


Abstract
Subanesthetic doses of ketamine can be used as a rapid-acting antidepressant in patients with treatment-resistant depression. Therefore, the brain kinetics of 123I-5-I-R91150 (4-amino-N-[1-[3-(4-fluorophenyl)propyl]-4-methylpiperidin-4-yl]-5-iodo-2-methoxybenzamide) and the influence of ketamine on the postsynaptic serotonin-2A receptor (5-hydroxytryptamine-2A, or 5-HT2A) status were investigated in cats using micro-SPECT. Methods: This study was conducted on 6 cats using the radioligand 123I-5-I-R91150, a 5-HT2A receptor antagonist, as the imaging probe. Anesthesia was induced and maintained with a continuous-rate infusion of propofol (8.4 ± 1.2 mg kg−1 followed by 0.22 mg kg−1 min−1) 75 min after tracer administration, and acquisition of the first image began 15 min after induction of anesthesia. After this first acquisition, propofol (0.22 mg kg−1 min−1) was combined with ketamine (5 mg kg−1 followed by 0.023 mg kg−1 min−1), and the second acquisition began 15 min later. Semiquantification, with the cerebellum as a reference region, was performed to calculate the 5-HT2A receptor binding indices (parameter for available receptor density) in the frontal and temporal cortices. The binding indices were analyzed with Wilcoxon signed ranks statistics. Results: The addition of ketamine to the propofol continuous-rate infusion resulted in decreased binding indices in the right frontal cortex (1.25 ± 0.22 vs. 1.45 ± 0.16; P = 0.028), left frontal cortex (1.34 ± 0.15 vs. 1.49 ± 0.10; P = 0.028), right temporal cortex (1.30 ± 0.17 vs. 1.45 ± 0.09; P = 0.046), and left temporal cortex (1.41 ± 0.20 vs. 1.52 ± 0.20; P = 0.046). Conclusion: This study showed that cats can be used as an animal model for studying alterations of the 5-HT2A receptor status with 123I-5-I-R91150 micro-SPECT. Furthermore, an interaction between ketamine and the 5-HT2A receptors resulting in decreased binding of 123I-5-I-R91150 in the frontal and temporal cortices was demonstrated. Whether the decreased radioligand binding resulted from a direct competition between ketamine and 123I-5-I-R91150 or from a decreased affinity of the 5-HT2A receptor caused by ketamine remains to be elucidated.
Serotonergic 5-hydroxytryptamine-2A receptors (5-HT2A) can be visualized in vivo using SPECT with the tracer 123I-5-I-R91150 (4-amino-N-[1-[3-(4-fluorophenyl)propyl]-4-methylpiperidin-4-yl]-5-iodo-2-methoxybenzamide). In dogs, activity in the frontal cortex peaks at 90–100 min after intravenous injection, and for approximately 100 min the frontocortical activity (specific and nonspecific tracer binding) remains stable, compared with the cerebellar activity (region void of 5-HT2A receptors; thus, only nonspecific tracer binding), indicating this to be the optimal scanning interval in this species (1). In healthy humans, the optimal scanning interval has been determined by several studies (2–4). To our knowledge, the existence of such a pseudoequilibrium has not yet been described in cats.
Alterations in the binding potential of the serotonergic 5-HT2A receptors, which are associated with major depression or suicidal behavior, have been described in several human studies (3,5–8). An increased or decreased binding potential of the 5-HT2A receptor antagonist 123I-5-I-R91150 was also reported in dogs with, respectively, impulsive aggression or pathologic anxiety (9,10). In humans as well as in dogs, the frontal, temporal, and parietal cortices are the regions mostly involved (5,9,10). Hyperthyroidism has been associated with alterations in serotonergic neurotransmission in humans and rats (11,12). Because this is a frequently occurring pathology in cats (13), this species can serve as an interesting research model. These findings confirm the potential for using dogs and possibly also cats as a model for humans in neuroimaging studies.
Subanesthetic doses of ketamine (0.5 mg kg−1), a dissociative anesthetic acting as an antagonist at the N-methyl-d-aspartate-receptor, can be used as a rapid-acting antidepressant in patients with treatment-resistant depression (14–17). Rapid antidepressant effects have also been reported when atypical antipsychotics, with 5-HT2A antagonistic properties, were administered in combination with selective serotonin reuptake inhibitors in patients with major depression (18,19). Our hypothesis was that ketamine possibly exerts its antidepressant action through interaction with 5-HT2A receptors, which are known to play an important role in the pathogenesis of major depressive disorders (20,21).
The aim of this study was to determine the existence and duration of a pseudoequilibrium and to study the effect of ketamine on 5-HT2A receptors in cats.
MATERIALS AND METHODS
Six healthy adult female cats (domestic shorthair; age, 6 y) with a body weight of 6.1 ± 0.78 kg (mean ± SD) were used for this study. The cats had no history of behavioral abnormalities or neurologic disorders and were used to being handled by humans. All manipulations were performed according to good animal practice, and no excitation or aggression was incited.
This study was approved by the local Ethical Committee of the Faculty of Veterinary Medicine, Ghent University (EC 2007/033).
The receptor-binding studies were performed with 123I-5-I-R91150, a tracer synthesized by electrophilic substitution on the 5-position of the methoxybenzamide group of 5-I-R91150, followed by purification with high-performance liquid chromatography. The tracer (31.0 ± 4.3 MBq kg−1) (mean ± SD) was injected intravenously into the cephalic vein 10 min after catheter placement.
Pseudoequilibrium
To determine the interval during which a pseudoequilibrium state occurs, anesthesia was induced 60 min after tracer injection with propofol (6–10 mg kg−1, Propovet; Abbott Animal Health), an intravenous anesthetic interacting with γ-aminobutyric acid-A receptors (22), followed by a continuous-rate infusion of propofol (0.22 mg kg−1 min−1) for maintenance of anesthesia. Four consecutive acquisitions of 20 min each were performed, starting at 70, 90, 110, and 130 min after tracer injection.
Condition Ketamine (Condition K)
To study the effect of ketamine on the 5-HT2A receptor status, 2 acquisitions of 20 min each were performed, starting at 90 and 130 min (Table 1). Therefore, anesthesia was induced at 75 min after tracer injection. Induction and maintenance were done with propofol, as described for the pseudoequilibrium study. After the first acquisition (condition propofol, or condition P), at 110 min after tracer injection, ketamine (Anesketin; Eurovet), 5 mg kg−1, was injected intravenously, followed by a continuous-rate infusion of the combination of propofol (0.22 mg kg−1 min−1) and ketamine (23 μg kg−1 min−1). The second acquisition (condition K) was subsequently started at 130 min after tracer injection.
Scanning Protocol for Studying Influence of Ketamine on 5-HT2A Receptor Status in Feline Brain
All acquisitions for both studies were thus performed under general anesthesia, during which the cats were intubated after the larynx had been sprayed with lidocaine (Xylocaine 2%; Astra Zeneca) (3.5-mm internal-diameter endotracheal tube; Cook Veterinary Products), received intravenous fluids at a rate of 10 mL kg−1 h−1 (NaCl 0.9%; Braun Medical), and were allowed to breathe 100% oxygen using a commercial pediatric circle anesthetic system (Narkose Spiromat 650; Dräger). Ventilation was controlled with intermittent positive pressure ventilation with a tidal volume of 10–20 mL kg−1, and the respiration frequency was adjusted to maintain the end-tidal carbon dioxide concentration at 40 mm Hg.
During the acquisition, the end-tidal CO2 concentration, inspiratory oxygen fraction, and peripheral hemoglobin saturation and heart rate were measured using a calibrated (Quick Cal Calibration Gas; GE Healthcare) multigas analyzer (Capnomac Ultima; Datex) and a pulse oximeter (N-20PA portable pulse oximeter; Nellcor Puritan Bennett). Systolic arterial blood pressure was measured during the acquisition using an ultrasonic Doppler flow detector (Parks Medical Electronics) and a neonatal inflatable cuff with a width approximately 40% of the tail circumference.
Acquisition
During all acquisitions, the cats were positioned in ventral recumbence. The detectors of the triple-head γ-camera (Triad; Trionix) were positioned as close as possible to the cat’s head (range of radius of rotation, 220 mm). To minimize interindividual positioning variability, a preformed head-resting cushion was used. The height of the imaging table, the radius of rotation, and the depth position of the cat’s head were noted for each subject, and the same parameters were used for all the acquisitions. The triple-head γ-camera was equipped with 3 multipinhole collimators (6 multifocused holes 3 mm in diameter) (HiSPECT; Bioscan).
Data were acquired in step-and-shoot mode (10 steps, 36° angular step, 120 s/step), resulting in an acquisition time of 20 min. Images were reconstructed with a dedicated iterative program for HiSPECT (9 iterations, 6 subsets) into transaxial images of 1.0-mm matrix size.
Image Analysis
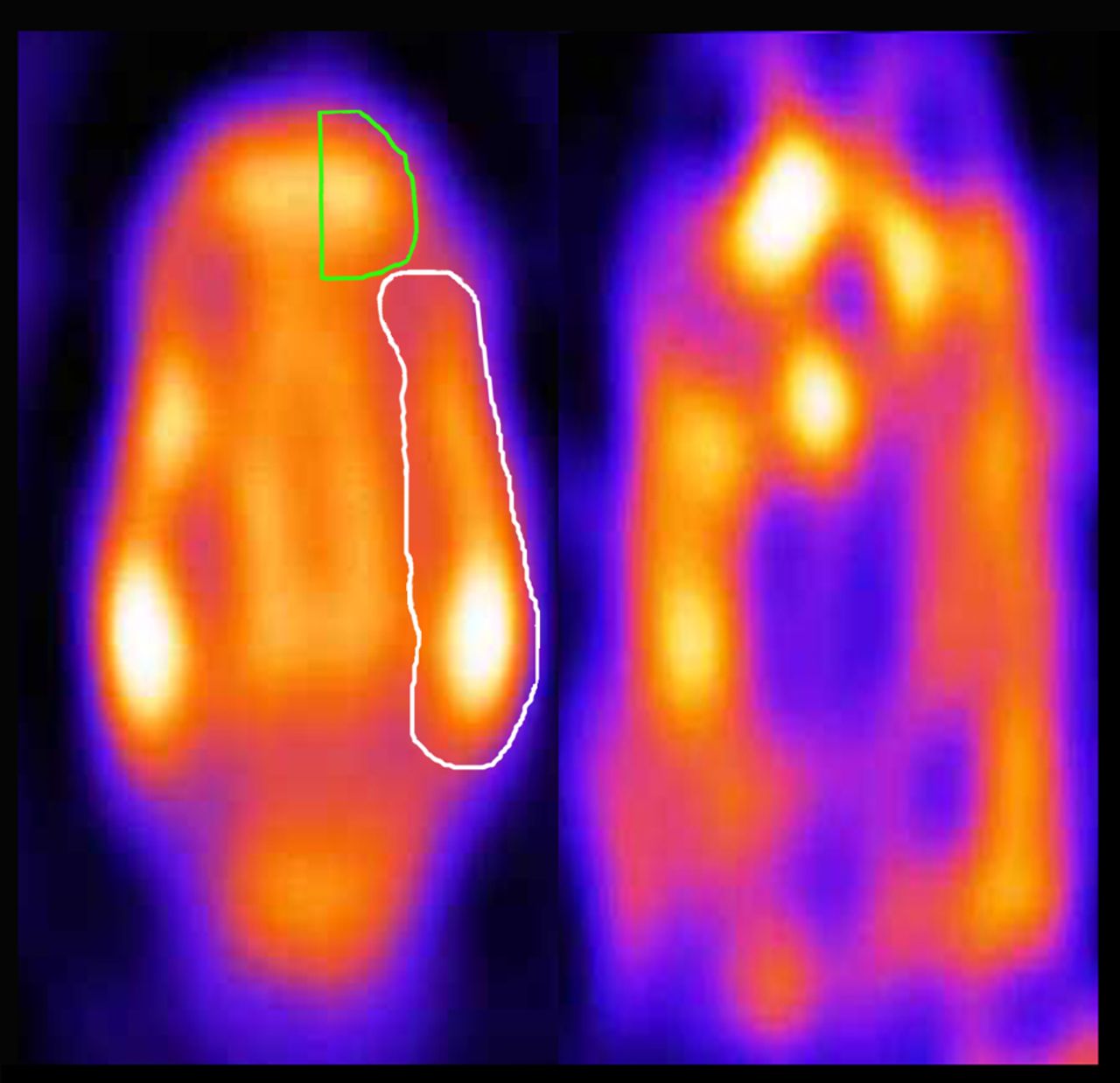
Because of the poor anatomic definition of neuroreceptor imaging studies, the 5-HT2A receptor ligand study data were matched with perfusion data obtained during a previously performed SPECT study (Fig. 1). In that study, 99mTc-ethylcysteinate dimer was used to investigate the influence of anesthetics on the regional perfusion of the brain in the same animals as were used in this study (23). Although small differences can be accounted for by the software, the same settings and configuration were used in both experiments. The multimodality software (version 5.0) from NUD, displaying images in a dual window setting and allowing for manual coregistration by providing tools for scaling, rotating, and translating images in 3 dimensions, was used to perform this fitting procedure. In this study, a region-of-interest analysis was performed, with the regions of interest (left and right frontal cortices, left and right temporal cortices, and cerebellum) defined on the perfusion images placed on the 5-HT2A receptor data. The uptake in the cerebellum was used as a reference region (24).
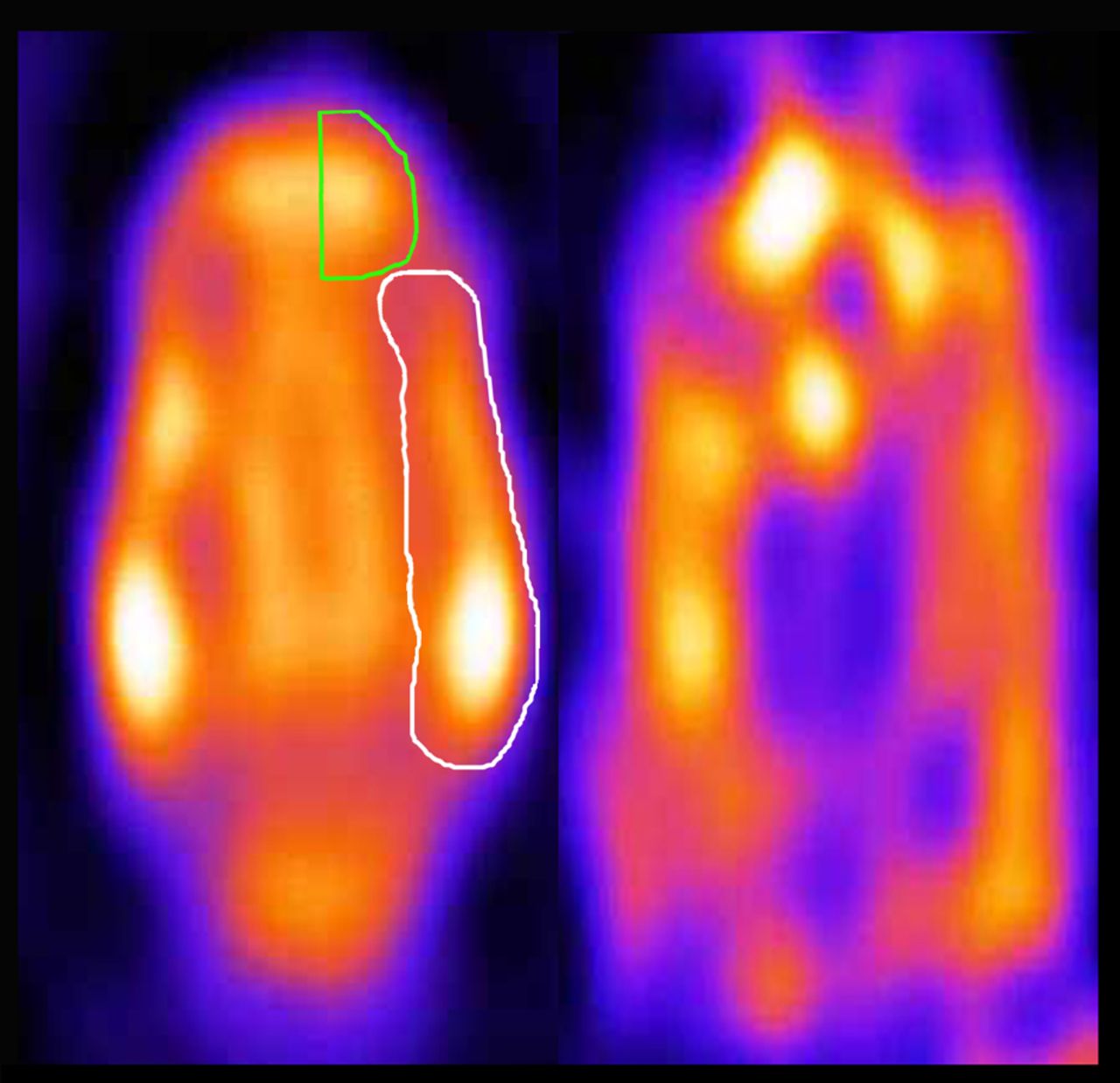
Regional perfusion (left side) and 5-HT2A receptor status (right side) in feline brain. Images were obtained with micro-SPECT and represent horizontal slice (not summed, averaged, or normalized) of individual cat. 5-HT2A receptor ligand study data were matched with perfusion data obtained during previously performed SPECT study. Unilateral regions of interest (frontal cortex in green, temporal cortex in white) were manually placed on perfusion image on the basis of 7-T MR images and cat brain atlas (23).
The existence and duration of a pseudoequilibrium were assessed by comparing the counts registered in the frontal cortex with those registered in the cerebellum as described earlier in dogs (Fig. 2) (1).

Graphic illustration of frontocerebellar ratio (mean ± SD) in function of time. Acquisition (duration, 20 min) started at 70, 90, 110, and 130 min after tracer (123I-5-I-R91150) injection in cat (n = 6), measured with micro-SPECT. This graph shows occurrence of constant frontocerebellar ratio (pseudoequilibrium state).
The binding index, which is proportional to the in vivo density of available receptors under pseudoequilibrium conditions, was calculated as ([counts per pixel in regional cortex] − [counts per pixel in cerebellum])/[counts per pixel in cerebellum]) under conditions P and K.
Statistical Analysis
The regional BIs under both conditions were analyzed using Wilcoxon signed ranks statistics. Within both conditions, left–right differences were also analyzed using the Wilcoxon signed ranks test. A value of P less than 0.05 was considered statistically significant. Since the 5-HT2A receptor binding studies cannot be performed on awake animals, condition P was chosen as baseline and condition K was compared with this baseline condition. Because the acquisitions took 30 min, the cardiovascular parameters were recorded every 10 min for each animal during each acquisition. Therefore, the mean values were calculated and analyzed using Wilcoxon signed ranks statistics.
The Statistical Package for Social Sciences (SPSS, version 17; SPSS Inc.) was used to perform the analyses.
RESULTS
During the 4 acquisitions started at 70, 90, 110, and 130 min after tracer injection, a pseudoequilibrium was present, as demonstrated by the constant frontocerebellar ratio (Fig. 2).
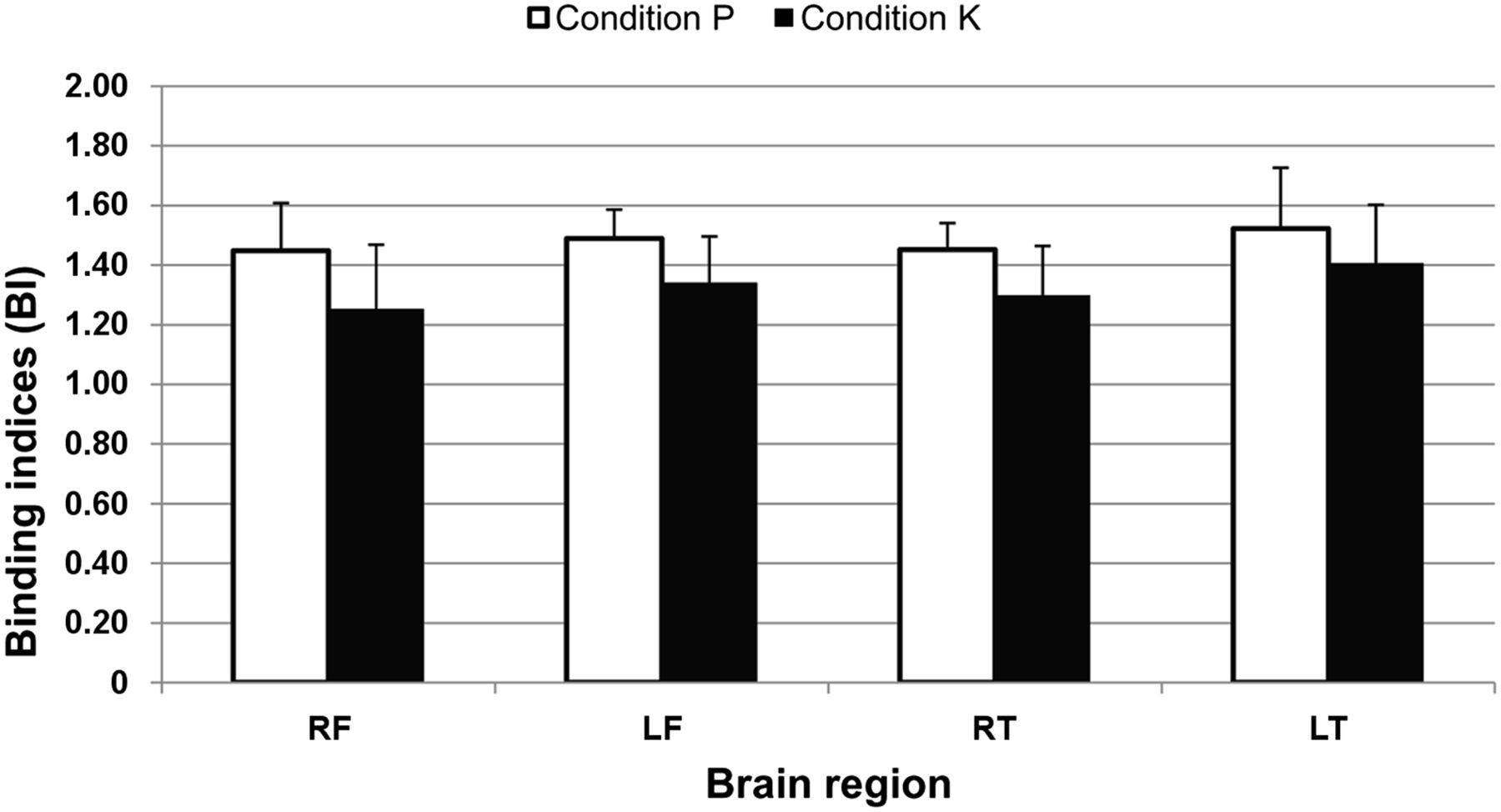
The addition of ketamine to the propofol continuous-rate infusion resulted in decreased binding indices in the right frontal cortex (1.25 ± 0.22 vs. 1.45 ± 0.16; P = 0.028), left frontal cortex (1.34 ± 0.15 vs. 1.49 ± 0.10; P = 0.028), right temporal cortex (1.30 ± 0.17 vs. 1.45 ± 0.09; P = 0.046), and left temporal cortex (1.41 ± 0.20 vs. 1.52 ± 0.20; P = 0.046) (Fig. 3).
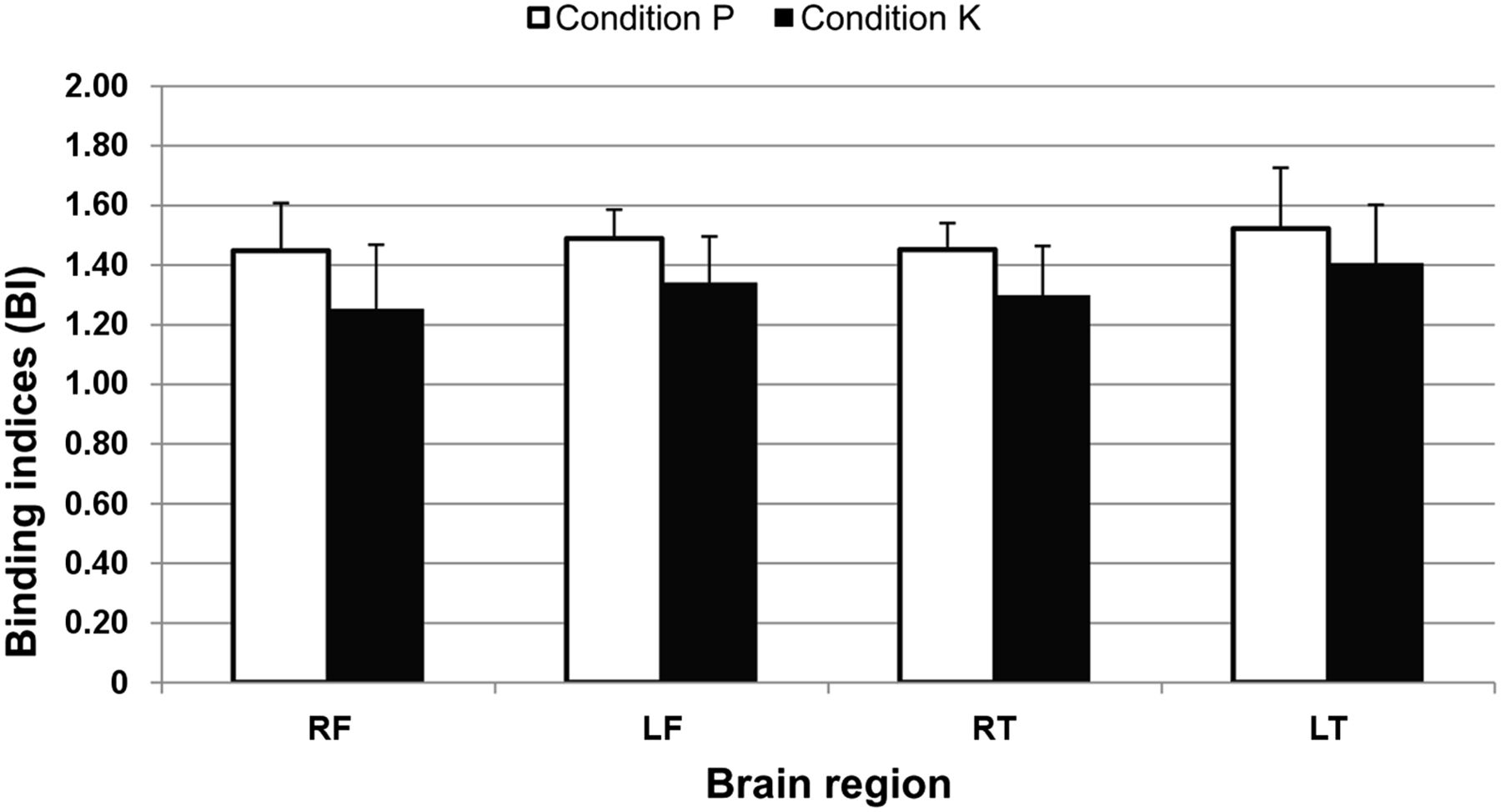
Effect of administration of ketamine on 5-HT2A binding indices (mean + SD) in feline brain. Addition of ketamine (condition K) to propofol continuous-rate infusion (condition P) caused significantly decreased 5-HT2A receptor availability in right and left frontal cortices (RF and LF, respectively) and in right and left temporal cortices (RT and LT, respectively).
The data concerning cardiovascular function during the acquisition are presented in Table 2. The addition of ketamine to the propofol continuous-rate infusion did not result in significant cardiovascular changes.
Cardiovascular Parameters During Both Acquisitions
DISCUSSION
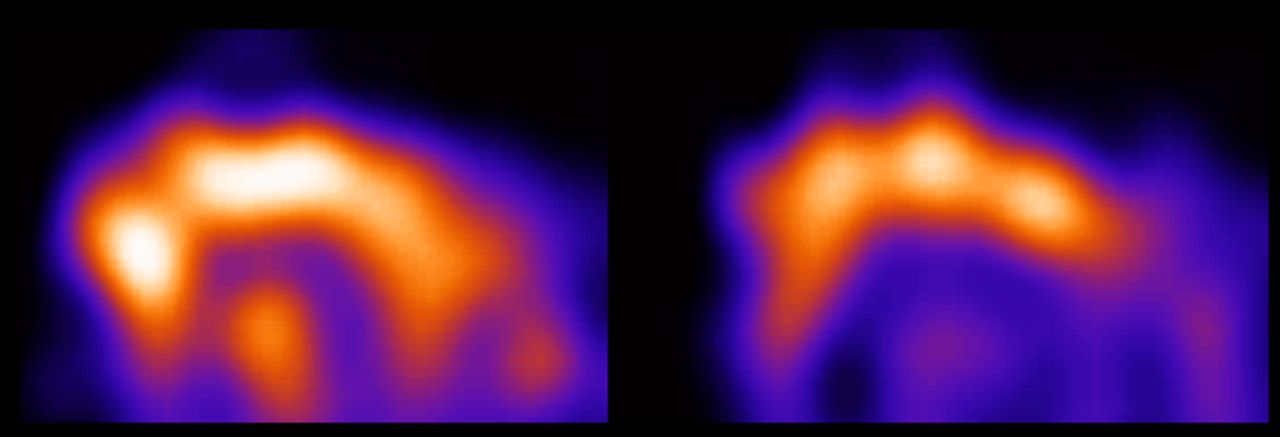
To investigate the availability of 5-HT2A receptors, 123I-5-I-R91150 SPECT can be used. The tracer uptake in the cortical areas represents free and nonspecifically bound ligand combined with ligand specifically bound to the 5-HT2A receptors. The measured radioactivity in the cerebellum, on the other hand (a region void of 5-HT2A receptors), represents only free and nonspecifically bound tracer (4,24). Semiquantification consists of the comparison of the tracer uptake in different regions of interest with the cerebellar uptake, resulting in a BI for the different regions of interest. This BI is directly related to the in vivo 5-HT2A receptor density and affinity (25). Semiquantification requires the presence of a pseudoequilibrium situation of tracer wash-in and wash-out in the regions under investigation. The interval during which a pseudoequilibrium is reached has been determined in humans, nonhuman primates, and dogs but not, to our knowledge, in cats (1–4,26,27). Therefore, before comparing BIs during different paradigms, we performed 4 consecutive acquisitions, starting at 70, 90, 110, and 130 min after tracer injection. The results of this study confirmed the presence of a stable frontocerebellar ratio during this interval (Fig. 2). It is possible that the pseudoequilibrium is already reached earlier than 70 min after injection. However, because the background noise resulting from the presence of tracer in nonbrain tissue hampered analysis of the images obtained at 70 min after injection, it seems that a shorter tracer injection–acquisition interval should not be considered in cats (Fig. 4). Although it is a bit harder to align the regions of interest perfectly on earlier acquisitions than on later acquisitions because of the higher background noise, the 70-min scan can be used for semiquantification purposes. It is possible that the pseudoequilibrium is maintained longer than 130 min after injection as well; however, this was not studied, because we believe there is no practical advantage to using a longer interval. We can thus conclude that, when 123I-5-I-R91150 micro-SPECT is used in cats to evaluate in vivo 5-HT2A receptor density and affinity, the acquisition should be started between 70 and 130 min after tracer injection.
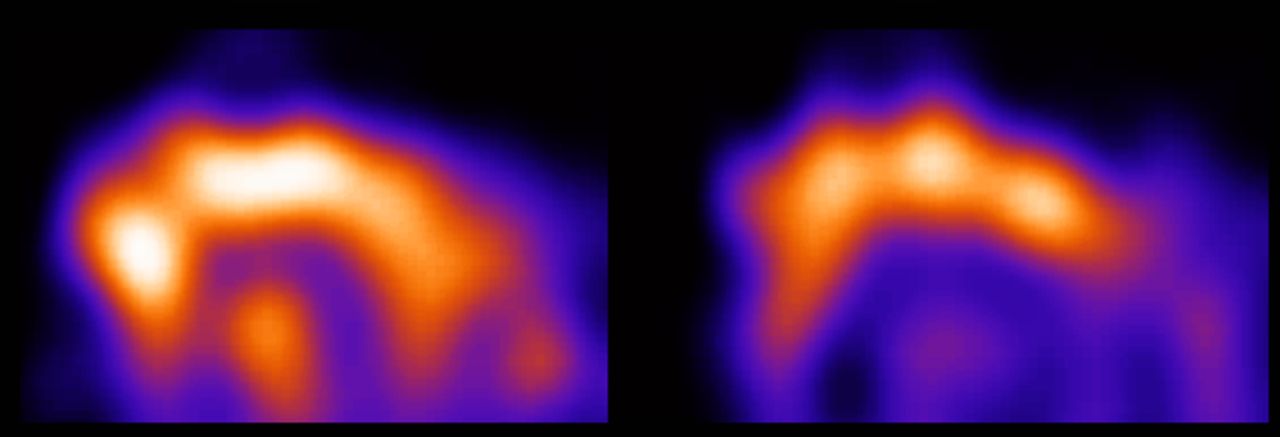
5-HT2A receptor status in feline brain at 70 min (left side) and 90 min (right side) after tracer injection. Images were obtained with micro-SPECT and represent sagittal slice (not summed, averaged, or normalized) of individual cat. Higher background noise, resulting from tracer uptake in nonbrain tissue, renders correct placement of regions of interest more difficult on image obtained at 70 min after tracer injection than on image obtained at 90 min after tracer administration.
Several authors reported a strong and rapidly occurring antidepressant effect after the administration of ketamine in patients with treatment-resistant depression (14–17). A similar effect was seen when risperidone, an atypical antipsychotic, was combined with selective serotonin reuptake inhibitors. Antagonism at the 5-HT2A receptors can possibly underlie the beneficial effect of the addition of such atypical antipsychotic drugs (28,29). Therefore, next to determining a pseudoequilibrium in cats, the main aim of this study was to look at a possible interaction between ketamine and the 5-HT2A receptors. The lower BIs under condition K than baseline condition P in the frontotemporal cortex suggest that ketamine interacts with the 5-HT2A receptors.
Lower BIs confirm an altered 5-HT2A receptor status. Four different mechanisms can possibly account for such alterations.
A higher regional cerebral blood flow can increase the amount of tracer reaching certain brain regions and thus result in a higher BI in those areas. Ketamine causes region-dependent blood flow alterations in the brain (23,30), but in a canine study those perfusion changes did not result in an altered binding of 123I-5-I-R91150 on the 5-HT2A receptors (31). Moreover, to be able to explain the decreased BIs in this study by regional blood flow alterations, the administration of ketamine should cause decreased perfusion in the frontal and temporal cortices or increased cerebellar (reference region) perfusion. Because the administration of ketamine did not result in such changes in a previous study on cats (23), the results of this study cannot be assigned to ketamine-induced perfusion changes.
N-methyl-d-aspartate receptor antagonists such as ketamine elevate the extracellular brain levels of serotonin in rodents (32). Hypothetically, as a result of competition between the tracer and serotonin, this increase can decrease the BI of 123I-5-I-R91150. Nevertheless a displacement study with the 5-HT2A receptor antagonist 3H-ketanserin showed that 5-HT antagonists were 100 to 1,000 times more potent inhibitors of the 3H-ketanserin binding at the 5-HT2 receptors than were 5-HT agonists such as serotonin itself, rendering this competition to be a theoretic explanation (33). Future brain microdialysis studies would be helpful to elucidate the effect of ketamine administration on serotonin levels in the feline brain. Acute downregulation or internalization of the 5-HT2A receptor as a consequence of increased serotonin levels in the synaptic cleft may be another option. However, Van Oekelen et al. reported that in vitro cell pretreatment with 5-HT for 15 min and 48 h did not provoke reduced 5-HT2A receptor numbers (34). This finding pleads against downregulation or internalization of the 5-HT2A receptor by a ketamine-invoked release of 5-HT. In summary it seems unlikely that the decreased BIs in this study resulted from a ketamine-induced increase in extracellular serotonin.
A third possibility is the existence of a direct competition between ketamine itself and the neuroreceptor ligand. Dose-dependent competition between 123I-5-I-R91150 and pipamperone, an antipsychotic agent acting by blocking the dopamine-2 receptors, was reported in dogs (27). A similar effect caused by ketamine can possibly account for the decreased BIs seen in this study.
Finally, ketamine can alter the binding of 123I-5-I-R91150 by decreasing the affinity of the 5-HT2A receptors. Similar alterations were reported for dopaminergic neurotransmission in the monkey brain (35,36). In those studies, ketamine facilitated the dopamine transporter, measured with 11C-β-CFT, while simultaneously decreasing the binding of the neuroreceptor ligand 11C-raclopride at the dopamine receptor. However, a distinction between direct competition and altered receptor affinity cannot be made with the technique used in this study.
This study had some limitations. A higher dose of ketamine (5 mg kg−1) was used than in the human studies (0.5 mg kg−1) (12–15). This higher dose was chosen because we performed a similar study on dogs that did not reveal decreased BIs after ketamine administration (2 mg kg−1) (31). Next to species differences, a dose-dependent effect of ketamine at the 5-HT2A receptors can be suggested. In future studies, lower doses should be used in cats as well. Increased BIs (baseline scans) are seen in dogs with behavioral disorders (37). The cats used in this study showed no obvious behavioral disorders, but investigation for behavioral disorders by specialists was beyond the scope of the study. In contrast to human neuroimaging studies, the use of animals requires sedation or anesthesia during the acquisition. Sedatives and anesthetics can interact with neuroreceptor ligand studies in 2 ways. Anesthesia-induced changes in the regional cerebral blood flow, possibly leading to a higher amount of free or nonspecifically bound ligand in certain brain regions, have been reported (23,38). Also, a direct influence on serotonergic neurotransmission by several anesthetics has been reported (39,40). In this study, propofol was chosen for induction and maintenance of anesthesia under the baseline condition (condition P), as this choice allowed us to use a single-drug anesthetic protocol, which could be combined with the drug under investigation: ketamine (condition K). Although propofol possibly interferes with the serotonergic and the N-methyl-d-aspartate-receptor system through its action at the GABAergic (γ-aminobutyric acid–mediated) neurotransmission (22), the changes seen in the BIs can be caused only by ketamine, as propofol was administered at the same rate under both conditions. The lack of randomization—with condition P always preceding condition K—is another limitation of this study. To lower the radiation burden and because of the cost of the tracer, acquisitions under both conditions were combined. The possible long-lasting effects at the 5-HT2A receptor caused by ketamine precluded randomization of both conditions. The constant frontocerebellar ratio observed in the pseudoequilibrium study under propofol anesthesia (acquisitions started between 70 and 130 min after tracer injection), however, proves that the changes in the BIs resulted from the administration of ketamine and cannot be attributed to the longer tracer administration–acquisition interval under condition K than under condition P.
CONCLUSION
This study showed that the 5-HT2A receptor status of cats can be studied with 123I-5-I-R91150 micro-SPECT and that, under propofol anesthesia, a pseudoequilibrium exists when the acquisition is started between 70 and 130 min after tracer injection. Furthermore, an interaction between ketamine and the 5-HT2A receptors resulting in decreased binding of 123I-5-I-R91150 in the frontal and temporal cortices was demonstrated. Whether this interaction causes the antidepressant action of ketamine reported in humans remains to be elucidated.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734. No potential conflict of interest relevant to this article was reported.
Footnotes
Published online Jul. 2, 2013.
- © 2013 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication September 21, 2012.
- Accepted for publication March 12, 2013.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.