


Abstract
99mTc-labeled dimercaptosuccinic acid (99mTc-DMSA) accumulates in the kidney cortex and is widely used for imaging of the renal parenchyma. Despite its extensive clinical use, the mechanism for renal targeting of the tracer is unresolved. Megalin and cubilin are cooperating receptors essential to the proximal tubule endocytic uptake of proteins from the glomerular ultrafiltrate. We have used megalin/cubilin-deficient mice produced by gene knockout to determine whether receptor-mediated endocytosis is responsible for the renal uptake of 99mTc-DMSA. Methods: Control or megalin/cubilin-deficient mice were injected intravenously with 0.5 MBq of 99mTc-DMSA or 99mTc-mercaptoacetyltriglycine (MAG3). Whole-body scintigrams and the activity in plasma, urine, and the kidneys were examined 6 h after injection. The size and identity of 99mTc-DMSA–bound proteins in urine were analyzed by fractionation by centrifugation and separation by sodium dodecyl sulfate polyacrylamide gel electrophoresis, followed by autoradiography and mass spectrometry. Results: No renal accumulation of 99mTc-DMSA was identified in scintigrams of megalin/cubilin-deficient mice. The renal accumulated activity of the tracer was reduced to 11.4% (±2.5%, n = 7) of the normal uptake in control mice, correlating with a reduction in renal megalin/cubilin expression in knockout mice to about 10% of normal. The reduced renal uptake in megalin/cubilin-deficient mice was accompanied by an increase in the urinary excretion of 99mTc-DMSA. Size separation of the urine by ultracentrifugation and sodium dodecyl sulfate polyacrylamide gel electrophoresis demonstrated that in megalin/cubilin-deficient mice an increased amount of 99mTc-DMSA was excreted in an approximately 27-kDa form, which by mass spectrometry was identified as the plasma protein α1-microglobulin, an established megalin/cubilin ligand. Conclusion: 99mTc-DMSA is filtered bound to α1-microglobulin and accumulates in the kidneys by megalin/cubilin-mediated endocytosis of the 99mTc-DMSA protein complex. Renal accumulation of 99mTc-DMSA is thus critically dependent on megalin/cubilin receptor function and therefore is a marker of proximal tubule endocytic activity.
Dimercaptosuccinic acid (DMSA) labeled with 99mTc is a major renal cortical imaging agent used in the diagnosis of renal parenchymal disorders (1). 99mTc-DMSA is highly accumulated in the kidney cortex (2), but the mechanism for the renal targeting of the tracer has not been established. Two main routes of 99mTc-DMSA tubular uptake have been proposed: by basolateral uptake from plasma by peritubular extraction and by tubular reabsorption from the glomerular ultrafiltrate. However, the significance of both routes is debated (3–12). The sodium-dependent dicarboxylate transporter NaDC-3 has been implicated in the basolateral uptake of 99mTc-DMSA from peritubular capillaries into proximal tubule cells (13). The mechanism by which 99mTc-DMSA may be reabsorbed from the glomerular ultrafiltrate is unknown; however, studies have indicated that this route may contribute substantially to the renal uptake of the tracer (4,5,14). The potential importance of this pathway is further stressed by recent observations of a decreased renal uptake of 99mTc-DMSA in patients with proximal tubular endocytic dysfunctions, such as Dent's disease, Lowes syndrome, and Fanconi syndrome (10,15,16).
The cooperating receptors megalin and cubilin are responsible for the endocytic uptake of proteins from the glomerular ultrafiltrate in the proximal tubule (17,18). Megalin, a member of the low-density lipoprotein receptor family, is a 600-kDa transmembrane glycoprotein with more than 50 known ligands, including a variety of plasma proteins. Megalin functions in a complex with the 460-kDa extracellular protein cubilin. Cubilin also binds several plasma proteins, including albumin, but depends on megalin for the endocytosis of cubilin–ligand complexes (19,20). Accordingly, megalin/cubilin-deficient mice excrete several low-molecular-weight proteins in the urine, including β2-microglobulin, α1-microglobulin, and retinol-binding protein, characteristic of tubular proteinuria caused by proximal tubule endocytic dysfunction (20–22). Megalin/cubilin dysfunction has been implicated in the pathogenesis of various renal diseases, such as the early stages of diabetic and hypertensive nephropathy, and other conditions of proteinuria (23). However, there are currently no assays for direct quantitation and imaging of proximal tubule receptor–mediated endocytic function.
Our aim was to investigate if the megalin/cubilin receptor complex is important for renal 99mTc-DMSA accumulation. Using conditional megalin/cubilin-deficient mice and examining the renal uptake and urinary excretion of tracer, we identified a crucial role of the megalin/cubilin receptors for the accumulation of 99mTc-DMSA and suggest a mechanism of uptake involving glomerular filtration and proximal tubule endocytosis of protein-bound tracer.
MATERIALS AND METHODS
Animals
Animal breeding and experiments were performed in a certified animal facility according to provisions approved by the Danish Animal Experiments Inspectorate. The conditional megalin/cubilin-deficient mice used were Meglox/lox, Cubnlox/lox,Wnt4-Cre+ mice, where the Cre-recombinase gene is driven by the Wnt4 promoter (22). Genotyping was made on tail DNA by polymerase chain reaction as described before (20). Cre-negative littermates were used as controls. The mice were on a mixed C57BL/6-129/Svj background and 8–12 wk old at the time of use. The body weights of megalin/cubilin-deficient and control mice were 27.4 ± 4.8 g and 26.9 ± 1.8 g (P = 0.39), respectively. Megalin/cubilin-deficient mice displayed left and right kidney weights of 91.8 ± 31.5 and 112.7 ± 42.8 mg, respectively, which was not significantly different from the kidney weights of controls of 79.3 ± 20.0 and 102.2 ± 20.4 mg (P = 0.20 and P = 0.28), respectively.
Radiotracers
DMSA was prepared using a commercially available kit, Renosis DMSA (RH-Isotopagenturet). Mercaptoacetyltriglycine (MAG3) was prepared using a commercially available kit, MAG3 Technescan (DRN 4334; RH-Isotopagenturet). The tracers were labeled according to the manufacturer’s instructions by adding 1,000 MBq of freshly eluted 99mTcO4− (pertechnetate) to a final concentration of 200 MBq per mL. Radiochemical purity was above 95%.
99mTc-DMSA and 99mTc-MAG3 Uptake Studies in Mice
Megalin-/cubilin-deficient or control mice were anesthetized with isoflurane for a few minutes for the intravenous injection in the femoral vein of 0.5 MBq of 99mTc-DMSA or 99mTc-MAG3 in a total volume of 0.1 mL of saline. The mice were placed in metabolic cages for 6 h for urine collection. At the end of the 6-h urine collection, the mice injected with 99mTc-DMSA (n = 7 in each group) were anesthetized, and the remaining urine in the bladder and the kidneys was collected. A blood sample was collected into a syringe containing heparin. Blood and urine samples were centrifuged for 10 min at 1,600g. Specimens (50 μL) of plasma and urine, and the whole kidneys, were counted using a γ-counter (Packard Biosciences). Renal imaging was performed by placing megalin/cubilin-deficient mice injected with 99mTc-DMSA or control mice (n = 3 in each group) directly on the detector of a γ-camera (BrightView, Philips Healthcare) with a low-energy high-resolution collimator. Planar images were acquired over 10 min in a 256 × 256 matrix.
Size Fractionation of Urine
Urine from megalin/cubilin-deficient mice injected with 99mTc-DMSA or control mice (n = 3 in each group) was pooled, and 800 μL were applied on centrifuge filters (Amicon; Millipore) with a molecular weight cutoff at 100, 30, 10, 3, and less than 3 kDa, and separated by centrifugation at 4,000g. Samples were counted using a γ-counter before and after the fractionation.
Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Protein Visualization
Urine samples (15 μL) collected from megalin/cubilin-deficient mice injected with 99mTc-DMSA or control mice (n = 3 in each group) were separated under nonreducing conditions by SDS-PAGE (16%). For protein identification, gels were silver-stained using Proteosilver Plus according to the manufacturer’s instructions (Sigma-Aldrich) and dried, and the position of the 99mTc-labeled protein was determined by autoradiography. For immunodetection, urine samples collected from megalin/cubilin-deficient mice or controls (n = 3 in each group) were separated by unreduced SDS-PAGE (16%) and immunoblotted as described previously (24), using rabbit-antirat α1-microglobulin (Agrisera).
Mass Spectrometry (MS)
The in-gel digestion procedure of proteins separated by SDS-PAGE was performed essentially as described before (25). The samples were digested by sequence-grade porcine trypsin, peptides were recovered by reverse-phase absorption (C18 Stagetip; Proxeon Biosystems), and the sample was eluted directly onto the matrix-assisted laser desorption/ionization (MALDI) target using 1 μL of α-cyano-4-hydroxycinnamic acid in 70% acetonitrile and 0.1% trifluoroacetic acid. Peptides were subsequently analyzed using an Autoflex Smartbeam III instrument (Bruker Daltonics GmbH) operated in both MS and MS/MS mode (LIFT). Before analyses, the instrument was calibrated by external calibration using a peptide mix containing 7 calibrants (Bruker Daltonics). On the basis of the generated mass spectra, proteins were identified using the Mascot search engine and the Swiss-Prot database as reference.
Kidney Tissue Preparation and Analysis
After being counted on the γ-counter, kidney extracts were prepared from the collected kidneys of the megalin/cubilin-deficient mice injected with 99mTc-DMSA or from control mice. In brief, kidney cortices were homogenized in ice-cold 0.3 M sucrose, 25 mM imidazole (pH 7.2), 8.5 M leupeptin, 1 mM phenylmethylsulfonylfluoride (PMSF), and 1 mM ethylenediaminetetraacetic acid for 30 s with an Ultra-Turrax T8 homogenizer (IKA-Werke) and centrifuged at 4,000g for 15 min at 4°C. Cleared supernatants (10 μg of protein) were analyzed by SDS-PAGE (16%) and visualized by immunoblotting (24), using sheep-antirat megalin (26), rabbit-antirat cubilin (27), and rabbit antiactin (A5060; Sigma-Aldrich). Band intensities were quantified using the Odyssey software (version 1.2; LI-COR Biosciences). The megalin and cubilin protein levels in the kidneys were expressed relative to actin protein levels.
In Vitro Binding Assay
99mTc-DMSA (0.005 MBq) was incubated for 1 h at 37°C with buffer or purified human α1-microglobulin (AbD Serotec) at a final concentration of 0.05 mg/mL, in a total volume of 250 μL diluted in phosphate-buffered saline (pH 7.4). Samples (50 μL) were counted with a γ-counter before and after fractionation on a filter (molecular weight cutoff, 30 kDa; Amicon) by centrifugation at 4,000g. The experiment was performed in triplicate.
Immunohistochemistry
Kidneys from megalin/cubilin-deficient or control mice (n = 3 in each group) were fixed by retrograde perfusion with 2% paraformaldehyde, and sections were processed as reported before (20), using sheep-antirat megalin and rabbit-antirat α1-microglobulin. Images were obtained using a confocal laser-scanning microscope (LSM 510-META; Carl Zeiss) and processed using Zeiss Zen software (2009, Light Edition).
Statistics
Data represent mean ± SD. Groups were compared using a Student unpaired t test, assuming equal variance. A P value of less than 0.05 was considered significant.
RESULTS
Renal Uptake and Urinary Excretion of 99mTc-DMSA
Accumulation of 99mTc-DMSA uptake in the kidneys of normal control mice was evident by whole-body γ-camera scintigraphy 6 h after intravenous injection of 99mTc-DMSA (Fig. 1A). In contrast, no accumulation was seen in the kidneys of megalin/cubilin-deficient mice (Fig. 1B). When quantified by γ-counting, the uptake of 99mTc-DMSA in isolated kidneys from megalin/cubilin-deficient mice was reduced to 1.8% ± 0.4% of the injected dose per 100 mg of kidney tissue (Fig. 2A), corresponding to 11.4% of the normal uptake in controls (n = 7 in each group). This uptake was associated with an approximate 90% increase in the urinary excretion of 99mTc-DMSA (Fig. 2B). Thus, whereas the total renal activity of megalin/cubilin-deficient mice was 8,882 ± 3,264 cpm, compared with 66,656 ± 9,336 cpm (P < 0.0001) in controls, the total activity of 99mTc-DMSA extracted by the kidneys (kidney plus urine activity) was not significantly different between megalin/cubilin-deficient mice and control mice (7.9⋅103 ± 1.6⋅103 vs. 6.5⋅103 ± 1.2⋅103cpm/g mouse, P = 0.07).

Megalin/cubilin-dependent renal accumulation of 99mTc-DMSA. Whole-body scintigram of mice 6 h after injection of 99mTc-DMSA shows accumulation in kidneys of control mice (A) but no accumulation in kidneys of megalin/cubilin-deficient mice (B). In megalin/cubilin-deficient mice, evidence of accumulation of tracer in bladder was seen. Images shown are representative of similar analyses (n = 3 in each group).

Renal uptake and urinary excretion of 99mTc-DMSA by megalin/cubilin-deficient and control mice. Renal uptake (A), urinary excretion (B), and plasma levels (C) of 99mTc-DMSA in megalin/cubilin-deficient mice are compared with controls. Columns represent mean of each group (n = 7) ± SD. *Significantly different from control mice (P < 0.0001).
The approximate 90% reduction in kidney uptake of the tracer correlates with a megalin and cubilin protein knockdown efficiency of 90.8% ± 8.7% and 89.8% ± 6.1%, respectively, as evaluated by immunoblotting of kidney cortex homogenates from megalin/cubilin-deficient mice, compared with controls, which is similar to previous reports in these mice (22). The plasma level of 99mTc-DMSA in megalin/cubilin-deficient mice and controls was not significantly different (Fig. 2C).
Urinary Excretion of 99mTc-MAG3
No significant difference (P = 0.11) in the urinary excretion of 99mTc-MAG3 was observed between megalin/cubilin-deficient (n = 7) and control mice (n = 7, Fig. 3) after 6 h. Although we cannot rule out a potential difference in the kinetic parameters of 99mTc-MAG3 handling, the total excretion of 99mTc-MAG3 appears to be unaffected by the megalin/cubilin deficiency.

Urinary excretion of 99mTc-MAG3 by megalin/cubilin-deficient and control mice. Columns represent mean of each group (n = 7) ± SD.
Urinary Excretion of Protein-Bound 99mTc-DMSA
The molecular weight of the 99mTc activity in urine was determined through size fractionation by successive application of urine to centrifuge filters, thereby producing fractions corresponding to molecular weights of more than 100, 30–100, 10–30, 3–10, and less than 3 kDa. In urine from megalin/cubilin-deficient mice, 60% of the injected dose of 99mTc-DMSA was associated with fractions corresponding to a molecular weight of more than 10 kDa, compared with only 17% in urine from control mice (Table 1). In both controls and megalin/cubilin-deficient mice, 2%–3% was bound to small peptides (3–10 kDa), and 19%–20% was excreted as small molecules, including amino acid–bound and free 99mTc-DMSA. Thus, the increased 99mTc-DMSA activity in the urine of megalin/cubilin-deficient mice reflects a 3.5-fold increase of protein-bound 99mTc-DMSA excreted in the urine.
Ultrafiltration of Urine from Mice Injected with 99mTc-DMSA
Identification of 99mTc-DMSA Bound to α1-Microglobulin
The analysis of urine samples collected from megalin/cubilin-deficient mice by SDS-PAGE and autoradiography showed that the 99mTc radioactivity was associated with a single band of approximately 27 kDa (Fig. 4A). To identify the protein associated with the tracer, the collected urine samples were subjected to SDS-PAGE, and proteins were visualized by silver staining. The sample representing megalin/cubilin-deficient mice presents unique staining at the position corresponding to 27 kDa (Fig. 4B). This band was excised, subjected to in-gel digestion using trypsin, and subsequently identified as the plasma protein α1-microglobulin by MALDI MS (Table 2). This finding was further corroborated by immunoblotting (Fig. 4C). The interaction between 99mTc-DMSA and α1-microglobulin was furthermore evaluated in vitro using purified α1-microglobulin, whereby approximately 80% of the 99mTc-DMSA activity was found bound to α1-microglobulin after 1 h of incubation. Finally, immunohistochemical staining of kidney sections from control and megalin/cubilin-deficient mice demonstrated proximal tubule uptake of α1-microglobulin in megalin-expressing cells only, confirming that megalin is essential for uptake of α1-microglobulin (Fig. 5).
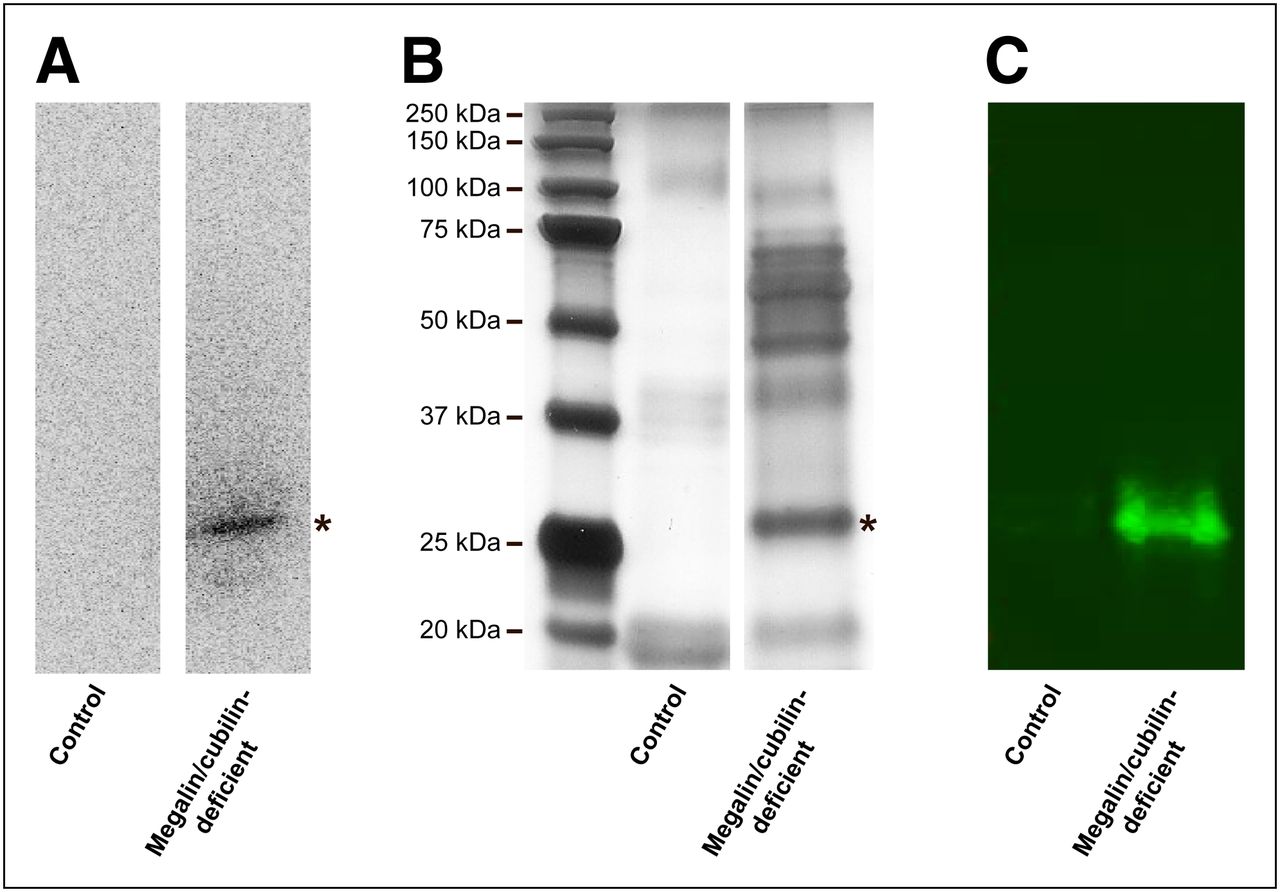
Urinary excretion of protein-bound 99mTc-DMSA by megalin/cubilin-deficient mice. (A) SDS-PAGE separation and autoradiography of urine from megalin/cubilin-deficient mice and controls injected with 99mTc-DMSA. A single, labeled band of approximately 27 kDa is observed in urine collected from megalin/cubilin-deficient mice only. (B) Urine profile analysis by SDS-PAGE and silver staining of urine from megalin/cubilin-deficient mice and controls. A distinct protein band of 27 kDa was identified in urine from megalin/cubilin-deficient mice (*). This band was excised and protein identified by MALDI MS as α1-microglobulin (Table 2). (C) Immunoblotting identifying α1-microglobulin as an approximate 27-kDa band in urine from megalin/cubilin-deficient mice but not from controls. Data shown are representative of 2 (autoradiography and MALDI MS) or 3 (SDS-PAGE, silver staining, and immunoblotting) independent experiments.
Mass Spectrometry (MS/MS) Analysis of Trypsin-Cleaved 27-kDa Band from Urine Separated by SDS-PAGE

Megalin/cubilin-dependent renal uptake of α1-microglobulin. Immunohistochemical staining is shown for megalin (green) and α1-microglobulin (red) in kidney cortex of control and megalin/cubilin-deficient mice. In megalin/cubilin-deficient mice, a few profiles of megalin-expressing proximal tubule cells can be identified (*) because of mosaic expression. Merged images showed that α1-microglobulin cannot be identified in proximal tubular cells lacking megalin.
DISCUSSION
We show that in megalin/cubilin-deficient mice, the renal accumulation of 99mTc-DMSA is almost abolished and associated with a corresponding increase in urinary excretion of the tracer. The increased 99mTc-DMSA activity in the urine is found to be associated with high-molecular-weight fractions, suggesting protein binding. MS and in vitro binding studies identified α1-microglobulin as the 99mTc-DMSA binding protein present in urine. α1-microglobulin is a 27-kDa plasma protein filtered in the glomeruli and an established ligand for both megalin and cubilin (21,24). This is confirmed by immunohistochemistry showing uptake of α1-microglobulin in megalin-expressing tubule cells only. On the basis of these observations, we proposed a new model for renal accumulation of 99mTc-DMSA (Fig. 6). In this model, 99mTc-DMSA binds to α1-microglobulin, which is filtered as a complex in the normal renal glomeruli followed by megalin/cubilin-mediated endocytic uptake and accumulation of the tracer in the kidney proximal tubules.

Mechanism for renal uptake of 99mTc-DMSA. (A) α1-microglobulin–bound 99mTc-DMSA is freely filtered by glomeruli and accumulates in renal proximal tubules by endocytosis mediated by multiligand-binding megalin/cubilin receptors (B). (C) Free 99mTc-DMSA and trace amounts of α1-microglobulin–bound 99mTc-DMSA are excreted in urine. Consequently, megalin/cubilin dysfunction leads to abolishment of renal uptake and increased urinary excretion of α1-microglobulin–bound 99mTc-DMSA.
An approximate 10% residual renal uptake of 99mTc-DMSA was observed in the kidneys from megalin/cubilin-deficient mice. Although an additional and minor pathway for renal uptake of 99mTc-DMSA cannot be excluded, this corresponds to the possibility that the approximately 10% remaining megalin expression in these mice is a likely explanation for the residual uptake. This argument is also consistent with a previous study using kidney-specific megalin-deficient mice, in which a renal uptake of 35%–55% of 99mTc-DMSA was reported in megalin-deficient mice, compared with normal controls, although the data were not shown in the study (12). Previous analyses, however, showed that these megalin-deficient mice have a partial and variable (35%–50%) megalin expression in the kidneys (28), explaining this variation in 99mTc-DMSA uptake.
The renal 99mTc-DMSA uptake in megalin/cubilin-deficient mice was reduced to 1.8% ± 0.4% of the total injected dose per 100 mg of kidney tissue, whereas the uptake in control mice was 15.6% ± 2.3%, similar to what has been reported in rats (12). Importantly, the total activity of 99mTc-DMSA (7.9⋅103 ± 1.6⋅103cpm/g) extracted by the kidneys (kidney plus urine activity) of megalin/cubilin-deficient mice was not significantly different from that of control mice (6.5⋅103 ± 1.2⋅103cpm/g) (P = 0.07), indicating similar renal extraction and filtration of 99mTc-DMSA.
α1-microglobulin, a member of the lipocalin protein superfamily, is synthesized in the liver, freely filtered by glomeruli, and reabsorbed by renal proximal tubule cells, where it is catabolized (29). Under normal conditions, little filtered α1-microglobulin appears in the final excreted urine; however, urinary excretion of α1-microglobulin has been demonstrated previously in megalin-deficient mice (21) and α1-microglobulin was recently shown to be excreted in the urine of a patient with cubilin dysfunction, where additional binding experiments confirmed that both megalin and cubilin bind α1-microglobulin (24). An increase in the urinary concentration of α1-microglobulin therefore indicates proximal tubule cell injury or impaired proximal tubular endocytosis and is considered a valuable marker of renal proximal tubular dysfunction (30–32). Interestingly, similar to our findings in urine from megalin/cubilin-deficient mice, the molecular size of excreted 99mTc-DMSA in the urine of patients with idiopathic tubular proteinuria is also in the range of 24–28 kDa (33), supporting binding to a urinary protein of this size. Our study showed no uptake of α1-microglobulin in megalin-negative tubular cells independent of cubilin expression. Thus, although α1-microglobulin may bind to both cubilin and megalin, megalin is essential for the internalization of the cubilin–α1-microglobulin complex, as also shown for other proteins binding to both receptors, such as albumin (20). It has previously been shown that most (65%–90%) 99mTc-DMSA in plasma is protein-bound (6,31,34), and it has been proposed that the sulfhydryls in DMSA bind to free cysteine residues. α1-microglobulin has a free cysteine side-chain located in a flexible loop, giving the protein reductase and dehydrogenase properties with a broad biologic substrate specificity (35). It is therefore likely that the binding of 99mTc-DMSA to α1-microglobulin occurs via its free cysteine residue.
On the basis of our findings, the renal uptake of 99mTc-DMSA depends on the binding to α1-microglobulin, whereby it becomes a ligand for the megalin/cubilin receptors; the glomerular filtration of α1-microglobulin; and the activity of proximal tubule megalin/cubilin-mediated endocytosis. Under conditions for which the first 2 factors may be assumed to be constant, a 99mTc-DMSA scan will reflect the receptor-mediated endocytic activity in the proximal tubule, as is supported by several studies showing poor 99mTc-DMSA renal uptake in patients with disturbed tubular endocytosis and subsequent low-molecular-weight proteinuria, including Dent's disease, Lowes syndrome, and Fanconi syndrome (10,15,16). Reduced 99mTc-DMSA uptake in these patients is associated with increased bladder activity, indicating an increased urinary excretion of 99mTc-DMSA, and correlates with the degree of tubular proteinuria (10). However, quantitative estimates of megalin/cubilin function will require further studies, including evaluation of the dynamics of 99mTc-DMSA uptake and relating it to the glomerular filtration rate in conditions with varying degree of tubulopathy. In the case of heavy proteinuria, an altered tracer distribution could be due to the increased glomerular filtration of proteins, leading to increased urinary loss and reduced 99mTc-DMSA uptake, as reported (36). Conversely, diminished filtration would lead to low 99mTc-DMSA kidney uptake, even while tubular function is retained, as it is observed under renal artery stenosis and captopril experiments (5,37). Thus, 99mTc-DMSA renal uptake should also be cautiously interpreted in the case of alterations in glomerular filtration.
Our findings may also explain previous observations made in a mouse model of Dent's disease caused by knockout of the renal, endosome-associated chloride channel ClC-5. In Clcn5 knockout mice, impaired uptake of 99mTc-DMSA was observed (11). The Clcn5 knockout mice are known to display disturbed megalin/cubilin-mediated endocytic activity due to impaired trafficking of the receptors (38), accompanied by an increased urinary excretion of α1-microglobulin (39). Thus, although it was proposed that CLC-5 affects trafficking of NaDC transporters (40), it seems likely that dysfunction of megalin/cubilin-mediated endocytosis is the actual cause of impaired 99mTc-DMSA uptake in these mice. Interestingly, similar to what our data indicate in megalin/cubilin-deficient mice, the renal uptake of 99mTc-MAG3 was normal in Clcn5 knockout mice (11), suggesting that the basolateral uptake mediated by an organic anion transporter (41), and its excretion by proximal tubular cells, is not disturbed by defective endocytosis.
CONCLUSION
We have shown that megalin/cubilin receptor–mediated endocytosis is essential for normal renal accumulation of the 99mTc-DMSA tracer. Renal uptake depends on the binding of 99mTc-DMSA to the plasma protein α1-microglobulin, followed by glomerular filtration and megalin/cubilin-mediated endocytosis by proximal tubular cells. Thus, the 99mTc-DMSA renography is a potential method for the evaluation of proximal tubule endocytic function in patients.
DISCLOSURE
The costs of publication of this article were defrayed in part by the payment of page charges. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Acknowledgments
The work was supported by the Lundbeck Foundation, the Danish Medical Research Council, Novo Nordisk Foundation, the European Commission EUNEFRON (FP7, GA#201590), The Danish Kidney Association, and The Aase and Ejnar Danielsens Foundation. No other potential conflict of interest relevant to this article was reported. Hanne Sidelmann, Inger Blenker Kristoffersen, Pia Kamuk Nielsen, Helle Salling Gittins, and Lene E. Nielsen are gratefully acknowledged for excellent technical assistance.
Footnotes
Published online Dec. 11, 2012.
- © 2013 by the Society of Nuclear Medicine and Molecular Imaging, Inc.
REFERENCES
- Received for publication June 29, 2012.
- Accepted for publication August 15, 2012.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}