, it remarkably decreased in the striatum of MPTP monkeys (44.7% of that of normal animals) (Fig. 5B), which was supported by immunohistologic assessment using anti-TH antibody (Fig. 1E). Although plotting of the striatal MTGA Ki values of 11C-6MemTyr, β-11C-l-DOPA, or 18F-FDOPA against BPND of 11C-β-CFT in the striatum of normal and MPTP monkeys demonstrated significant positive correlations, Ki values of 11C-6MemTyr provided greater correlation coefficients with BPND of 11C-β-CFT than any other PET probes evaluated in the present study (Fig. 5C).
DISCUSSION
When β-11C-l-DOPA or 18F-FDOPA was injected into monkeys, the preadministration of carbidopa, at a clinical dose of 5 mg/kg by mouth, increased the uptake of radioactivity into the whole brain. As typically demonstrated using 18F-FDOPA, this increased brain uptake was induced by the increase in brain-available radiolabeled compounds (the parent compounds themselves and corresponding 3-O-methyl-metabolites) by shifting to COMT metabolism in plasma by the AADC inhibition. In contrast, although the chemical structure of 11C-6MemTyr raised the expectation that it is a substrate of peripheral AADC for decarboxylation to facilitate the brain uptake by carbidopa, it was not significantly affected by a clinical by-mouth dose of carbidopa. Previous studies using 18F-FmT demonstrated that the administration of carbidopa increased the level of 18F-FmT in plasma only by 15% (24) and also that the products of AADC-mediated decarboxylation were detected in plasma even with carbidopa administration (33). To assess the substrate specificity of 11C-6MemTyr to AADC, carbidopa was intravenously administered to achieve higher plasma concentration than clinical by-mouth administration. As a result, the plasma level of 11C-6MemTyr was increased and its metabolites, 11C-6MemTA and 11C-6MHPAA, were decreased in a dose-dependent manner, suggesting that 11C-6MemTyr could also be a substrate of peripheral AADC. The plasma metabolic analyses of 11C-6MemTyr revealed that the inhibitory action of AADC by carbidopa at 5 mg/kg by mouth may be comparable to that determined by its intravenous administration at 0.15 mg/kg, both of which could be too low to affect the brain uptake of 11C-6MemTyr in the living brain. Taken together, 11C-6MemTyr should be useful as a PET probe for presynaptic dopamine imaging, because it can provide the sufficiently high Ki values in the striatum of the living brain even without carbidopa, and its MTGA Ki value was as high as that of 18F-FmT in the monkey brain (22).
Because 3-O-methyl metabolites of l-DOPA in plasma are known to cross the blood–brain barrier into the brain, the COMT products of l-DOPA–based PET probes, β-11C-l-DOPA and 18F-FDOPA, resulted in the higher background activity in nondopamine regions than that of 11C-6MemTyr. Among these 3 probes, 18F-FDOPA revealed the lowest detection capability for MPTP-induced dopaminergic neuronal impairment in the living monkey brain. The metabolisms were substantially different between β-11C-L-DOPA and 18F-FDOPA in plasma; thus, 18F-FDOPA is less stable with more production of its 3-O-methyl metabolite than 3H/11C-l-DOPA in the plasma of rats (7,34) and monkeys (6,35). Metabolic analyses of rat brain homogenates indicated that the radioactivity of l-11C-O-methyl-DOPA contributed 17% of total radioactivity at 40 min after the injection of β-11C-l-DOPA (36), whereas 18F-O-methyl-FDOPA from 18F-FDOPA contributed about 40%–50% (37), resulting in the low MTGA Ki value of 18F-FDOPA. In contrast, because, along with no O-methyl-metabolite by COMT, the peripherally produced metabolites of 11C-6MemTyr, 11C-6MemTA, and 11C-6MHPAA may not cross the blood–brain barrier assumed from the metabolism of structure-relevant 18F-FmT in plasma (24), the time–activity curve data exhibited the rapid and efficient clearance of its unconverted 11C-6MemTyr from nondopamine regions in the brain resulting in the high MTGA Ki value of 11C-6MemTyr. 18F-FmT is decarboxylated to 18F-FmTA by AADC in brain tissue, which has low affinity for the vesicular transporter, and is oxidized and trapped in axon terminals as 18F-FHPAA by monoamine oxidase-A without being released or processed further (20). The metabolic analysis of 11C-6MemTyr in the brain has not been conducted yet, but its metabolic analysis in plasma may be assumed to follow the same metabolic fate as 18F-FmT.
Because AADC is present in the terminals of all monoaminergic neurons, the uptake of 18F-DOPA, 11C-6MemTyr, and β-11C-l-DOPA into extrastriatal regions provides an index of the density of not only the dopamine, but also the serotonergic and noradrenergic terminals (38). However, when applied to assess density in the striatal regions, these uptake indices could exclusively be defined as the AADC activity of dopaminergic neurons. This interpretation is strongly supported by the present results that the striatal dopamine synthetic indices measured using these 3 probes were markedly decreased after the treatment by MPTP, a dopaminergic neuron-specific toxin; and MPTP-induced reduction degrees of striatal dopamine syntheses were significantly correlated with those of DAT availability. The present study applied 11C-β-CFT, a DAT PET probe, as the gold standard for presynaptic dopamine function, for correlation analyses with each PET probe for dopamine synthesis. MTGA Ki values of 11C-6MemTyr revealed a greater correlation coefficient to the degrees of dopaminergic neuronal activity than any of the other 2 PET probes. One reason for this result could be a common metabolic property among β-11C-l-DOPA and 18F-FDOPA, with catechol structure for a substrate of COMT. In contrast, because the 11C-6MemTyr does not have a catechol structure for the 3-O-methyl reaction, the present result should be reasonable from a kinetic point of view. The improved uptake constant of 11C-6MemTyr is also expected to result in its improved sensitivity to changes in presynaptic dopaminergic function compared with β-11C-l-DOPA and 18F-FDOPA.
CONCLUSION
The present study demonstrated that 11C-6MemTyr could be a potential PET probe for quantitative imaging of the presynaptic dopamine activity in the living brain with PET. Noninvasive assessment of AADC activity using 11C-6MemTyr could be useful for diagnosis, prognosis, and treatment monitoring of diseases related to impaired presynaptic dopamine function.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. No potential conflict of interest relevant to this article was reported.
Acknowledgments
We gratefully acknowledge the technical assistance of Shingo Nishiyama, Dai Fukumoto, Aiko Iwazaki, and Naomi Takino.
Footnotes
Published online Nov. 12, 2015.
- © 2016 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication June 2, 2015.
- Accepted for publication October 22, 2015.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}