


Abstract
Preoperative noninvasive imaging to estimate the quantity and spatial distribution of protoporphyrin IX (PpIX) accumulation in tumors induced by 5-aminolevulinic acid (ALA) administration is expected to improve the efficacy of ALA-based fluorescence-guided resection and photo- and sonodynamic therapies. PpIX synthesis from exogenous ALA has been reported to be regulated by ALA influx or ALA dehydratase (ALAD) activity, which catalyzes the first step of the synthesis. In this study, we characterized the properties of a 11C-labeled ALA analog, 5-amino-4-oxo-[6-11C]hexanoic acid (11C-MALA), as a PET tracer to estimate PpIX accumulation. Methods: In vitro uptake of 11C-MALA and 3H-ALA was determined in 5 tumor cell lines after 10-min incubation with each tracer at 37°C. The expression levels of ALAD were determined by Western blot analysis. In vivo distribution and dynamic PET studies were conducted in tumor-bearing mice. In vitro and in vivo accumulation of ALA-induced PpIX was determined by measuring fluorescence in extracts of cells or tumors. Results: In vitro uptake of 11C-MALA in 5 tumor cell lines was correlated with ALAD expression levels and PpIX accumulation. In vivo biodistribution and dynamic PET studies showed that 11C-MALA was rapidly incorporated into tumors, and the tumor-to-muscle ratio of 11C-MALA at 1 min after injection was significantly correlated with that of 3H-ALA. 11C-MALA in tumors was continuously decreased thereafter, and the elimination rate of 11C-MALA from AsPC-1 tumors with the highest ALAD expression level was slower than from other tumors with lower expression levels. These results suggest that the influx and intracellular retention of 11C-MALA reflect ALA influx and ALAD expression levels, respectively. Tumor accumulation of 11C-MALA at 60 min after injection was strongly correlated with PpIX accumulation in tumor tissues. Conclusion: 11C-MALA PET has the potential to noninvasively estimate the quantitative and spatial accumulation of exogenous ALA-induced PpIX.
Fluorescence-guided resection (FGR) for resectable glioma (1,2) and photodynamic therapy (PDT) (2,3) and sonodynamic therapy (SDT) (4) for small-sized, diffused, or residual tumors using tumor-specific accumulation of protoporphyrin IX (PpIX) induced by 5-aminolevulinic acid (ALA) administration are promising therapeutic strategies. ALA, a precursor of PpIX, is a nonfluorescent and photo- and sonodynamically inert prodrug. ALA induces selective accumulation of PpIX, a fluorescent photo- and sonosensitizer, in tumor tissues by tumor-specific metabolism (3). ALA administration enables intraoperative identification of tumors in FGR and tumor-selective cytotoxicity by photo- or sonoirradiation in PDT/SDT with low toxicity in normal tissues (5). Progression-free survival in patients with malignant glioma has been improved by FGR as it facilitated more complete resection of tumors, compared with conventional surgery (6). ALA PDT also showed objective responses in several cancerous diseases such as nonmelanoma skin (1) and bladder cancers (7). On the other hand, there are apparent differences in the accumulation of ALA-induced PpIX among tumors (8,9). These differences represent key factors influencing the therapeutic efficacy of ALA-based PDT/SDT (10,11). The precise assessment of PpIX accumulation in each tumor type can therefore play a significant role in predicting the therapeutic effects and selecting the appropriate patients who could potentially benefit from ALA-based PDT/SDT. Furthermore, PpIX accumulates heterogeneously within tumor tissues, leading to incomplete resection in FGR for glioma (9,12). The spatial distribution of ALA-induced PpIX in whole tumor regions determined before FGR should contribute to complete resection. The preoperative assessment of ALA-induced PpIX-accumulating regions in each tumor would provide landmarks during operations for glioma.
Fluorescence imaging of PpIX can directly evaluate the quantity of PpIX accumulation (13); however, the poor penetration of the light restricts the application for deeply seated tumors in patients before therapy. CT, MR imaging, and PET have no tissue penetration limit. Several studies were undertaken to detect regions of ALA-induced PpIX accumulation in malignant tumors with noninvasive tomographic imaging such as gadolinium-enhanced MR imaging (14) or PET with l-[methyl-11C]-methionine (11C-MET) (15) or O-(2-18F-fluoroethyl)-l-tyrosine (18F-FET) (12). Those studies showed that there were some differences between gadolinium-enhanced MR imaging or 11C-MET or 18F-FET PET imaging and PpIX accumulation (12,15), because these diagnostic images provided information different from PpIX accumulation—that is, gadolinium-enhanced MR imaging and PET with 11C-MET or 18F-FET reflect the blood–brain barrier breakdown (16) or amino acid metabolism (17) but not PpIX accumulation. Therefore, a new noninvasive imaging method based on the mechanism of PpIX accumulation is needed. PpIX accumulation has been considered to be modulated by the influx of ALA via transporters (10,18) and the activity of enzymes that catalyze the biosynthesis of heme from ALA (19–21). PET enables functional and quantitative assessments of several biologic processes such as transport and metabolism (22,23). PET with an appropriate tracer is therefore an attractive candidate modality to estimate the quantitative and spatial accumulation of PpIX in tumors.
The influx of ALA into cells (10,18) and expression levels of ALA dehydratase (ALAD) that catalyzes the first step of PpIX synthesis from ALA play a major and rate-determining role in regulating PpIX accumulation (19,20). Accordingly, a PET tracer to target ALA influx and ALAD expression would be useful for predicting PpIX accumulation in tumors. The ALA analog 5-amino-4-oxohexanoic acid (MALA) is an inhibitor of ALAD by covalent binding with the catalytic center of ALAD to form a carbinolamine intermediate (24). Recently, we have successfully synthesized 11C-labeled MALA (5-amino-4-oxo-[6-11C]hexanoic acid, 11C-MALA) as a PET tracer for the above-mentioned purpose (25). We have previously shown that 11C-MALA is incorporated into tumor cells by transporters similar to ALA (25). In the present study, we examined the relationship between 11C-MALA and ALA-induced PpIX accumulation in tumors to evaluate the properties of 11C-MALA as a PET tracer to reflect tumor accumulation of PpIX induced by ALA administration.
MATERIALS AND METHODS
Chemicals
11C-MALA was prepared as described previously (25). Aminolevulinic acid [3,5-3H(N)] hydrochloride (3H-ALA, 740 GBq/mmol) was obtained from American Radiolabeled Chemicals. Other chemicals were of analytic grade without further purification.
Cells and Animals
Human tumor cell lines (AsPC-1, Sk-Br-3, U-87 MG, BxPC-3, and MIA PaCa-2) and tumor-bearing mice were prepared as described in the Supplemental Methods (supplemental materials are available at http://jnm.snmjournals.org). All animal experiments were performed in accordance with the institutional guidelines regarding animal care and handling after approval by the Animal Care and Use Committee of the National Institute of Radiological Sciences.
Cellular Uptake Studies
Cells were incubated with 11C-MALA (∼15 MBq) or 3H-ALA (18.5 kBq) for 10 min, and then radioactivity in cells was measured as described in the Supplemental Methods. Cellular uptake was expressed as the percentage of administered radioactivity per milligram of protein (n = 3).
Western Blotting
Western blotting was conducted using anti-ALAD or anti–β-actin antibodies as described in the Supplemental Methods. Relative ALAD expression levels were expressed as the ratio of intensity of ALAD to those of β-actin (n = 3).
In Vitro Accumulation of ALA-Induced PpIX
Cells were incubated with or without 1 mM ALA (Wako Pure Chemical Industries). After 4-h incubation, fluorescence of PpIX in cells was measured as described in the Supplemental Methods. Results were normalized to protein content (n = 3).
Biodistribution Studies
Normal (n = 5 each group) or tumor-bearing mice (n = 4 each group) were intravenously injected with the mixture of 11C-MALA and 3H-ALA. Mice were sacrificed, and 11C and 3H radioactivity of tissues was measured as described in the Supplemental Methods. Results are given as the percentage injected dose per gram of tissue (%ID/g).
In Vivo Stability of 11C-MALA
AsPC-1 tumor–bearing mice were intravenously injected with 11C-MALA (50.06 MBq) and sacrificed at 10 min after injection. Plasma and homogenates of organs from mice were deproteinized and then analyzed by radio–high-performance liquid chromatography as described in the Supplemental Methods.
Dynamic PET Imaging
Tumor-bearing mice were intravenously injected with 11C-MALA (11.35 ± 0.94 MBq), and dynamic PET scans were obtained for 90 min as described in the Supplemental Methods. The region of interest was manually drawn over tumors, and radioactivity was quantified as the mean concentration in the regions of interest (n = 4). The clearance of radioactivity from tumors was analyzed by biexponential least-square curve-fitting. Retention fractions (RFs) were determined by dividing the intercept in the extrapolated slope of the late phase of time–activity curve back to the time of maximal activity by maximal activity (Supplemental Fig. 1).
In Vivo Blocking Study
Unlabeled ALA (1 or 4 mg) was coinjected with 11C-MALA (11.59 ± 0.82 MBq) into AsPC-1 tumor–bearing mice. The radioactivity in blood was measured at various time points (n = 4 at each time point). Dynamic PET scans were obtained, and data were analyzed (n = 4).
In Vivo Tumor Accumulation of PpIX
Tumor-bearing mice were intravenously injected with ALA (6 mg). At 4 h after injection, PpIX accumulation in tumor tissues was determined as described in the Supplemental Methods (n = 4).
Ex Vivo Autoradiography, Immunofluorescence, and Florescence Microscopy
AsPC-1 tumor–bearing mice were intravenously injected with ALA (6 mg). At 3 h after injection of ALA, the mice were intravenously injected with 11C-MALA (84.91 MBq). After a further 60 min, removed tumors were frozen, and distribution of radioactivity, ALAD, and PpIX were observed in the same sections as described in the Supplemental Methods. The sections were also stained with hematoxylin and eosin for histologic evaluation.
Statistical Analysis
Data are presented as mean ± SD. The association between variables was calculated by Pearson product moment correlation coefficient. Data of the blocking study were compared using ANOVA with the Dunnett multiple comparison test.
RESULTS
Cellular Uptake of 11C-MALA and 3H-ALA
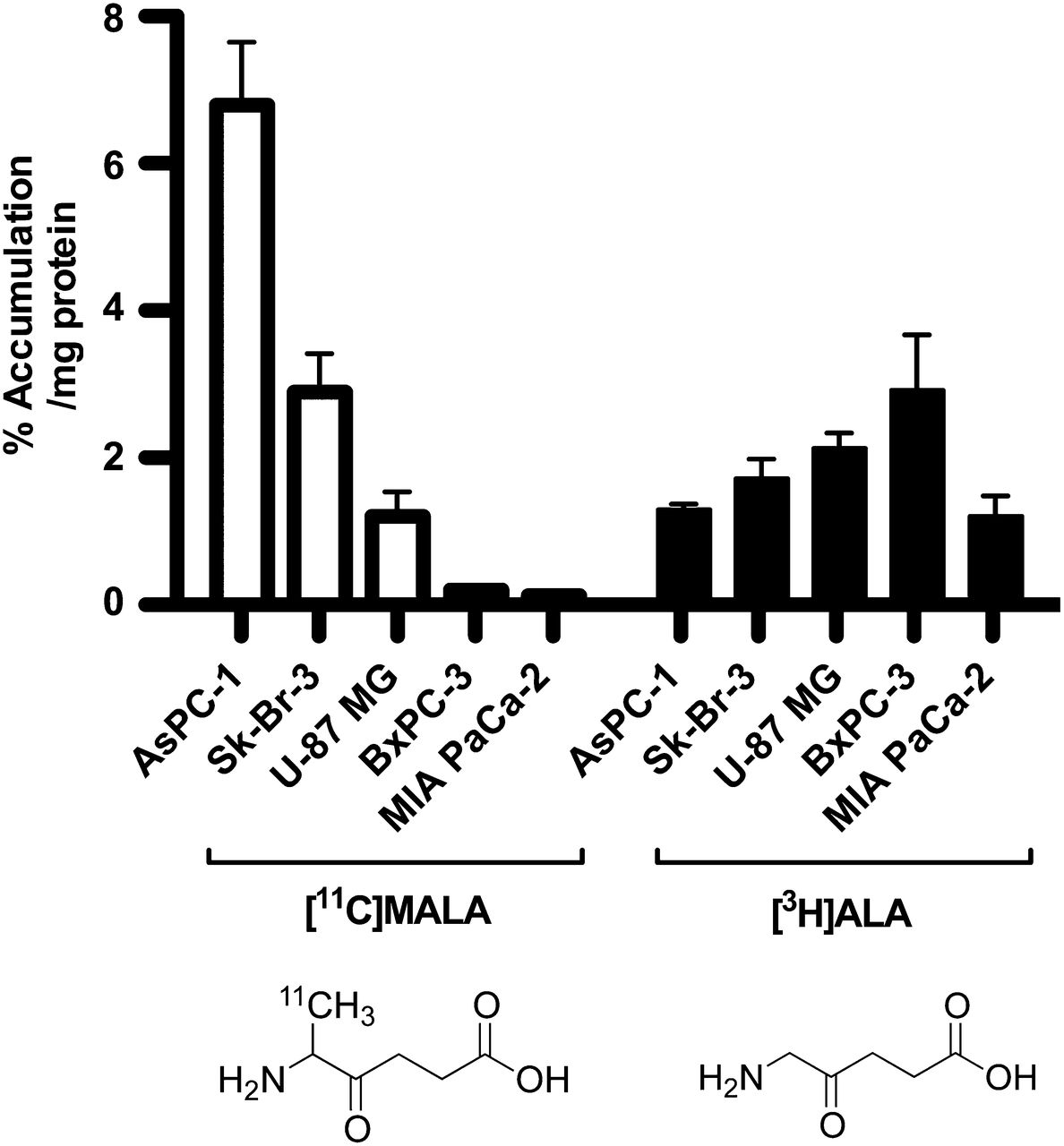
The highest uptake of 11C-MALA was observed in the AsPC-1 cell line, followed by Sk-Br-3 and U-87 MG. Uptake of 11C-MALA in BxPC-3 and MIA PaCa-2 cells was less than 5% of that in AsPC-1 (Fig. 1). In contrast, there was little difference in the cellular uptake of 3H-ALA between the 5 cell lines, compared with 11C-MALA (Fig. 1). There was no correlation between the cellular uptake of 11C-MALA and 3H-ALA (R2 = 0.23, P = 0.48).
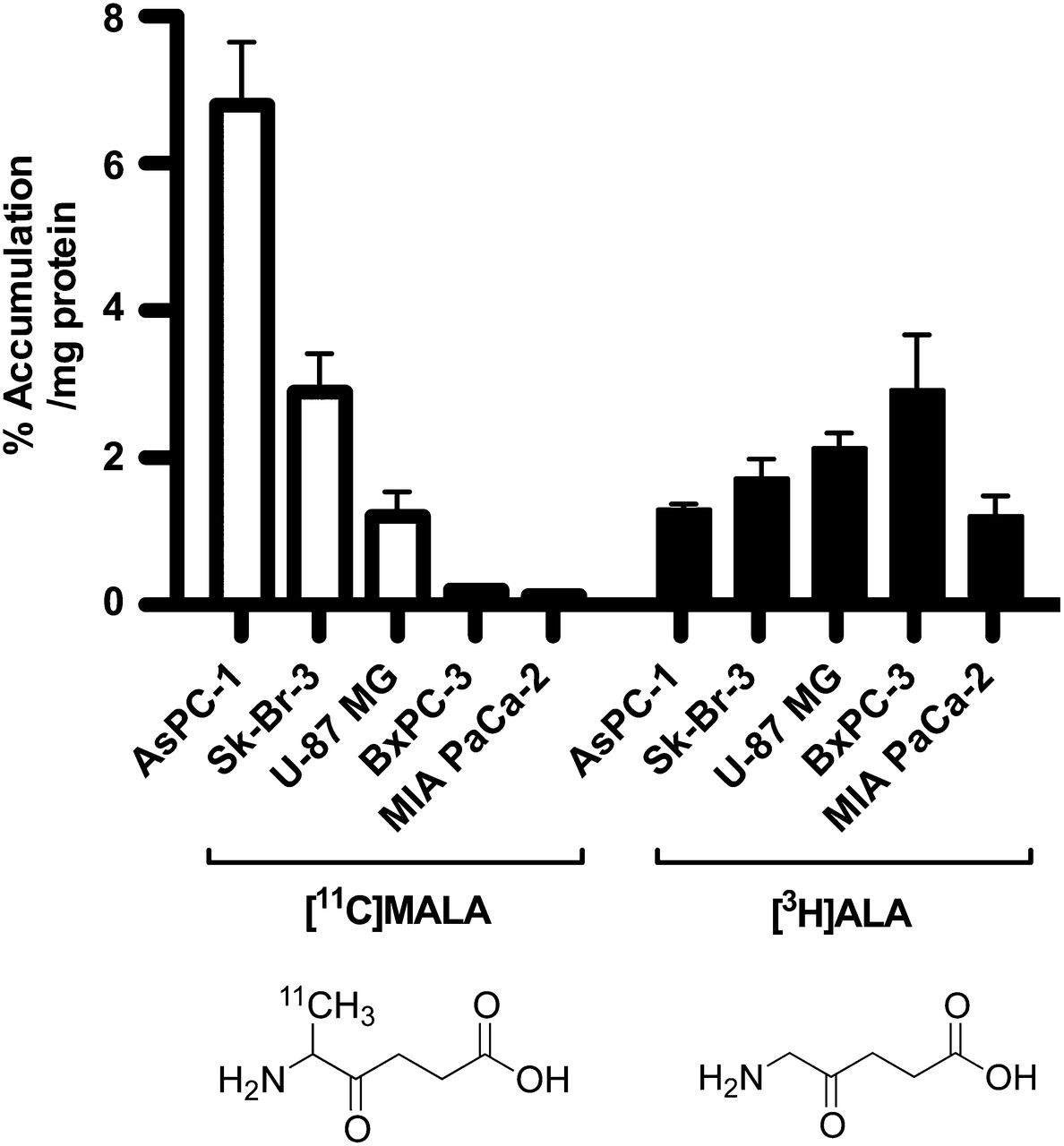
In vitro accumulation of 11C-MALA (open bars) and 3H-ALA (closed bars) in 5 cell lines.
ALAD Expression in Tumor Cell Lines
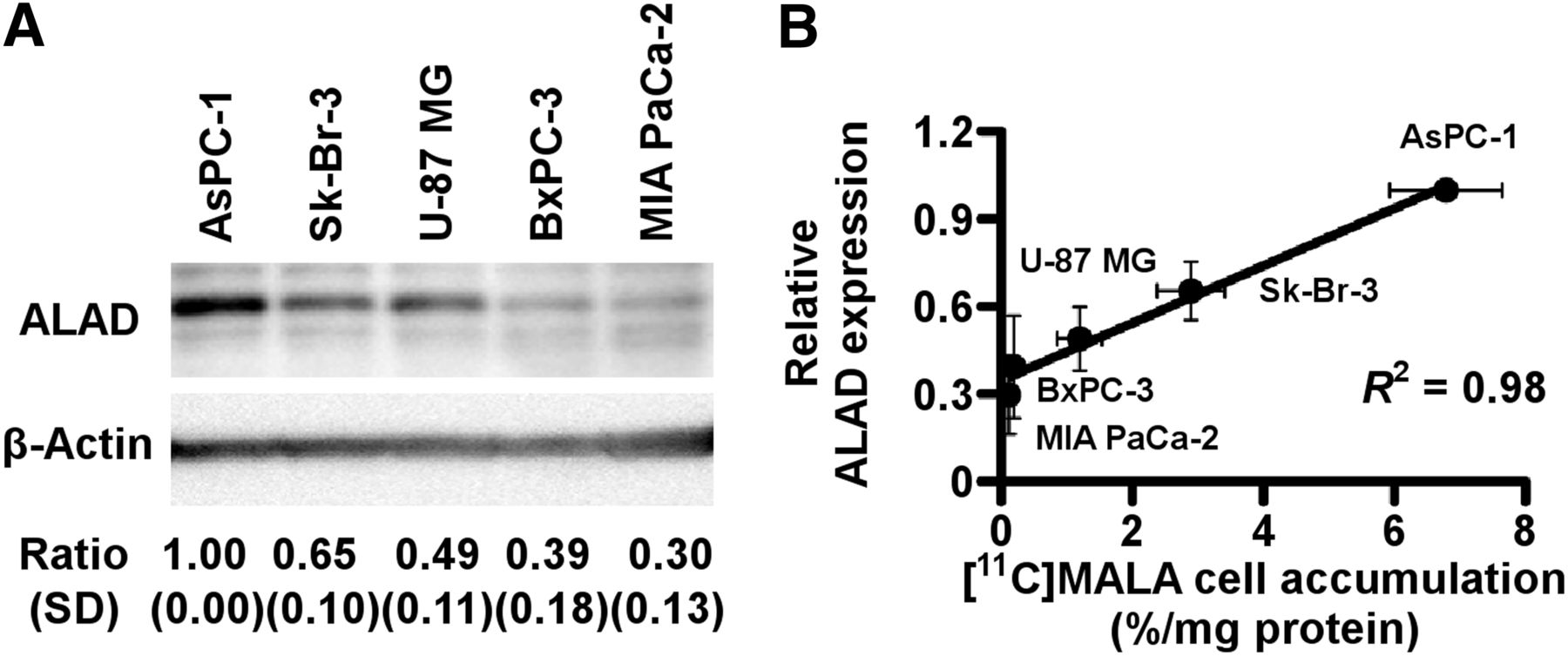
AsPC-1 showed the highest expression, followed by Sk-Br-3 and U-87 MG. BxPC-3 and MIA PaCa-2 showed low expression levels (Fig. 2A). The expression levels of ALAD were strongly correlated with the cellular uptake of 11C-MALA, as determined by the above-mentioned cell uptake studies (Fig. 2B, R2 = 0.98, P < 0.01).
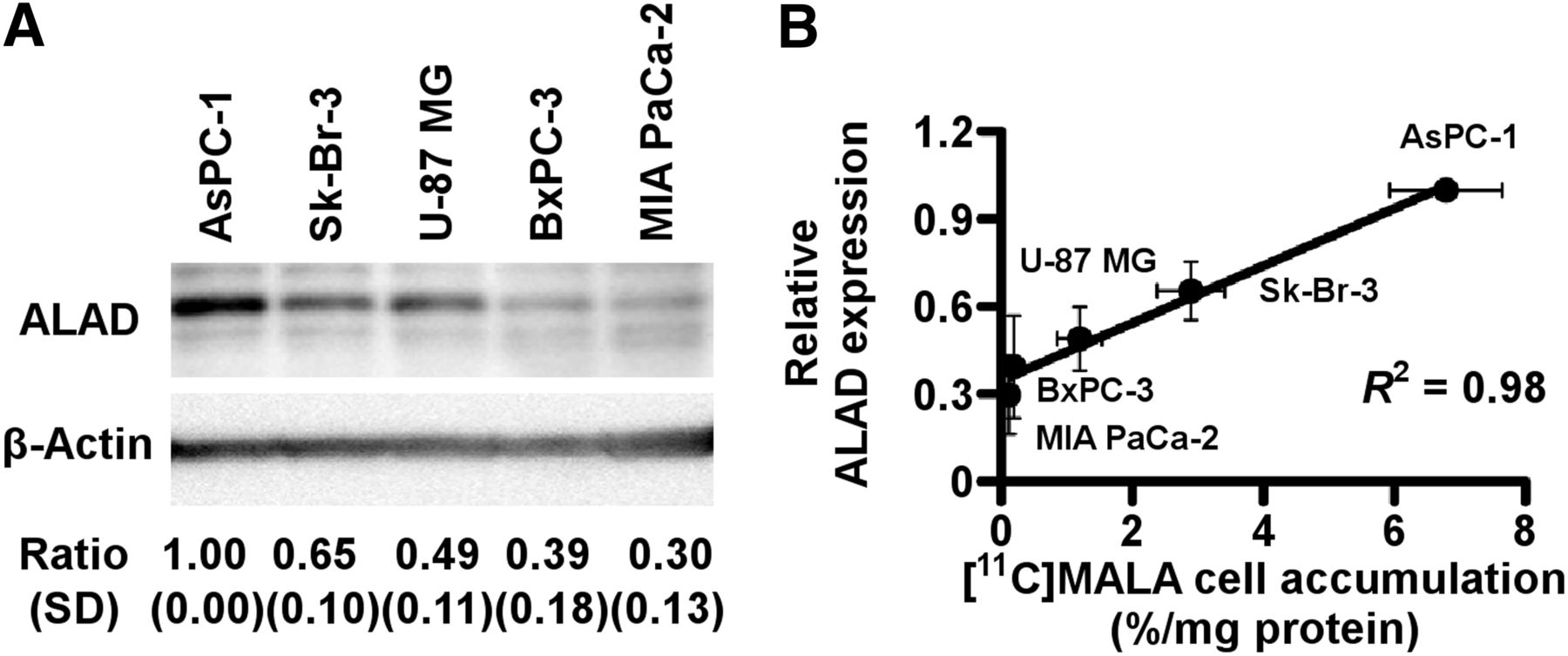
ALAD expression levels in 5 tumor cell lines. (A) Relative ALAD protein expression of 5 tumor cell lines determined by Western blotting. ALAD expression levels were normalized to β-actin. (B) Correlation between cellular 11C-MALA accumulation and relative ALAD expression levels.
In Vitro ALA-Induced PpIX Accumulation
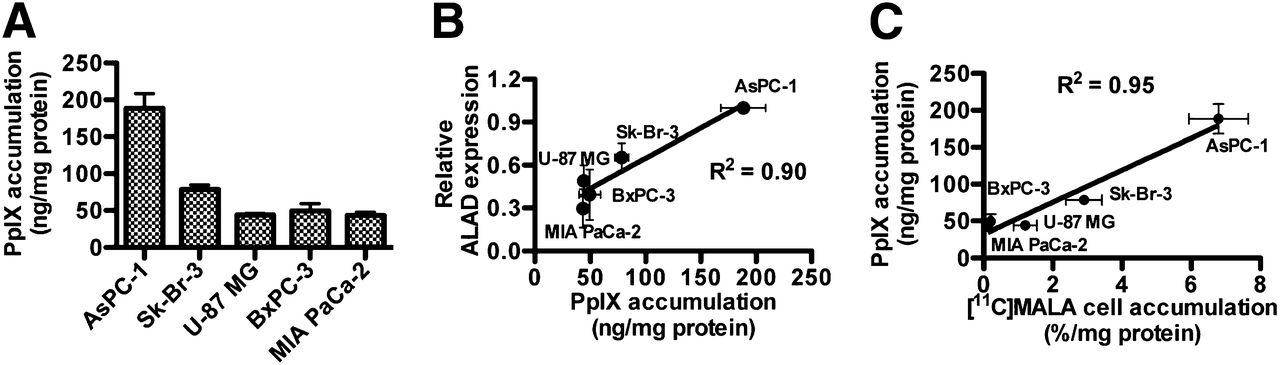
Although fluorescence was undetectable (<30 pg PpIX/well) in all 5 cell lines after incubation without ALA (data not shown), marked increases in cellular PpIX were induced by incubation with ALA (Fig. 3A). AsPC-1 showed the highest ALA-induced PpIX accumulation, and Sk-Br-3 showed approximately 40% PpIX accumulation of AsPC-1. PpIX accumulation in the other 3 cells was less than 30% of that in AsPC-1. A strong linear relationship was observed between ALA-induced PpIX accumulation and ALAD expression levels (Fig. 3B, R2 = 0.90, P < 0.05). ALA-induced PpIX accumulation was also strongly correlated with cellular uptake of 11C-MALA (Fig. 3C, R2 = 0.95, P < 0.01). Cellular uptake of 3H-ALA was not correlated with the accumulation of PpIX (Supplemental Fig. 2, R2 = 0.23, P = 0.48).

In vitro PpIX accumulation induced by ALA. (A) ALA-induced PpIX accumulation in cells after incubation with ALA. (B) Correlation between relative ALAD expression levels and ALA-induced PpIX accumulation. (C) Correlation between cellular accumulations of 11C-MALA and ALA-induced PpIX.
Biodistribution of 11C-MALA and 3H-ALA in Normal Mice
11C radioactivity in blood rapidly decreased from 10.54 ± 0.93 %ID/g at 1 min to 3.94 ± 1.80 %ID/g at 10 min after injection (Supplemental Table 1). The highest accumulation of 11C-MALA was observed in the kidneys at 10 min (91.30 ± 6.53 %ID/g), and renal radioactivity rapidly decreased thereafter, indicating that the kidneys are the major excretion route of 11C-MALA (Supplemental Table 1). 3H-ALA in blood also decreased rapidly (Supplemental Table 2), just as 11C-MALA.
In Vivo Tumor Accumulation of 11C-MALA and 3H-ALA
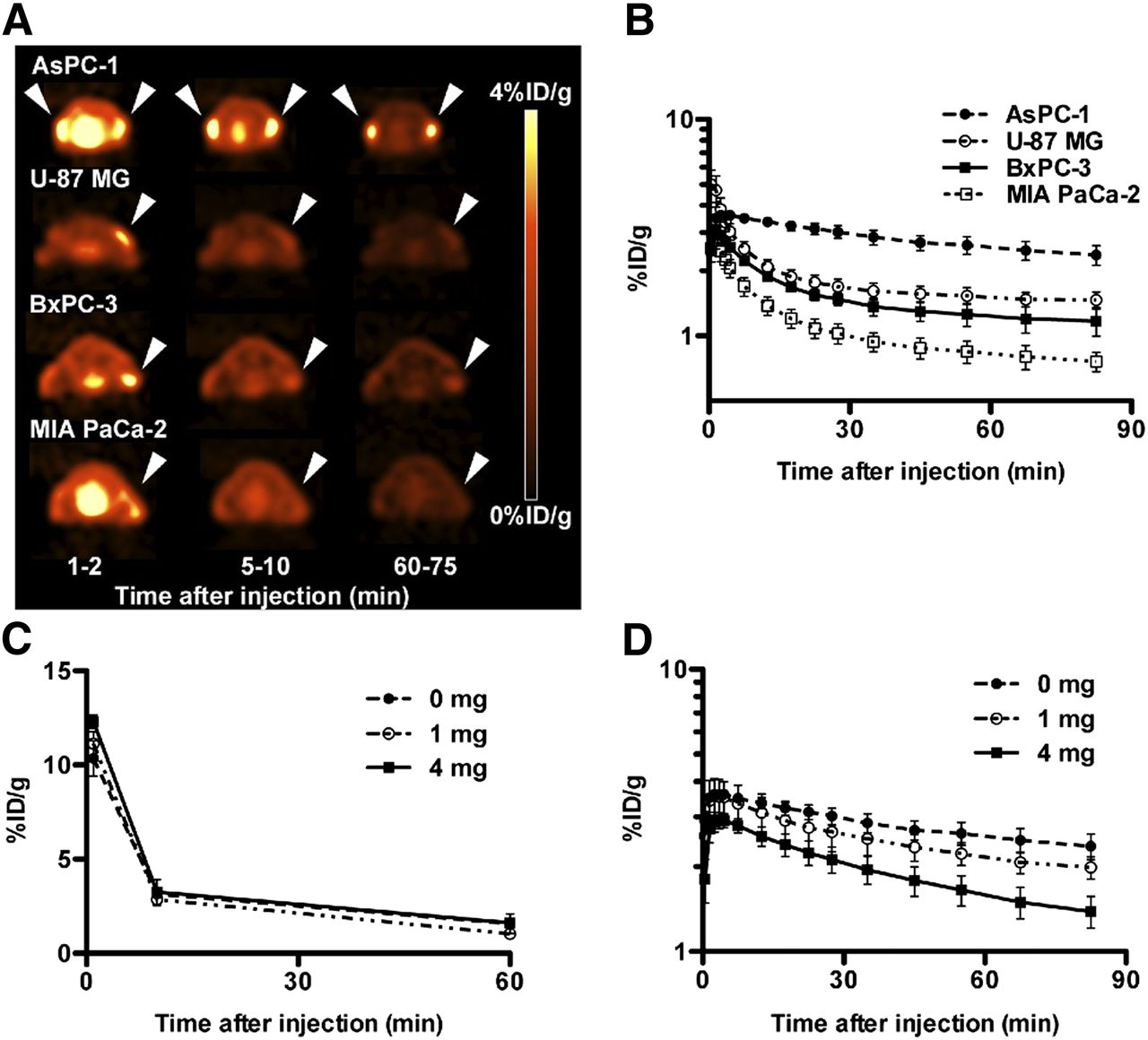
Because Sk-Br-3 cells formed no xenograft tumor even in female nude mice under our conditions, the tumor accumulations of 11C-MALA, 3H-ALA, and ALA-induced PpIX were evaluated using 4 cell lines, AsPC-1, U-87 MG, BxPC-3, and MIA PaCa-2. AsPC-1 tumors highly retained 11C-MALA, but the other 3 tumors did not (Table 1). There was little difference in 3H radioactivity in tumors at 10 and 60 min after 3H-ALA injection between the 4 tumors (Table 1). The tumor-to-muscle ratios of 11C-MALA at 1 min after injection were significantly correlated with those of 3H-ALA (Supplemental Fig. 3, R2 = 0.96, P < 0.05).
Tumor Accumulation and Tumor-to-Muscle Ratios of 11C-MALA and 3H-ALA in Tumor-Bearing Mice
In Vivo Stability of 11C-MALA
Chemical structures of radioactivity at 10 min after injection of 11C-MALA were analyzed by radio–high-performance liquid chromatography. No metabolite was observed in the deproteinized plasma and supernatants of liver, spleen, and AsPC-1 tumor (Supplemental Fig. 4).
Dynamic PET Imaging
The uptake of 11C-MALA in the 4 different tumors rapidly reached the peak value within 5 min after injection, and then decreased continuously (Figs. 4A and 4B). 11C-MALA showed much slower elimination from AsPC-1 than the other 3 tumors (Figs. 4A and 4B). The RF of AsPC-1 was highest, and RFs of U-87 MG, BxPC-3, and MIA PaCa-2 were lower (Table 2).
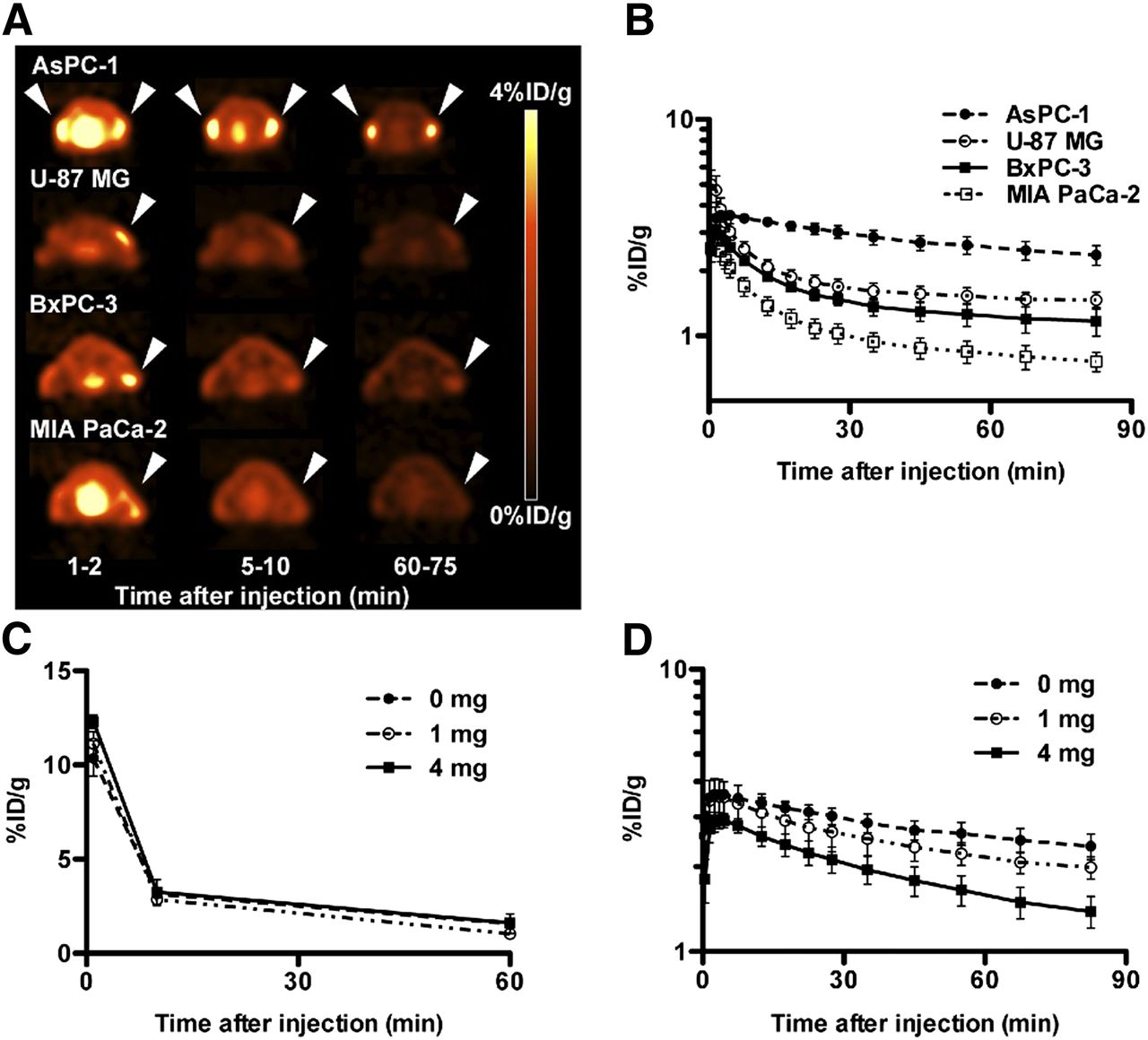
Dynamic PET and in vivo blocking studies of 11C-MALA. (A) Transaxial PET images of mice bearing tumor xenografts (arrowheads) injected with 11C-MALA (∼11.1 MBq). (B) Time–activity curves of 11C-MALA in tumors determined by PET studies. (C) Time–activity curves of 11C-MALA in blood of mice coinjected with unlabeled ALA (0, 1, or 4 mg) determined by biodistribution study. (D) Time–activity curves of 11C-MALA in AsPC-1 tumors coinjected with unlabeled ALA (0, 1, or 4 mg) determined by PET study.
RFs of Tumors Determined by Time–Activity Curve Analysis of 11C-MALA PET
In Vivo Blocking Study
The radioactivity of 11C-MALA in blood was hardly affected by coinjected ALA (Fig. 4C), whereas the initial tumor uptake was decreased and the elimination rate from tumors was accelerated by increasing the amount of ALA (Fig. 4D). By adding excess ALA as a carrier, the RFs of 11C-MALA were dose-dependently decreased (Table 2).
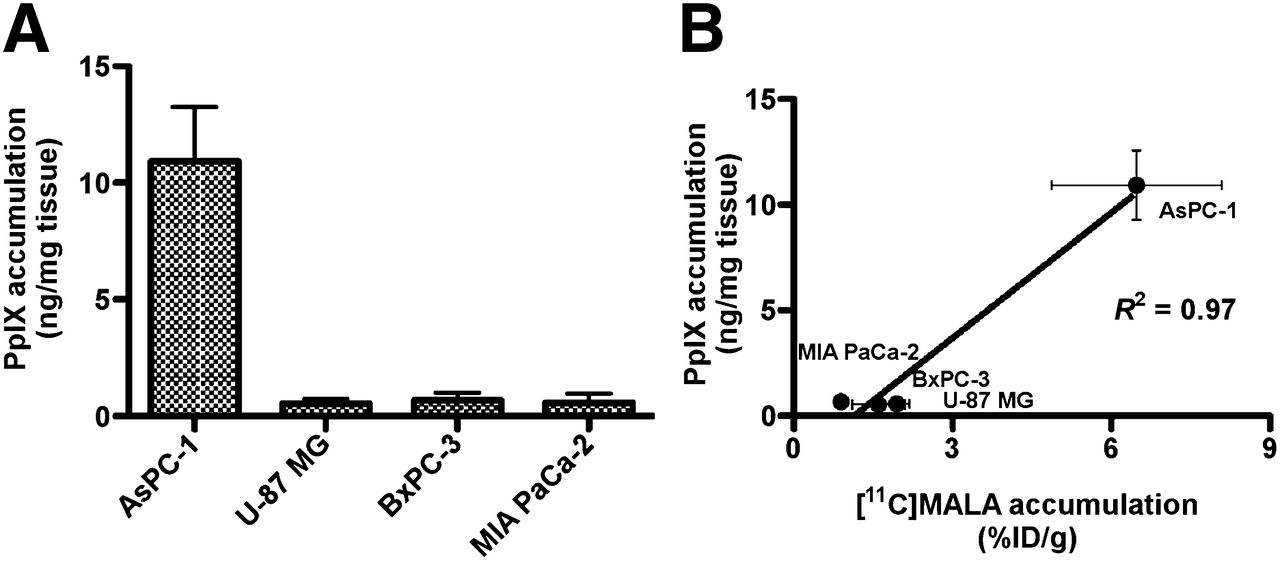
In Vivo Tumor Accumulation of PpIX
The concentration of porphyrins in several tumors has been reported to reach a maximum from 3 to 5 h after intravenous administration of ALA (300 mg/kg of body weight) (26). We therefore measured PpIX fluorescence in tumors at 4 h after administration of 6 mg of ALA. The highest PpIX accumulation was observed in AsPC-1 tumor. PpIX accumulation in the other 3 tumors was less than one tenth of that in AsPC-1 (Fig. 5A). A strong positive correlation was found between tumor accumulation of PpIX and 11C-MALA at 60 min after injection (Fig. 5B, R2 = 0.97, P < 0.05).
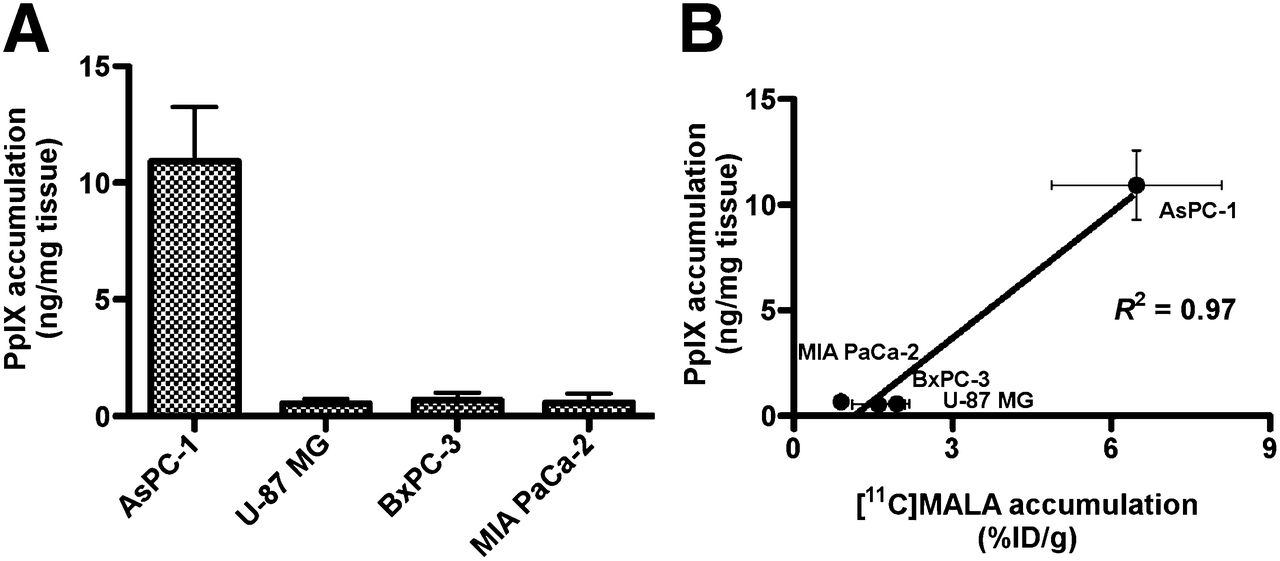
ALA-induced PpIX accumulation in xenograft tumors. (A) In vivo ALA-induced PpIX accumulation in tumors after ALA injection. (B) Correlation between in vivo tumor accumulation of ALA-induced PpIX at 4 h after injection of ALA and 11C-MALA at 60 min after injection.
Intratumoral Distribution of 11C-MALA, ALAD Expression, and ALA-Induced PpIX
Autoradiograms showed heterogeneous and focal accumulation of 11C radioactivity within tumor sections (Fig. 6A). ALA-induced PpIX accumulated in almost the same area as that in which the strong signal of ALAD was observed (Figs. 6B and 6C). Intratumoral distribution patterns of 11C-MALA were not completely identical but tended to be similar to the distribution patterns of ALAD expression levels and PpIX accumulation. These heterogeneous distributions were not consistent with necrotic regions evaluated by hematoxylin and eosin staining (Fig. 6D).
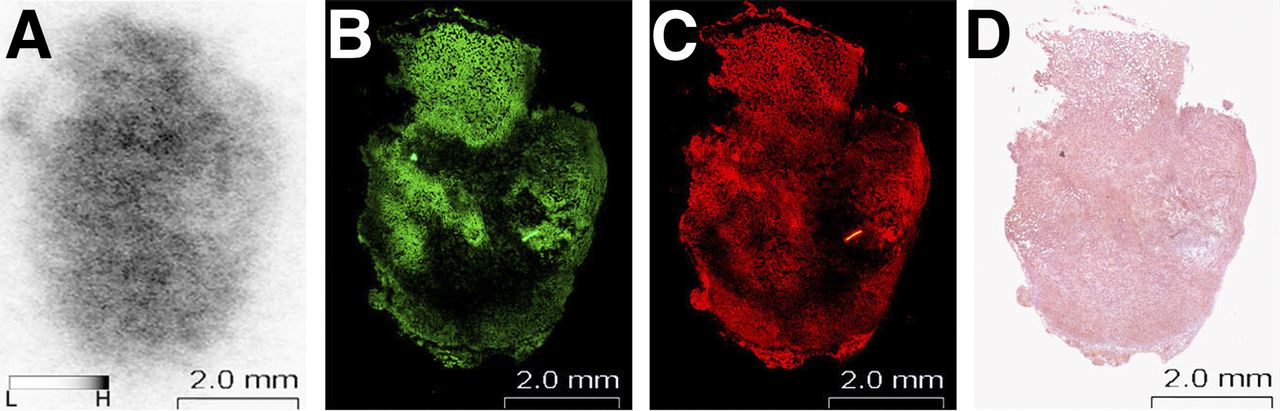
Intratumoral distribution of 11C-MALA, ALAD expression, and ALA-induced PpIX in same tumor section. (A) Autoradiogram of 11C radioactivity at 60 min after injection of 11C-MALA. (B) Immunofluorescent image for ALAD. (C) Fluorescent image of PpIX. (D) Hematoxylin and eosin stain.
DISCUSSION
ALA-induced PpIX accumulation has been reported to be affected by ALA influx into tumor cells (10,18). The present in vitro experiments, however, showed no correlation between 3H-ALA radioactivity and PpIX accumulation in the tumor cells. Similar results have been reported in different tumors (27). The reason for this would be primarily due to the difference in PpIX formation rate mediated by some enzyme activities among tumor cells. These findings imply that a PET tracer for estimating ALA-induced PpIX accumulation should possess the ability to evaluate not only ALA influx but also ALA metabolism. The cell uptake studies showed no correlation between 3H-ALA and 11C-MALA in the tumor cells, even though 11C-MALA was incorporated into tumor cells through transport systems similar to ALA (25). These results suggest that there are some differences in intracellular behavior, such as metabolism or efflux or both, between 3H-ALA and 11C-MALA after the incorporation of each tracer. 3H would exist intracellularly as 3H-ALA and various heme-biosynthesis metabolic intermediates, and 11C-MALA unbound to the catalytic center of ALAD would be presumably excreted from cells. In the present study, 11C-MALA uptake in tumor cells was strongly correlated with ALAD expression levels and ALA-induced PpIX accumulation. Because ALAD plays a rate-limiting role in regulating PpIX accumulation (19,20), 11C-MALA would reflect PpIX accumulation in tumor cells as a sum of the ALA influx and metabolism by ALAD.
In vivo biodistribution and dynamic PET studies in tumor mouse models showed that 11C-MALA was rapidly incorporated into tumors. Considering that 11C-MALA is incorporated into tumor cells via transport systems similar to ALA (25), the initial uptake of 11C-MALA would mainly reflect the influx or efflux rate of ALA rather than the metabolism of ALA to PpIX. This is supported by the similar tumor accumulation and tumor-to-muscle ratios of 11C-MALA and 3H-ALA at an early time point after injection.
11C-MALA was rapidly cleared from blood, and no radiometabolite was observed in plasma and supernatants of liver, spleen, and AsPC-1 tumor. These results indicate that 11C-MALA would be stable in vivo, and there is little additional supply of radioactivity from blood except for that just after injection. Thus, the decrease in radioactivity after the initial uptake in tumors would mainly reflect the efflux of 11C-MALA from tumors. On the basis of the PET studies, the elimination rates of 11C-MALA from tumor tissues differed between the 4 xenograft models and followed biphasic patterns, suggesting that there are 2 components, each having a distinct elimination rate. These results raise the following hypothesis: there could be 2 radiochemical species, 11C-MALA and 11C-MALA-ALAD complex, in tumor cells (Fig. 7). The 11C-MALA-ALAD complex would be retained intracellularly, whereas 11C-MALA would be eliminated from cells. The RFs determined by 11C-MALA PET would reflect the ratio of the 11C-MALA-ALAD complex formation to total intracellular 11C-labeled components in this model. AsPC-1 tumors with high ALAD expression levels showed longer retention and higher RF of 11C-MALA than the tumors with low ALAD expression levels. Furthermore, the blocking studies using excess unlabeled ALA as a carrier showed decreases in both the initial uptake and the RFs in tumors in a dose-dependent manner, even though blood clearance hardly changed. These results suggest that not only the influx of 11C-MALA into cells but also the formation of 11C-MALA-ALAD complex are competitively inhibited by the presence of excess ALA, supporting our hypothesis that 11C-MALA would be intracellularly retained through the formation of 11C-MALA-ALAD complex after incorporation into tumor cells.

Putative model for retention of 11C-MALA in tumor cells.
Tumor radioactivity levels at the latter phase of 11C-MALA elimination would reflect both the initial uptake and the RFs, which would indicate ALA influx and ALAD expression levels, respectively; namely, tumor accumulation of 11C-MALA from approximately 30 min onward would reflect ALA-induced PpIX accumulation. The biodistribution studies revealed that the tumor accumulation of 11C-MALA at 60 min after injection was strongly correlated with that of ALA-induced PpIX, suggesting that 11C-MALA PET could estimate ALA-induced PpIX accumulation defined by influx and metabolism of ALA. Although our findings demonstrated that 11C-MALA PET has the potential to estimate PpIX accumulation in tumors, Hagiya et al. reported that PpIX excretion from tumor cells could play a key role in the PpIX accumulation in their clinical study (18). The mechanism of tumor-specific PpIX accumulation is so complicated that further clinical studies are needed to validate our findings.
It is known that differences in PpIX accumulation among tumors can affect the therapeutic efficacy of ALA-based PDT and SDT (10,11). The noninvasive estimation of ALA-induced PpIX accumulation in each tumor by 11C-MALA PET could enable the selection of appropriate patients who potentially gain benefit from these therapies. In ALA-based FGR, the heterogeneous distribution of PpIX in tumor tissues could be a cause of incomplete resection (9,12). From the autoradiographic studies, although intratumoral distribution of 11C-MALA tended to be similar to that of ALAD expression and PpIX accumulation, there were some differences between them probably because of the relatively long positron range of 11C on autoradiography and different dynamic ranges of autoradiography and immunostaining. Because PET can give not only quantitative information but also spatial information, preoperative 11C-MALA PET combined with morphologic imaging such as CT or MR imaging could estimate the spatial distribution of PpIX accumulation within tumors, providing useful information to facilitate complete resection of tumors in ALA-based FGR.
11C-MALA was evaluated as a racemic mixture in this study. Each enantiomer could show different influx, efflux, or binding to ALAD, because chiral biomolecules generally show enantiospecific biologic properties such as substrate recognition of transporters or enzymes (28). The promising results obtained from the present study with racemic 11C-MALA support further studies to investigate the stereochemical evaluation of 11C-MALA for PpIX accumulation.
CONCLUSION
We demonstrated that the pharmacokinetics of 11C-MALA reflect ALA influx into tumor cells and ALAD expression levels, resulting in a means to estimate the ALA-induced PpIX accumulation. 11C-MALA PET therefore has the potential to noninvasively evaluate the accumulation and spatial distribution of ALA-induced PpIX in tumor tissues and provide helpful information for improving the efficacy of ALA-based FGR and PDT/SDT.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. This work was supported in part by KAKENHI 23510289, 24591803, and 25861136 from MEXT or Japan Society for the Promotion of Science. No other potential conflict of interest relevant to this article was reported.
Acknowledgments
We thank the staff of the Cyclotron Operation and Radiochemistry sections for the production of the radioisotope and the technical support of the remote-controlled synthesis and Hidekatsu Wakizaka for operation and quality control of the PET system.
Footnotes
Published online Aug. 14, 2014.
- © 2014 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication June 29, 2014.
- Accepted for publication July 22, 2014.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.