


Abstract
Vesicular monoamine transporter 2 (VMAT2) is a putative molecular target for the quantitative imaging of pancreatic β-cell mass by PET. The VMAT2 PET tracer 11C-dihydrotetrabenazine (11C-DTBZ) exhibits high pancreatic uptake that is reduced in type 1 diabetes. The aim of this study was to assess the islet and VMAT2 specificity of DTBZ binding in the pancreas. Methods: The biodistribution of 11C-DTBZ in rats was determined 10 and 60 min after injection. The localization of DTBZ radioactivity in rat and human pancreatic tissue sections was investigated by autoradiography. Saturation and competition binding assays were performed with 3H-DTBZ and sections of rat pancreatic and control tissues. The binding of 11C-DTBZ in pancreatic sections from rats with streptozotocin-induced diabetes was compared with that in control rats. Results: The values for the pancreatic uptake of 11C-DTBZ (percentage injected dose per gram of tissue) were 3.0 at 10 min and 2.7 at 60 min. At 10 min, pancreatic radioactivity was heterogeneously distributed, with higher levels toward the head of the pancreas (head-to-tail ratio, 1.7). No such gradient was observed in pancreatic sections incubated with 11C-DTBZ and 3H-DTBZ in vitro. In rats, 11C-DTBZ and 3H-DTBZ binding in pancreatic islets did not exceed binding in the exocrine pancreas. Saturable 3H-DTBZ binding was observed in the rat brain striatum (dissociation constant [Kd], 1.3 nM) and the bovine adrenal medulla (Kd, 3.3 nM), whereas in the rat pancreas, 3H-DTBZ binding was nonsaturable. Competition binding with 3H-DTBZ and VMAT2 antagonists also indicated that DTBZ binding in the rat pancreas was nonspecific and did not represent binding to VMAT2. Nonspecific pancreatic 11C-DTBZ binding was lower in rats with streptozotocin-induced diabetes than in control rats. In sections of human pancreas, a subset of pancreatic islets were weakly but VMAT2-specifically labeled with 3H-DTBZ. Conclusion: The results showed that the pancreatic uptake of 11C-DTBZ is mainly due to nonspecific binding in the exocrine pancreas and suggested that the reduction in pancreatic 11C-DTBZ binding observed in type 1 diabetes is not specific for the loss of β-cell mass.
PET and SPECT methods for the imaging of congenital and adult hyperinsulinism (1,2) and pancreatic cancer, including pancreatic islet cell tumors (3,4), are already in clinical use. In addition, efforts have been made to develop PET tracers specific for pancreatic β-cells, which would provide a noninvasive method for estimating β-cell mass. The loss of pancreatic β-cells is a feature of both type 1 and advanced type 2 diabetes. At present, estimations of pancreatic β-cell integrity rely on measurements of stimulated insulin secretion capacity.
Two distinct vesicular monoamine transporters, vesicular monoamine transporter 1 and vesicular monoamine transporter 2 (VMAT2), transport biogenic amines into intracellular storage vesicles (5–7). Neurons and pancreatic β-cells accumulate monoamines by using VMAT2 (8–10), the pharmacology of which has been extensively characterized with bovine adrenal medulla chromaffin granules (11). The VMAT2 antagonist 11C-dihydrotetrabenazine (11C-DTBZ) is an established tracer for PET imaging of the dopaminergic system of the brain (12). High uptake of 11C-DTBZ radioactivity is evident in the pancreas of nondiabetic rodents. PET studies in rats showed that the uptake of 11C-DTBZ radioactivity in the pancreas declined with the onset of autoimmune and streptozotocin-induced diabetes (13,14). Moreover, the pancreatic uptake of 11C-DTBZ radioactivity was lower in a group of patients with long-standing type 1 diabetes than in a group of healthy volunteers, as determined by PET (15). Therefore, 11C-DTBZ has been considered to be a candidate molecule for the noninvasive quantitative imaging of pancreatic β-cell mass in humans (16,17). However, the localization of 11C-DTBZ radioactivity specifically to the pancreatic islets has not been verified.
This investigation was performed to determine the islet and VMAT2 specificity of DTBZ binding in rat pancreas and human pancreas. After injection of 11C-DTBZ into rats, pancreatic tissue sections were prepared, and the accumulation of radioactivity in pancreatic islets and the exocrine pancreas was assessed. The effect of type 1 diabetes on 11C-DTBZ binding in the pancreas was studied in vitro with pancreatic sections from rats with streptozotocin-induced diabetes. Saturation and competition binding assays were performed with 3H-DTBZ and sections of rat pancreatic tissues and VMAT2-rich control tissues to determine the VMAT2 specificity of pancreatic DTBZ binding. 3H-DTBZ autoradiography was used to assess binding to pancreatic islets in humans.
MATERIALS AND METHODS
Chemicals
(+)-α-11C-DTBZ (11C-DTBZ) was synthesized by 11C methylation of (+)-9-O-desmethyl-α-dihydrotetrabenazine by a previously described method (18), with minor modifications. The radiochemical purity of the final product exceeded 98%. α-2-3H-DTBZ with a specific radioactivity of 0.74 MBq/nmol was purchased from American Radiolabeled Chemicals. Chlorpromazine, haloperidol, ketanserin, lobeline, reserpine, streptozotocin, and tetrabenazine were obtained from Sigma-Aldrich Co.
Experimental Animals and Tissues from Human Patients
Sprague–Dawley male rats (200–350 g), bred and housed under standard conditions at the Central Animal Laboratory, University of Turku, Turku, Finland, were used. Bovine adrenal gland was obtained from a local slaughterhouse (Paimion Teurastamo Paimio, Finland). The animal experiments were approved by the Animal Care and Use Committee of the University of Turku and by the State Provincial Office of Southern Finland. Human pancreatic tissue was obtained from 4 patients who underwent pancreatic resection at Turku University Hospital (Turku, Finland). The patients, 3 women and 1 man, were 59 ± 12 (mean ± SD) y old, were not diabetic, and had pancreatic cystadenoma or pancreatic cystadenocarcinoma. Approval for the use of human pancreatic tissue was obtained from the Ethics Committee of the Hospital District of Southwest Finland. All patients gave written informed consent before participating in the study.
Biodistribution and Intrapancreatic Localization of 11C-DTBZ Radioactivity
Rats (n = 14) weighing 269 ± 45 g were briefly anesthetized with CO2:O2 (50:50) gas and injected via a tail vein with 11C-DTBZ at 170 ± 105 MBq/kg. The specific radioactivity at the time of injection was 155 ± 93 MBq/nmol, and the injected molecular mass was 2.1 ± 2.2 nmol/kg. At 10 min (n = 7) or 60 min (n = 7) after injection, the rats were anesthetized with CO2 gas, and blood was collected by cardiac puncture. Plasma, tissues, and organs were weighed, and their radioactivities were measured. The biodistribution of the radioactivity, corrected for radionuclide decay, was reported as the percentage injected dose per gram of tissue.
The pancreata and brains were frozen by immersion in isopentane chilled with dry ice and then were sectioned at 20 μm on a cryostat. The sections were dried under a fan and apposed to imaging plates (Fuji BAS-TR2025; Fuji Photo Film Co.) for approximately 1 h. Digital images were obtained by scanning of the imaging plates with a BAS-5000 scanner (Fuji Photo Film Co.) at a 25-μm resolution or an FLA-5100 scanner (Fuji Photo Film Co.) at a 10-μm resolution. Visual inspection of the pancreatic images suggested that at 10 min after the injection of 11C-DTBZ, the radioactivity levels were higher toward the head than toward the tail of the pancreas. The most distal 5 mm at each end of the pancreatic sections was outlined and analyzed with image analysis software (Aida 4; Raytest), and the results were reported as photostimulated luminescence per square millimeter. Three sections per rat (n = 6, 10-min time point) were evaluated. The pancreatic head-to-tail binding ratio was determined for each rat, and the results were statistically analyzed by comparing the head-to-tail ratios to hypothetical value 1.
Pancreatic sections that had been processed for autoradiography were stained with hematoxylin and eosin, and the distributions of 11C-DTBZ radioactivity and pancreatic islets were compared by visual inspection (n = 3, 10-min time point, 2–4 sections per rat). The results were documented by taking a high-magnification composite photograph of a stained section with a 10× objective on a Zeiss Axiovert 200 M microscope (Carl Zeiss Inc.) equipped with a digital camera, a motorized stage, and software for the generation of mosaic images. Pancreatic islets were identified and manually outlined on the micrographs, and the outlines were combined with the autoradiographic images by use of Photoshop CS3 (Adobe Systems Inc.).
Metabolite Analysis
A rat was injected with 11C-DTBZ at 361 MBq/kg (3.8 nmol/kg); the specific radioactivity at the time of injection was 222 MBq/nmol. The presence of radioactive metabolites in plasma, urine, liver, and pancreas 10 min after injection was assessed by thin-layer chromatography. The head, tail, and middle portions of the pancreas were analyzed separately. 11C-DTBZ added to rat plasma was used as a standard to allow identification of the parent compound. Plasma and urine proteins were precipitated with 1.25 times the sample volume of acetonitrile and removed by centrifugation. The tissue samples were homogenized in acetonitrile:H2O (30:70), and the radioactive material was purified in 2 centrifugation steps. Supernatants (10 μL) were applied to an HPTLC Silica Gel 60 plate (105547; Merck). Chloroform:methanol (85:15), 2 × 10 mL, was used as the mobile phase. The vapor space was presaturated in a twin developing chamber (Camag Twin Through Chamber; Camag) for 30 min. The plate was developed 4 cm, dried under a fan, and apposed to an imaging plate for approximately 1 h as described earlier.
Induction of Diabetes and In Vitro Binding of 11C-DTBZ in Pancreatic Sections from Diabetic and Control Rats
On day 1, 2 groups of rats, weight-matched on day 0, were fasted for 4 h. Blood glucose from a drop of blood from a tail vein was measured with an ACCU-CHEK Aviva glucose meter (Roche Diagnostics) and plasma-calibrated test strips. The 2 groups of rats were anesthetized with isoflurane, and either streptozotocin (65 mg/kg) dissolved in 0.1 M citrate buffer (pH 4.6) (n = 8) or citrate buffer alone (n = 6) was intravenously administered. Blood glucose levels and body weights were monitored regularly and compared with baseline values obtained on day 1. The control rats remained euglycemic (7.0 ± 0.7 mM on day 8), and their body weights increased from 332 ± 19 g on day 0 to 346 ± 16 g on day 8. Two of the rats in the streptozotocin group displayed only moderate elevations in blood glucose levels, and their body weights increased. These rats were excluded from the study. The rest of the rats in the streptozotocin group (n = 6) exhibited blood glucose levels exceeding 22 mM from day 3 onward, and their body weights decreased from 329 ± 21 g on day 1 to 283 ± 22 g on day 7. On day 7 (streptozotocin group) or day 8 (control group), the rats were decapitated under isoflurane anesthesia, and their pancreata were dissected and frozen. Pancreatic sections, 20 μm thick, were cut on a cryostat and stored frozen until used.
Two pancreatic sections from each animal were incubated with 2.8 nM 11C-DTBZ (at a specific radioactivity of 180 MBq/nmol at the start of incubation) in 50 mM potassium phosphate buffer (pH 7.4) at room temperature for 20 min. The sections were washed in ice-cold buffer for 10 min, rinsed in ice-cold distilled water, dried under a fan, and apposed to imaging plates (Fuji BAS-TR2025) for approximately 1 h. Whole pancreatic sections were manually outlined and analyzed as described earlier. Occasional hot-spot artifacts, not corresponding to pancreatic islets (e.g., lower left-hand corners of Figs. 3B and 3C), were excluded from the regions of interest. Results from duplicate sections were averaged, and the levels of binding in diabetic and control rats were compared.
Pharmacologic Characterization of 3H-DTBZ Binding In Vitro
3H-DTBZ saturation and competition binding assays were performed by incubating duplicate sections of rat pancreas, rat brain, and bovine adrenal gland with 3H-DTBZ and various unlabeled compounds in 50 mM potassium phosphate buffer for 2 h at room temperature. In the saturation binding experiment, sections from 1 rat and 1 bull were incubated with 11 concentrations of 3H-DTBZ, ranging from 0.38 to 47.3 nM. Adjacent sections were incubated in the presence of an excess of a VMAT2 antagonist (100 μM ketanserin) to determine nonspecific binding. The competition binding experiment was performed with 1.8 nM 3H-DTBZ and 7 concentrations of unlabeled compounds, ranging from 0 nM to 0.1 mM. The pancreata from 2 rats, the brain from 1 rat, and the adrenal glands from 1 cow and 1 bull were used in the competition binding experiment. Because similar results were obtained from the cow and the bull, their corresponding binding values were averaged.
After the incubations, the sections were washed for 20 min in ice-cold buffer, rapidly rinsed in ice-cold distilled water, and dried under a fan. A 20-min wash was found to be optimal on the basis of the 3H-DTBZ binding ratios for the rat brain striatum to the cerebral cortex and the bovine adrenal medulla to the adrenal cortex after different washing times (data not shown). The dry sections were apposed to imaging plates (Fuji BAS-TR2025) for 10 d. 3H standards (ART-123; ARC Inc.) were coexposed with the sections to allow comparisons of the results from separate experiments. 3H-DTBZ binding levels in the rat pancreas, rat striatum (bilateral readings), and bovine adrenal medulla were measured. Results from duplicate sections were averaged.
Equilibrium dissociation (Kd) and inhibition (Ki) constants were determined by least squares nonlinear regression analysis of the binding results with GraphPad Prism 5 (GraphPad Software). The saturation binding results were analyzed with simultaneous fitting of total and nonspecific binding. Kd values obtained from the saturation binding experiment were used to calculate Ki values for the unlabeled compounds applied in the competition binding experiment; for this purpose, we used the Cheng–Prusoff equation (18): Ki = IC50/(1 + [radioligand]/Kd), where IC50 is the 50% inhibitory concentration.
3H-DTBZ Binding in Human Pancreas and Insulin Immunohistochemistry Analysis
Human pancreas sections (n = 4) were frozen immediately after surgery by immersion in isopentane chilled with dry ice. Duplicate sections, 20 μm thick, were incubated with 5.4 nM 3H-DTBZ for 30 min in the absence or presence of 100 μM ketanserin. Sections of rat brain and bovine adrenal medulla were included as controls. The sections were washed for 20 min, dried, and exposed to imaging plates for 14 d. After autoradiographic exposure, the sections were fixed with 4% paraformaldehyde and processed for insulin immunohistochemistry analysis with a Vectastain ABC Kit (Vector Laboratories). A polyclonal guinea pig primary antibody raised against human insulin (Abcam), diluted 1:500, and then a biotinylated goat anti–guinea pig secondary antibody (Jackson ImmunoResearch), diluted 1:500, were used. The sections were counterstained with hematoxylin and photographed as described earlier. Pancreatic islets were identified and manually outlined on the micrographs, and the outlines were combined with the autoradiographic images by use of CorelDRAW X3 (Corel Corp.).
Statistics
Results were analyzed with Microsoft Excel 2002 (Microsoft Corp.) and GraphPad Prism 5 and reported as mean ± SD or as mean and 95% confidence interval. Differences between groups were assessed with a 2-tailed Student t test. Differences between ratios and 1 were analyzed with a 2-tailed 1-sample t test. A P value of less than 0.05 was considered statistically significant.
RESULTS
Biodistribution and Intrapancreatic Localization of 11C-DTBZ Radioactivity in Rats
The largest accumulation of 11C-DTBZ radioactivity was found in the pancreas, with similarly high levels at 10 and 60 min after injection (Table 1). In addition, the striatum exhibited high uptake of radioactivity 10 min after injection, but at 60 min, radioactivity in the striatum was significantly reduced; these results suggest different kinetics for 11C-DTBZ binding in the pancreas and the VMAT2-rich striatum. The amounts of nonmetabolized tracer, as determined by thin-layer chromatography 10 min after the injection of 11C-DTBZ, were 94% in plasma, 85% in urine, 83% in liver, 98% in the head of the pancreas, 99% in the middle of the pancreas, and 98% in the tail of the pancreas. Two to 3 radioactive metabolites were separated; their Rf values were lower than the Rf value of 11C-DTBZ, indicating that the metabolites were more polar than the parent compound (data not shown). The possible formation of radioactive volatile metabolites was not assessed.
11C-DTBZ Biodistribution in Rats 10 and 60 Minutes After Injection
A representative example of autoradiographic and histologic images of a pancreatic section, with pancreatic islets outlined, is shown in Figure 1. At 10 min after the injection of 11C-DTBZ, radioactivity was heterogeneously distributed; levels toward the head of the pancreas were 1.7 ± 0.6-fold (i.e., 70%) higher than those toward the tail (P = 0.046, n = 6). At 60 min after injection, this gradient was no longer seen. At either time point, 11C-DTBZ radioactivity did not accumulate specifically in pancreatic islets. In the rat brain, the expected 11C-DTBZ binding pattern, corresponding to the known distribution of VMAT2, was observed (data not shown).

Intrapancreatic distribution of radioactivity 10 min after injection of 11C-DTBZ at 1.0 nmol/kg. Same pancreatic section used for autoradiography (A) was stained with hematoxylin and eosin (B). 11C-DTBZ radioactivity did not accumulate in pancreatic islets (outlined in A and B). Radioactivity gradient with higher levels toward head of pancreas, as seen in A, was consistently observed at 10 min but not at 60 min after injection. Scale bar = 5 mm. H = head of pancreas; T = tail of pancreas.
In Vitro Binding of 11C-DTBZ in Pancreatic Sections from Diabetic and Control Rats
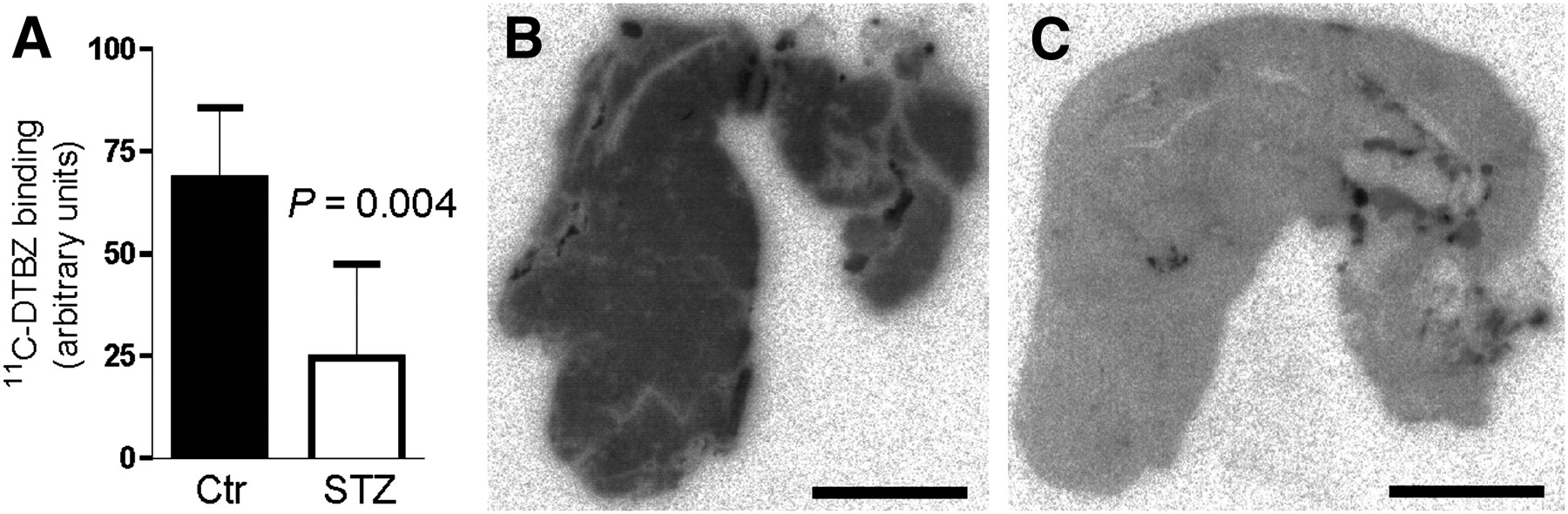
11C-DTBZ binding in the pancreas was lower in diabetic rats than in control rats (Fig. 2A), supporting previous findings obtained by in vivo PET. The decrease in 11C-DTBZ binding was observed throughout the pancreatic tissue, indicating that it was not directly associated with a decrease in β-cell mass (Figs. 2B and 2C).

(A) Pancreatic 11C-DTBZ (2.8 nM) binding was lower in sections from diabetic (STZ) rats (n = 6) than in sections from control (Ctr) rats (n = 6) (Student t test). Sections from STZ and Ctr rats were processed in parallel in same experiment. (B and C) Representative sections from control rat (B) and diabetic rat (C). Scale bar = 5 mm.
Pharmacologic Characterization of 3H-DTBZ Binding In Vitro
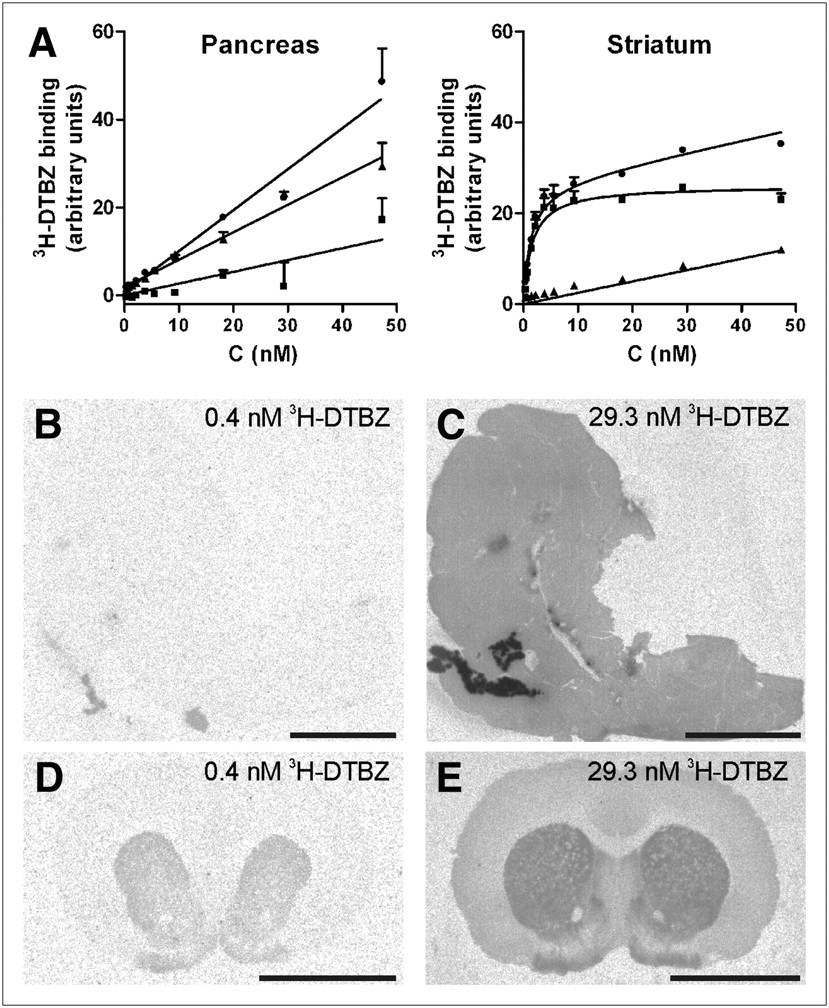
3H-DTBZ binding in sections of the rat pancreas was homogeneous, and pancreatic islets could not be visualized. In the rat pancreas, 3H-DTBZ binding increased in a concentration-dependent, nonsaturable manner (Fig. 3). In contrast, saturable 3H-DTBZ binding was observed in the rat striatum (Kd, 1.3 nM; 95% confidence interval, 1.0–1.5 nM) and bovine adrenal medulla (Kd, 3.3 nM; 95% confidence interval, 2.1–4.6 nM). In the rat striatum and bovine adrenal medulla, VMAT2 antagonists competed for 3H-DTBZ binding with the following rank order of potency: tetrabenazine > ketanserin > reserpine > haloperidol > lobeline ≈ chlorpromazine; this profile corresponded to the pharmacologic profile of VMAT2 (11,20,21). All competitors exhibited somewhat higher affinities for the rat striatum than for the bovine adrenal medulla (Table 2). A non–VMAT2 competition binding profile was observed in the rat pancreas, with the following rank order of potency: lobeline ≈ ketanserin > haloperidol ≈ chlorpromazine > tetrabenazine ≈ reserpine. The confidence intervals were wide, however, and reliable 50% inhibitory concentrations could not be determined. This situation was partly attributable to the poor displacement of pancreatic 3H-DTBZ binding by the unlabeled compounds and to the lower levels of 3H-DTBZ binding in the pancreas than in the striatum and adrenal medulla at the concentration of 3H-DTBZ used (1.8 nM). Together, the in vitro binding results suggest that 3H-DTBZ binding in the rat pancreas, in contrast to binding in the rat striatum and bovine adrenal medulla, is nonspecific.

(A) 3H-DTBZ binding curves for sections of pancreas and striatum from same rat, processed in parallel. Nonspecific binding was defined as binding in presence of 100 μM ketanserin. Specific binding (▪) was difference between total binding (•) and nonspecific binding (▲). Pancreatic 3H-DTBZ binding was nonsaturable within investigated concentration range. In striatum, 3H-DTBZ binding was saturable, with Kd of 1.3 nM. (B–E) Relative increases in pancreatic (B and C) and striatal (D and E) binding were observed with increasing concentrations of 3H-DTBZ. Scale bar = 5 mm.
Pharmacology of 3H-DTBZ Binding
3H-DTBZ Binding in Human Pancreas
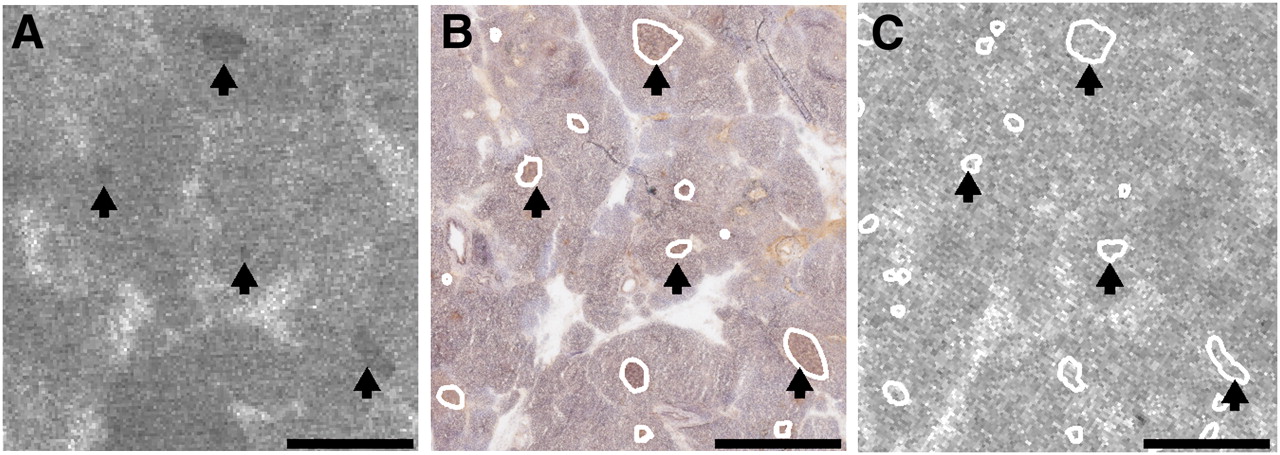
Insulin-immunoreactive pancreatic islets were observed in samples from all patients. 3H-DTBZ binding in most islets did not exceed binding in the exocrine pancreas. Some intermediate to large islets were, however, discernible against the exocrine background labeling (Fig. 4). Ketanserin at 100 μM completely inhibited islet-specific 3H-DTBZ binding, suggesting that it represented binding to VMAT2. At the 3H-DTBZ concentration used (5.4 nM), the labeling intensity in human islets was less than 2-fold the intensity in the surrounding exocrine tissue. For comparison, the ratio for the binding of the rat striatum to the rat pancreas was 3.3, that for the binding of the rat striatum to the rat cerebral cortex was 6.7, and that for the binding of the bovine adrenal medulla to the bovine adrenal cortex was 6.8. The levels of nonspecific binding were similar in rat exocrine pancreas and human exocrine pancreas.

3H-DTBZ binding in sections of human pancreas. (A) Arrows indicate subset of pancreatic islets that were labeled with 3H-DTBZ (5.4 nM). (B) Islets were identified and outlined after insulin immunohistochemistry analysis was performed on same section. (C) Adjacent section incubated with 3H-DTBZ (5.4 nM) and 100 μM ketanserin. Islets labeled with 3H-DTBZ in A were also visible in C. Absence of islet-specific binding in C indicated that labeling of islets with 3H-DTBZ in A was VMAT2-specific. Pancreatic islets in C were identified by insulin immunohistochemistry analysis (not shown) of same section. Scale bar = 1 mm.
DISCUSSION
The results of the present study demonstrated that the binding of 11C-DTBZ and 3H-DTBZ in islets of the rat pancreas did not exceed the binding in exocrine tissue. Pancreatic binding increased rapidly with increasing concentrations of 3H-DTBZ but did not represent binding to VMAT2. Nonspecific pancreatic 11C-DTBZ binding was lower in rats with streptozotocin-induced diabetes than in control rats. In the human pancreas, only weak 3H-DTBZ labeling of a small number of islets was detected.
Although β-cells, which reside in pancreatic islets scattered throughout the exocrine tissue, constitute only a small fraction of the total pancreatic volume, the highest levels of radioactivity after the injection of 11C-DTBZ into rats were found in the pancreas. Similar levels of pancreatic uptake of 3H-DTBZ and 11C-DTBZ as well as DTBZ derivatives were reported previously (13,14,22,23). In those studies, the islet and VMAT2 specificity of the accumulated radioactivity was not assessed directly. If pancreatic 11C-DTBZ uptake represented primarily binding to β-cells, then strong labeling of pancreatic islets would be expected. However, we did not observe islet-specific accumulation of radioactivity in the rat pancreas either after intravenous injection of 11C-DTBZ or under in vitro conditions with 11C-DTBZ or 3H-DTBZ; under such conditions, the concentrations of radiolabeled DTBZ could be strictly controlled, and in vivo metabolism of the tracer was not a concern. 11C-DTBZ and 3H-DTBZ displayed the expected binding patterns in the rat brain and bovine adrenal gland, demonstrating the integrity of the tracers under the binding conditions used. Transiently higher radioactivity levels toward the head than toward the tail of the rat pancreas were consistently observed 10 min after the injection of 11C-DTBZ. Because the heterogeneous distribution of radioactivity did not correspond to the distribution of pancreatic islets, the possibility of regional differences in either blood flow or blood perfusion or binding to sympathetic nerves remains. The fact that a binding gradient was not observed in pancreatic sections incubated with 11C-DTBZ or 3H-DTBZ in vitro argues against the latter alternative.
Pharmacologic comparisons of 3H-DTBZ binding in sections of the rat pancreas, rat striatum, and bovine adrenal medulla confirmed the notion that DTBZ binding in the rat pancreas did not represent binding to VMAT2. At concentrations of less than 10 nM, total 3H-DTBZ binding was considerably lower in the rat pancreas than in the rat striatum, whereas at 50 nM, the opposite was true. The amount of 11C-DTBZ administered to living rats was 2.1 ± 2.2 nmol/kg. It is conceivable that during the bolus intravenous injection, the amount of 11C-DTBZ that reached the pancreas was large enough to explain the abundant (non-VMAT2) pancreatic uptake of 11C-DTBZ observed in the biodistribution study. Nonspecific DTBZ binding in the exocrine pancreas was not readily reversible, as evident from the slow clearance of 11C-DTBZ in vivo and the poor displacement of 3H-DTBZ by VMAT2 antagonists in vitro.
Previous assumptions that the pancreatic uptake of radiolabeled DTBZ and its derivatives represented binding to VMAT2 were based on indirect evidence. In the human pancreas, VMAT2 is found in endocrine cells and sympathetic nerve fibers, as demonstrated by in situ hybridization and immunohistochemistry analysis (9). In the endocrine human pancreas, VMAT2 is mainly localized to β-cells, although a subset of cells immunoreactive for pancreatic polypeptide was also reported to express VMAT2 (24). Mei et al. (25) reported the loss of VMAT2-immunoreactive sympathetic fibers in pancreatic islets at the onset of autoimmune but not streptozotocin-induced type 1 diabetes in rats. In that report, VMAT2 immunostaining was not evident in islets. In contrast, Souza et al. (13) and Simpson et al. (14) described decreases in islet VMAT2 and insulin immunoreactivity in association with reduced pancreatic VMAT2 and insulin messenger RNA expression in both rats with streptozotocin-induced diabetes and rats with autoimmune type 1 diabetes. These reductions were correlated with reduced pancreatic uptake of 11C-DTBZ, as determined by PET. We found similar reductions in 11C-DTBZ binding in pancreatic sections from rats with streptozotocin-induced diabetes, but because the reduction in binding occurred uniformly throughout the pancreatic tissue, it could not be attributed to a decrease in β-cell mass. Supporting evidence was provided by a recent report in which sleeve gastrectomy and duodenal–jejunal bypass surgery in Goto–Kakizaki type 2 diabetic rats increased pancreatic 11C-DTBZ uptake and circulating insulin levels in association with a decrease, rather than an increase, in β-cell mass (26). The mechanism for the reduction of non-VMAT2 DTBZ binding in the exocrine pancreas in rats with streptozotocin-induced or autoimmune type 1 diabetes is still unknown. In rats with autoimmune diabetes, the reduction in pancreatic 11C-DTBZ uptake preceded the onset of diabetes (13), suggesting that hyperglycemia may not be necessary for the downregulation of DTBZ binding sites.
Although some islets were weakly labeled with 3H-DTBZ in sections of the human pancreas, 3H-DTBZ binding in the great majority of islets did not exceed binding in the exocrine pancreas. Similar levels of nonspecific 3H-DTBZ binding were observed in rat exocrine pancreas and human exocrine pancreas. This finding is in agreement with the residual pancreatic uptake of 11C-DTBZ detected by PET in patients with long-standing type 1 diabetes (15), expected to be devoid of β-cells. Together, these results imply that the pancreatic uptake of 11C-DTBZ in humans also represents mainly nonspecific binding. Extrapolating from our results for rats, we suggest that this nonspecific binding may be reduced in type 1 diabetes, resulting in the misinterpretation of PET results. Recently, radiolabeled derivatives of DTBZ with a higher affinity for VMAT2, including 18F-9-fluoropropyl-(+)-DTBZ and 18F-(+)-2-oxiranyl-3-isobutyl-9-(3-fluoropropoxy)-10-methoxy-2,3,4,6,7,11b-hexahydro-1H-pyrido[2,1-a]isoquinoline, were developed as alternative VMAT2 PET tracers. A higher affinity for VMAT2 could improve the labeling of pancreatic islets. However, because 11C-DTBZ and its derivatives displayed similarly high levels of pancreatic uptake when injected into rats (22,23), the derivatives are likely to exhibit the same type of nonspecific pancreatic binding as the parent compound.
CONCLUSION
The results of the present study suggested that 11C-DTBZ cannot be used for the quantitative imaging of pancreatic β-cell mass. Further studies are needed to determine whether the observed changes in nonspecific pancreatic 11C-DTBZ binding in association with type 1 diabetes and, possibly, with other pathologic or physiologic conditions will provide some biologically or clinically relevant information. In addition, the possibility that VMAT2 tracers other than 11C-DTBZ will be better suited for the imaging of pancreatic β-cells remains.
Acknowledgments
The technical assistance of Tarja Marttila, Marko Vehmanen, and Elisa Riuttala is acknowledged. The research leading to these results has received funding from the European Community's Seventh Framework Program (FP7/2007–2013) under grant agreement 222980 (“Use of Innovative Strategies for β-Cell Imaging in Diabetes Mellitus”) and from the Diabetes Research Foundation, Tampere, Finland. This study was conducted within the Finnish Centre of Excellence in Molecular Imaging in Cardiovascular and Metabolic Research, which is financially supported by the Academy of Finland, University of Turku, Turku University Hospital, and Åbo Akademi University.
- © 2010 by Society of Nuclear Medicine
REFERENCES
- Received for publication December 31, 2009.
- Accepted for publication March 24, 2010.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Positron Emission Tomography Ligand [11C]5-Hydroxy-Tryptophan Can Be Used as a Surrogate Marker for the Human Endocrine Pancreas
- In Vivo Imaging of Endogenous Pancreatic {beta}-Cell Mass in Healthy and Type 1 Diabetic Subjects Using 18F-Fluoropropyl-Dihydrotetrabenazine and PET
- Multimodal image coregistration and inducible selective cell ablation to evaluate imaging ligands
- 18F-Fallypride PET of Pancreatic Islets: In Vitro and In Vivo Rodent Studies
- 11C-Dihydrotetrabenazine {beta}-Cell Imaging