


Abstract
The herpes simplex virus type 1 thymidine kinase (HSV1-tk)/green fluorescent protein (TKGFP) dual-reporter gene and a multimodality imaging approach play a critical role in monitoring therapeutic gene expression, immune cell trafficking, and protein–protein interactions in translational molecular–genetic imaging. However, the cytotoxicity and low temporal resolution of TKGFP limits its application in studies that require a rapid turnover of the reporter. The purpose of this study was to construct a novel mutant TKGFP fusion reporter gene with low cytotoxicity and high temporal resolution for use in the real-time monitoring of temporal dynamics and spatial heterogeneity of hypoxia-inducible factor 1 (HIF-1) signal transduction activity mediated by hypoxia and reoxygenation in vitro and in vivo. Methods: Destabilized TKGFP was produced by inserting the nuclear export signal (NES) sequence at the N terminus and fusing the degradation domain of mouse ornithine decarboxylase (dMODC) at the C terminus. The stability of TKGFP in living NG4TL4 cells was determined by Western blot analysis, HSV1-tk enzyme activity assay, and flow cytometric analysis. The suitability of NESTKGFP:dMODC as a transcription reporter was investigated by linking it to a promoter consisting of 8 copies of hypoxia-responsive elements, whose activities depend on HIF-1. The dynamic transcriptional events mediated by hypoxia and reoxygenation were monitored by NESTKGFP:dMODC or TKGFP and determined by optical imaging and PET. Results: Unlike TKGFP, NESTKGFP:dMODC was unstable in the presence of cycloheximide and showed a short half-life of protein and enzyme activity. Rapid turnover of NESTKGFP:dMODC occurred in a 26S proteasome–dependent manner. Furthermore, NESTKGFP:dMODC showed an upregulated expression and low cytotoxicity in living cells. Studies of hypoxia-responsive TKGFP and NESTKGFP:dMODC expression showed that NESTKGFP:dMODC as a reporter gene had better temporal resolution than did TKGFP for monitoring the dynamic transcriptional events mediated by hypoxia and reoxygenation; the TKGFP expression level was not optimal for the purpose of monitoring. Conclusion: In translational molecular–genetic imaging, NESTKGFP:dMODC as a reporter gene, together with optical imaging and PET, allows the direct monitoring of transcription induction and easy determination of its association with other biochemical changes.
- herpes simplex virus type 1 thymidine kinase/green fluorescent protein (TKGFP)
- mouse ornithine decarboxylase (MODC)
- hypoxia
- hypoxia-inducible factor 1 (HIF-1)
- molecular–genetic imaging
In translational molecular–genetic imaging, the multimodality imaging approach combined with reporter genes plays a critical role in detecting endogenous signal transduction activity or transgene expression for molecular therapies or in monitoring in vivo cell trafficking for adaptive cell therapies (1). Combining 2 different technologies via the construction of a fusion gene has the advantage of speedier and easier validation of approaches in cells or small animals and, in turn, in humans (2,3). Furthermore, the combination approach can also overcome the shortcomings of individual imaging modalities and promote the application of translational research in molecular medicine (4,5). Therefore, the herpes simplex virus type 1 thymidine kinase (HSV1-tk)/green fluorescent protein (TKGFP) dual-reporter gene was constructed and used in various research studies such as noninvasive in vitro and in vivo monitoring of therapeutic gene expression, endogenous signal transduction activities, immune cell trafficking, and protein–protein interactions (6–10). The use of this reporter gene imaging technology in clinical applications is critical for various preclinical studies on disease therapy.
Although TKGFP is widely used as a reporter in molecular–genetic imaging for noninvasive and repetitive imaging of endogenous biologic processes and in vivo cell trafficking together with optical and nuclear medicine imaging (3), there are several potential problems associated with its applications in translational research. First, the potential cytotoxicity triggered by the nuclear tropism of HSV1-tk may limit its applications in clinical practice. In addition, the TKGFP fusion protein seems to be highly stable in mammalian cells and, therefore, has lower temporal resolution than that required for monitoring the temporal dynamics of endogenous signal transduction activity or transgene expression. The GFP is not cytotoxic to mammalian cells or animals (11,12); however, the HSV1-tk enzyme has been shown to be cytotoxic when used as a selection marker or a therapeutic gene in transgenic mice or in gene therapy strategies because of the nuclear tropism of its protein and the presence of a putative cryptic testis-specific promoter (13,14). It has not been determined whether the TKGFP fusion protein is cytotoxic, but it seems to exist because of the presence of the DNA sequences of HSV1-tk in the N-terminal range of TKGFP.
It has been shown that both GFP and HSV1-tk are highly stable in mammalian cells (15,16). This high stability limits the application of GFP and HSV1-tk in monitoring short-time-scale dynamic processes such as kinetic gene expression controlled by inducible promoters or by a less stable protein with a more rapid turnover. The reason is the long-half-life protein or enzyme activity of the reporter that interferes with the subsequent protein or enzyme activity, respectively, when the inducible promoter is turned on or off by the triggered events. Although the half-life of the TKGFP has not been elucidated in detail, it can be expected that TKGFP is highly stable in mammalian cells because of the stability of GFP and HSV1-tk. Therefore, TKGFP as a reporter may have low temporal resolution and may be unsuitable for monitoring short-time-scale dynamic processes. For these reasons, it is necessary to generate a novel mutant form of TKGFP with low cytotoxicity and high temporal resolution for use in translational molecular–genetic imaging.
The hypoxia-inducible factor 1 (HIF-1) plays a central role in eliciting transcriptional responses to changes in oxygen availability (17,18). More than a 100 genes experience changes in their expression, directly or indirectly regulated by HIF (19,20). HIF-mediated pathways influence metabolic adaptation, erythropoiesis, angiogenesis and vascular tone, cell growth and differentiation, and survival and apoptosis and are, thus, critical factors involved in development, physiology, and pathology (21–24). In cancer, HIF-1 functions not only as an important transcriptional mediator of tumor response to hypoxia but also as an attractive therapeutic target (25). HIF-1 activates the transcription of genes that are involved in the crucial aspects of cancer biology, including angiogenesis, cell survival, glucose metabolism, tumor invasion, and metastasis (26–28). HIF-1 signal transduction activity in a tumor is a spatially and temporally heterogeneous phenomenon, which occurs as a result of the effects of tumor microenvironment and genetic alteration (29,30). Furthermore, the dynamics of HIF-1 signal transduction activity mediated by cyclic hypoxia in a tumor are fast because of the instability of the HIF-1α protein under the reoxygenation condition; therefore, a reporter gene with a high temporal resolution is required for monitoring such dynamic processes. Although TKGFP has been used for monitoring the temporal dynamics and spatial heterogeneity of HIF-1 signal transduction activity in tumors in living subjects (31), because of its poor temporal resolution its use seems difficult for the real-time monitoring of the dynamics of HIF-1 signal transduction activity mediated by hypoxia and reoxygenation in tumors.
The purpose of the present study was to construct a novel mutant TKGFP fusion reporter gene with low cytotoxicity and high temporal resolution for use in translational molecular–genetic imaging. This novel mutant TKGFP was used for monitoring temporal dynamics and spatial heterogeneity of HIF-1 signal transduction activity mediated by hypoxia and reoxygenation in tumors in living mice.
MATERIALS AND METHODS
Plasmid Construct
The HSV1-tk was digested with BglII and XmaI from the pCMV-HSV1-TK vector described elsewhere (16). This fragment was inserted into the pEGFP-N1 and pd2EGFP vectors, which were digested with BglII and XmaI. These constructed vectors were termed pCMV-TKGFP and pCMV-TKGFP:dMODC, respectively. The HSV1-tk gene containing the leucine-rich nuclear export signal (NES) sequence of the mitogen-activated protein kinase cDNA of Xenopus encoding 20 amino acids was amplified from the plasmid SFG-NES-TKGFP described elsewhere (32), using polymerase chain reaction (PCR). The primers used, (5′-GAAGATCTATGGCCTCGTACCCCGGCCA-3′) and (5′-AATTCCCCGGGTAGTTGTACTCCAGCTTGTG-3′), contain the restriction enzyme sites of BglII and XmaI. The PCR product was subcloned into the pEGFP-N1 and pd2EGFP vectors, which were digested with BglII and XmaI. The resulting vectors were termed pCMV-NESTKGFP and pCMV-NESTKGFP:dMODC, respectively. For the monitoring of HIF-1 signal transduction activity, the cDNA fragment bearing 8 repeats of the hypoxia-responsive elements from the 3′ enhancer region of the human Epo gene was amplified using PCR from the plasmid dxHRE-tk/eGFP-cmvRed2XPRT described elsewhere (31). The primers used, (5′-CGCCAATTAATAATTGGCCCTACGTGCTGTC-3′) and (5′-TAATAGCTAGCGAGCTTCCATTATATACCCT-3′), contain the restriction enzyme sites of AseI and NheI for insertion into the pCMV-TKGFP and pCMV-NESTKGFP:dMODC vectors, from which was deleted the human cytomegalovirus (CMV) immediate early promoter using AseI and NheI restriction enzymes. These constructed vectors were termed p8xHRE-TKGFP and p8xHRE- NESTKGFP:dMODC, respectively.
Determination of Protein Localization of Mutant TKGFP in Cells
The constructed DNAs—pCMV-TKGFP, pCMV-NESTKGFP, pCMV-TKGFP:dMODC, and pCMV-NESTKGFP:dMODC—were introduced into the NG4TL4 cells using jetPEI (Polyplus-Transfection). After 48 h, the subcellular localization of the TKGFP, NESTKGFP, TKGFP:dMODC, or NESTKGFP:dMODC protein in transduced tumor cells was visualized by fluorescence microscopy (Nikon) using a 488-nm excitation beam and 510-nm emission filters.
Determination of Protein Stability of Mutant TKGFP in Living Cells
The pCMV-TKGFP, pCMV-NESTKGFP, pCMV-TKGFP:dMODC, and pCMV-NESTKGFP:dMODC plasmids were transfected into the NG4TL4 cells using jetPEI. At 24 h after the transfection, the culture medium was changed to the medium containing 600 μg of G418 disulfate per milliliter, and the cells were cultured for an additional 2 wk. The stable transfectants were isolated by fluorescence-activated cell sorting (FACS). The stably transfected cells were treated with cycloheximide (50 μg/mL; Sigma) dissolved in minimal essential medium at 0, 1, 2, 3, 4, and 5 h. At each time point, cells were trypsinized, counted by hemocytometry, and concentrated by centrifugation. The supernatant was removed and then dissolved in 40 μL of phosphate-buffered saline and stored at −80°C for Western blot analysis and HSV1-tk enzyme activity assay. Conversely, the cells were fixed in 2% paraformaldehyde after cell harvest and stored at 4°C for flow cytometry analysis.
Western Blot Analysis
The procedures for Western blot analysis are described in detail elsewhere (16). The protein expression of TKGFP, HIF-1α, and CXCR4 was detected with monoclonal anti-HSV1-tk antibody, monoclonal anti-HIF-1α antibody, and polyclonal anti-CXCR4 antibody, respectively.
HSV1-tk Enzyme Activity Assay
The HSV1-tk enzyme activity assay was based on the protocol described elsewhere (33). Briefly, the stored cells were lysed in a lysis buffer, and then the cell extracts were incubated for 60 min with 8-3H-ganciclovir to determine the formation of phosphorylated 8-3H-ganciclovir. The extracts were then separated from phosphorylated 8-3H-ganciclovir using a DE-81 filter (Whatman). These DE-81 filter papers were digested by cellulase and counted in a scintillation counter (LS 6500; Beckman).
Flow Cytometry Analysis
Green fluorescence was analyzed with a flow cytometer (FACSCanto; BD Biosciences), and the data were processed with FACSDvia software (BD Biosciences). Histograms of GFP were plotted with the log scale for green fluorescence, and the results were recorded as means of the middle of the peaks.
Proteasome Inhibition Assay
Two vectors, pCMV-TKGFP and pCMV-NESTKGFP:dMODC, were transfected into NG4TL4 cells using jetPEI. After 24 h, cells were treated with cycloheximide (50 μg/mL; Sigma) with or without 40 μM MG-132. After an additional 6 h of incubation, cells were trypsinized and counted by hemocytometer and concentrated by centrifugation. The supernatant was removed and then dissolved in 40 μL of phosphate-buffered saline and stored at −80°C for the Western blot analysis.
In Vitro Cell Proliferation Assay
Wild-type NG4TL4, NG4TL4-TKGFP, and NG4TL4-NESTKGFP:dMODC cells were seeded into a 96-well plate (Falcon; Becton Dickinson Labware) at 5 × 105 cells per well in minimum essential medium containing 10% fetal calf serum. The number of cells was quantified using trypan blue staining.
Hypoxia Induction and Reoxygenation
The NG4TL4 cells were seeded on a 60-mm culture dish and transfected with 5 μg of p8xHRE-TKGFP and p8xHRE-NESTKGFP:dMODC using jetPEI. At 24 h after the transfection, the culture medium was changed to the medium containing 600 μg of G418 disulfate per milliliter, and the cells were cultured for an additional 2 wk. The stable transfectants were treated under hypoxic conditions (0.02%) for 20 h before being isolated. The cell population expressing TKGFP or NESTKGFP:dMODC at high levels was selected using FACS. To assay the hypoxia-responsive TKGFP and NESTKGFP:dMODC expression in living NG4TL4 cells, the reporter cells were treated in a Biospherix C-Chamber (model C-274; Biospherix) inside a standard culture chamber by means of exhausting and gassing with 95% N2 and 5% CO2 for various cycles, to produce oxygen concentrations of 20%, 2%, 0.2%, 0.02%, and 0.01% for 24 h. The HSV1-tk enzyme activity and fluorescent intensity from these reporter cells under various hypoxia treatments were measured by HSV1-tk enzyme activity assay and flow cytometry analysis, respectively. For the reoxygenation study, the reporter cells were cultured in the incubator with 95% air and 5% CO2 for the times indicated in figures 1–6⇓⇓⇓⇓⇓ to reoxygenate the cells after the hypoxia treatment (0.02% O2) for 24 h. The dynamics of HIF-1 signal transduction activity mediating hypoxia and reoxygenation in the reporter cells were also determined by HSV1-tk enzyme activity assay and flow cytometry analysis.
Animal Studies
To control the same tumor size for imaging, the different number of reporter tumor cells was used for tumor inoculation because of the consideration of the effect from the difference of tumor growth rate. The NG4TL4-TKGFP (2.0 × 105) and NG4TL4-NESTKGFP:dMODC reporter cells (1.2 × 105) were injected subcutaneously into the dorsal aspects of both anterior limbs of nude mice, and small (30 ± 5.0 mm3) subcutaneous tumors developed 12 d later. At that point, 6 mice bearing NG4TL4-TKGFP and NG4TL4-NESTKGFP:dMODC reporter tumors were imaged to determine baseline levels of expression of the TKGFP and NESTKGFP:dMODC reporters using PET and in vivo optical imaging. To induce the acute hypoxia in vivo, a tourniquet was applied for 1 h proximally to the tumor on both anterior limbs after animal imaging. PET and in vivo optical imaging were performed at 24 and 48 h after the first animal imaging. Furthermore, another 3 mice from each group bearing NG4TL4-TKGFP and NG4TL4-NESTKGFP:dMODC reporter tumors were sacrificed with chloroform. Tumors were removed and assayed for HIF-1α expression by immunohistochemistry. The animal experiments were approved by the Laboratory Animal Care Panel of the National Yang-Ming University and China Medical University.
Immunohistochemistry
The tumor tissues were fixed in 4% paraformaldehyde and embedded in optimal-cutting-temperature compound. Five-micrometer sections were immunostained with mouse monoclonal anti-HIF-1α (Novus Biologicals), visualized with an AEC kit (InnoGenex), and counterstained with hematoxylin.
Animal Imaging
Animals were injected with 9.25 × 106 Bq of 18F-9-(4-fluoro-3-hydroxymethyl-butyl)guanine (FHBG) and imaged on a small-animal PET scanner (microPET; Concorde Microsystems). Images were acquired for 15 min under inhalation anesthesia (isoflurane 2%). The level of radioactivity in tumors (percentage dose per gram) was estimated from images according to the method published by Tjuvajev et al. (34). Furthermore, in vivo GFP expression was determined using the IVIS Imaging System 200 Series (Xenogen Corp.) under a 488-nm excitation light. Living Image software (Xenogen Corp.) was used for quantification analysis, and data were analyzed using photons/s/cm2/steradian in a region of interest. All of the animals were imagined under inhalation anesthesia (isoflurane, 2%).
Statistical Analysis
All values are shown as mean ± SD. Statistical analyses were performed using an unpaired Student t test to evaluate the significance of differences in values between 2 treated groups. A 2-tailed value of P less than 0.05 was considered significant.
RESULTS
Subcellular Localization of Different Mutant TKGFP Fusion Proteins
Fluorescent microscopic analysis of transfected NG4TL4 cells revealed distinct differences in the subcellular localization of mutant TKGFP fusion proteins (Fig. 1). TKGFP and TKGFP:dMODC localized predominantly in the cell nucleus because of the function of the nuclear location signal present in HSV1-tk. NESTKGFP and NESTKGFP:dMODC were predominantly distributed in the cytoplasm, and the cell nuclei were dark. These results indicated that when the NES sequence from mitogen-activated protein kinase was added at the N terminus, the original localization pattern of TKGFP changed and the function of NES triggered protein export from the cell nucleus to cytoplasm.
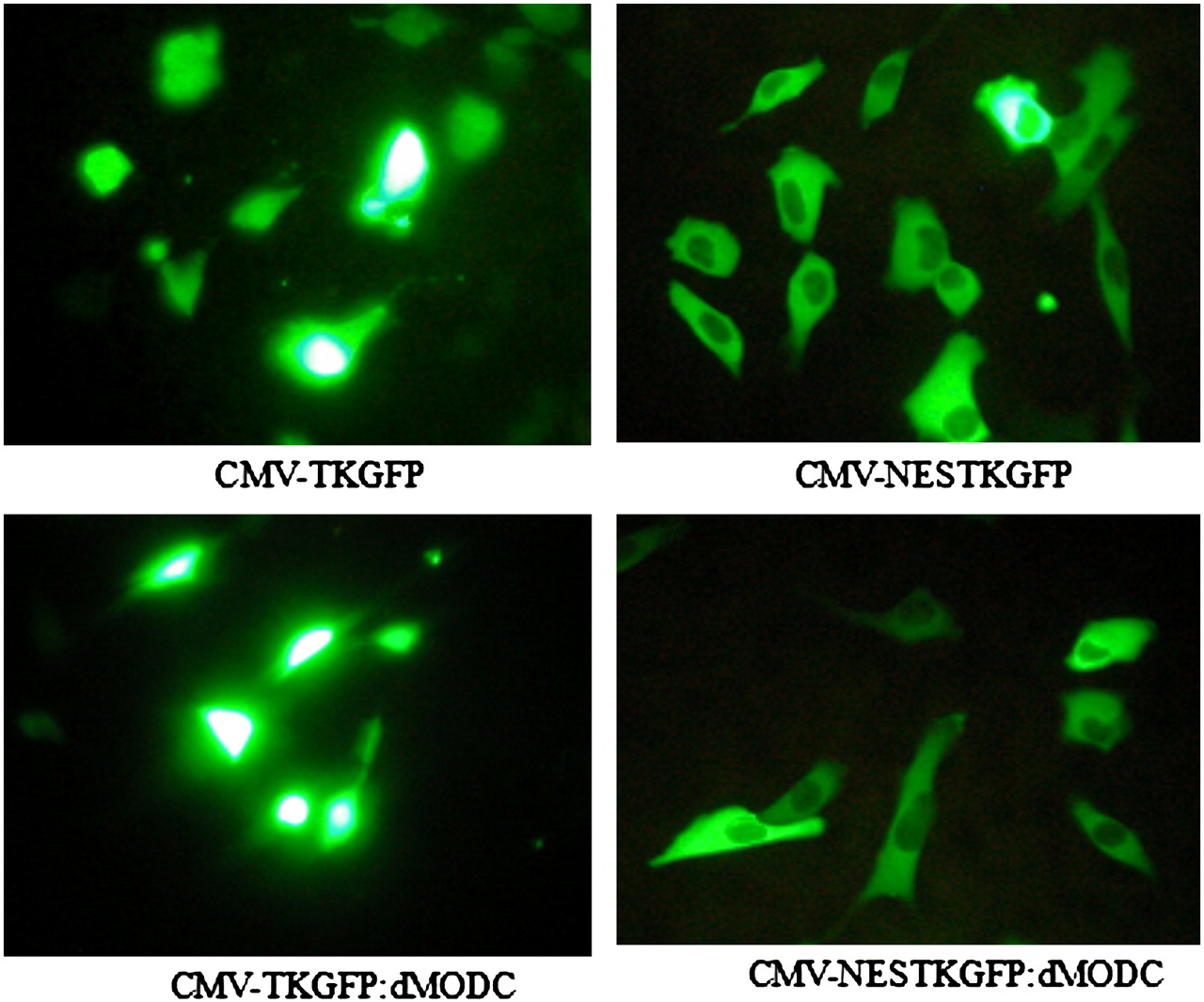
Fluorescent photomicrographs of NG4TL4 cells expressing CMV-TKGFP, CMV-NESTKGFP, CMV-TKGFP:dMODC, and CMV-NESTKGFP:dMODC.
Protein and Enzyme Stability of Different Mutant TKGFP Fusion Proteins
No noticeable change was observed in the TKGFP level between the TKGFP- and NESTKGFP-transfected cells at 0 h after the cycloheximide treatment (Fig. 2). However, NESTKGFP:dMODC was unstable, as determined by a measured decline after the cycloheximide treatment. This unstable condition of the protein was also observed, although to a small extent, in the TKGFP:dMODC-transfected cells. About 50% NESTKGFP:dMODC was degraded within 3 h of the cycloheximide treatment. In contrast to NESTKGFP:dMODC, TKGFP and NESTKGFP were stable during the observation period. Furthermore, HSV1-tk enzyme activity assay and flow cytometric analysis revealed similar results for the enzyme activity and fluorescence intensity, respectively, of TKGFP, NESTKGFP, and NESTKGFP:dMODC in the NG4TL4 cells treated with cycloheximide (Figs. 3A and 3B). The half-lives of TKGFP and NESTKGFP:dMODC in the living cells calculated by fitting an exponential function were about 26 and 2.5 h, respectively.

Western blot analysis of TKGFP, NESTKGFP, TKGFP:dMODC, and NESTKGFP:dMODC stability in living NG4TL4 cells, with or without treatment with 0.1 mg of cycloheximide per milliliter.
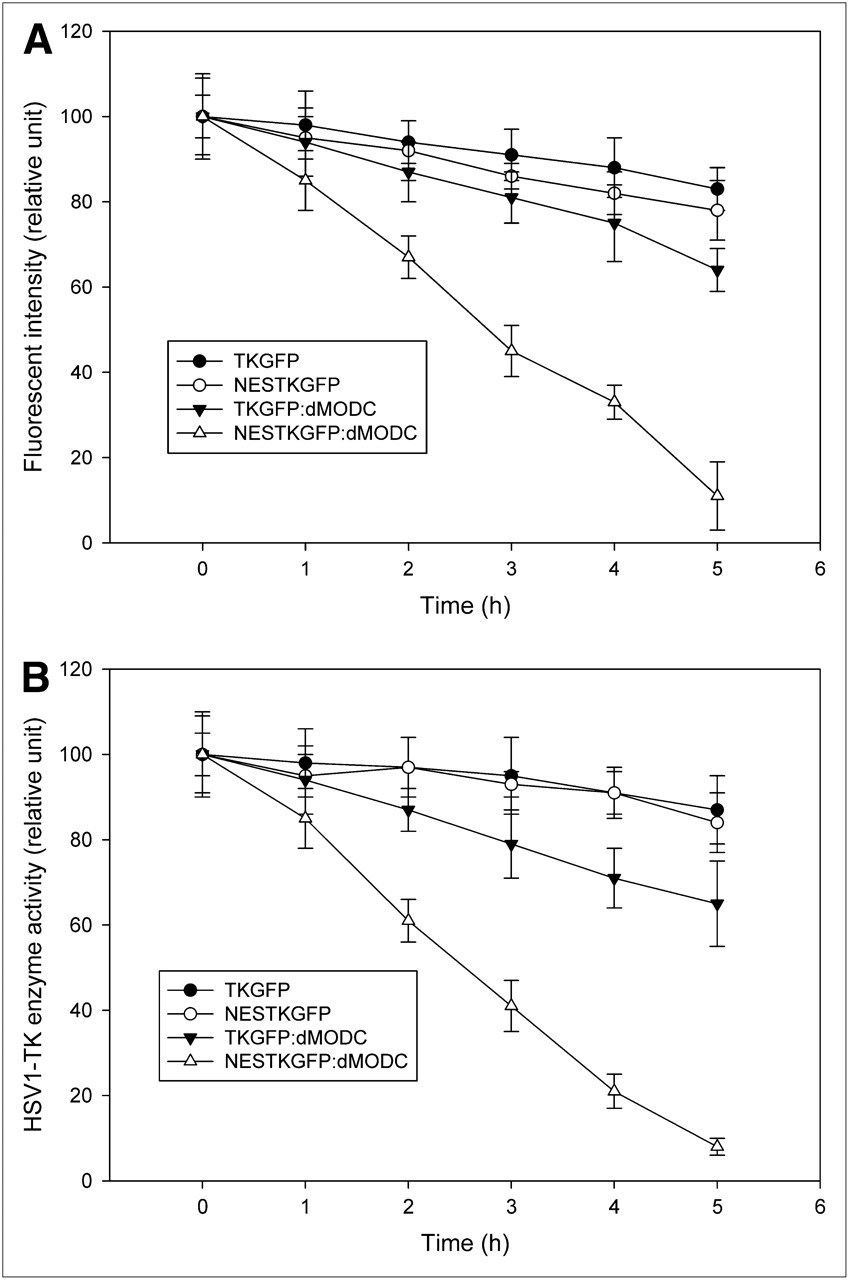
Stability of TKGFP, NESTKGFP, TKGFP:dMODC, and NESTKGFP:dMODC in living NG4TL4 cells, with or without treatment with 0.1 mg of cycloheximide per milliliter. (A) Fluorescent intensity was determined by FACS. (B) HSV1-tk activity in lysed cells was determined by in vitro HSV1-tk enzyme activity assay. Data represent mean ± SD of values obtained from 3 independent experiments.
Degradation of Different Mutant TKGFP Fusion Proteins
Proteasome inhibition assay revealed no significant decrease in the protein expression level in the TKGFP- or NESTKGFP-transfected NG4TL4 cells, with or without the cycloheximide treatment (Supplemental Fig. 1; supplemental materials are available online only at http://jnm.snmjournals.org). In contrast, a significant decrease in the protein expression level was observed in the NESTKGFP:dMODC-transfected NG4TL4 cells after the cycloheximide treatment, and this decrease could be inhibited using the proteasome inhibitor MG-132. This observation indicated that the rapid turnover of NESTKGFP:dMODC occurred in a 26S proteasome–dependent manner.
Toxicity of TKGFP and NESTKGFP:dMODC in Transfected Cells
The comparison of cell growth curves indicated no significant difference in the growth rate between the NG4TL4 control cells and the NESTKGFP:dMODC-transfected NG4TL4 cells (Supplemental Fig. 2). However, the growth of the TKGFP-transfected NG4TL4 cells, compared with that of the NG4TL4 control cells, was inhibited. These findings indicated that overexpression of the NESTKGFP:dMODC reporter gene did not induce any significant cytotoxicity in the mammalian cells.
HIF-1 Transcription Activity Under Different Hypoxic Conditions
Hypoxia-responsive TKGFP and NESTKGFP:dMODC expression in living NG4TL4 cells is shown in Supplemental Figure 3. Both TKGFP and NESTKGFP:dMODC represented the HIF-1 transcription activity in the NG4TL4 cells under 0.1% oxygen during 24 h of incubation. To confirm whether NESTKGFP:dMODC can represent the HIF-1 transcription activity mediated by different hypoxic conditions, the HIF-1 signal transduction activity was measured after exposure of the NESTKGFP:dMODC reporter cells to reduced oxygen concentrations. HSV1-tk enzyme activity assay and flow cytometric analysis revealed that the HSV1-tk enzyme activity and fluorescence intensity, respectively, of TKGFP and NESTKGFP:dMODC changed in a dose-dependent manner under different oxygen concentrations (Figs. 4A and 4B). Reduced oxygen levels also induced an inverse concentration-dependent activation of HIF-1 transcription and upregulation of TKGPF or NESTKGFP:dMODC expression in the NG4TL4 reporter cells. The threshold value of oxygen concentration for TKGPF or NESTKGFP:dMODC reporter gene induction was approximately 0.2% atmospheric oxygen. In brief, these data demonstrated that both TKGFP and NESTKGFP:dMODC reporter genes represented the HIF-1 transcription activity under different hypoxic conditions.
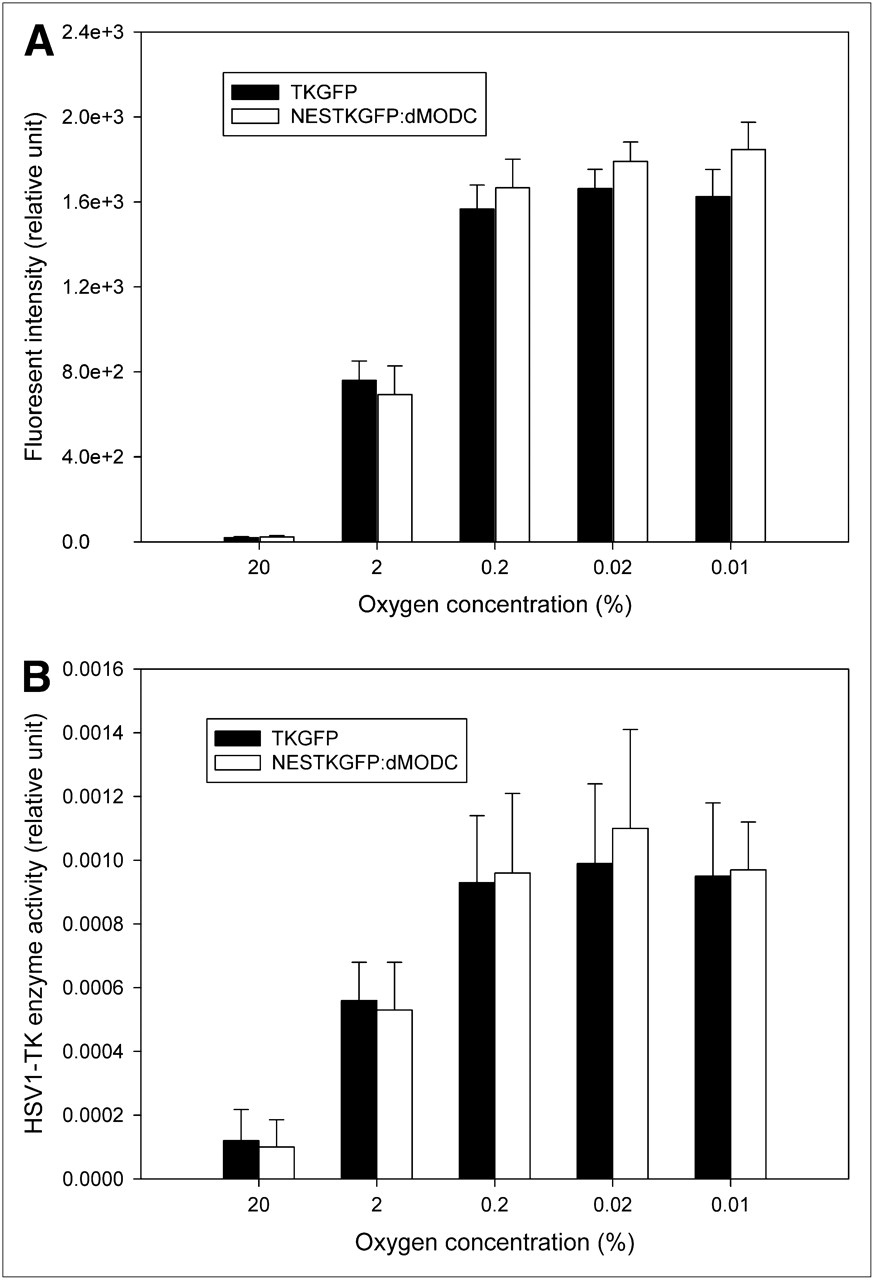
Dose-dependent TKGFP and NESTKGFP:dMODC expression in reporter cells under different hypoxic conditions. FACS (A) and HSV1-tk (B) enzyme activity assay of reporter cells exposed to various oxygen concentrations for 24 h. Bars represent mean ± SD of values obtained from 3 independent experiments.
Hypoxia- and Reoxygenation-Mediated Dynamics of HIF-1 Transcription Activity In Vitro
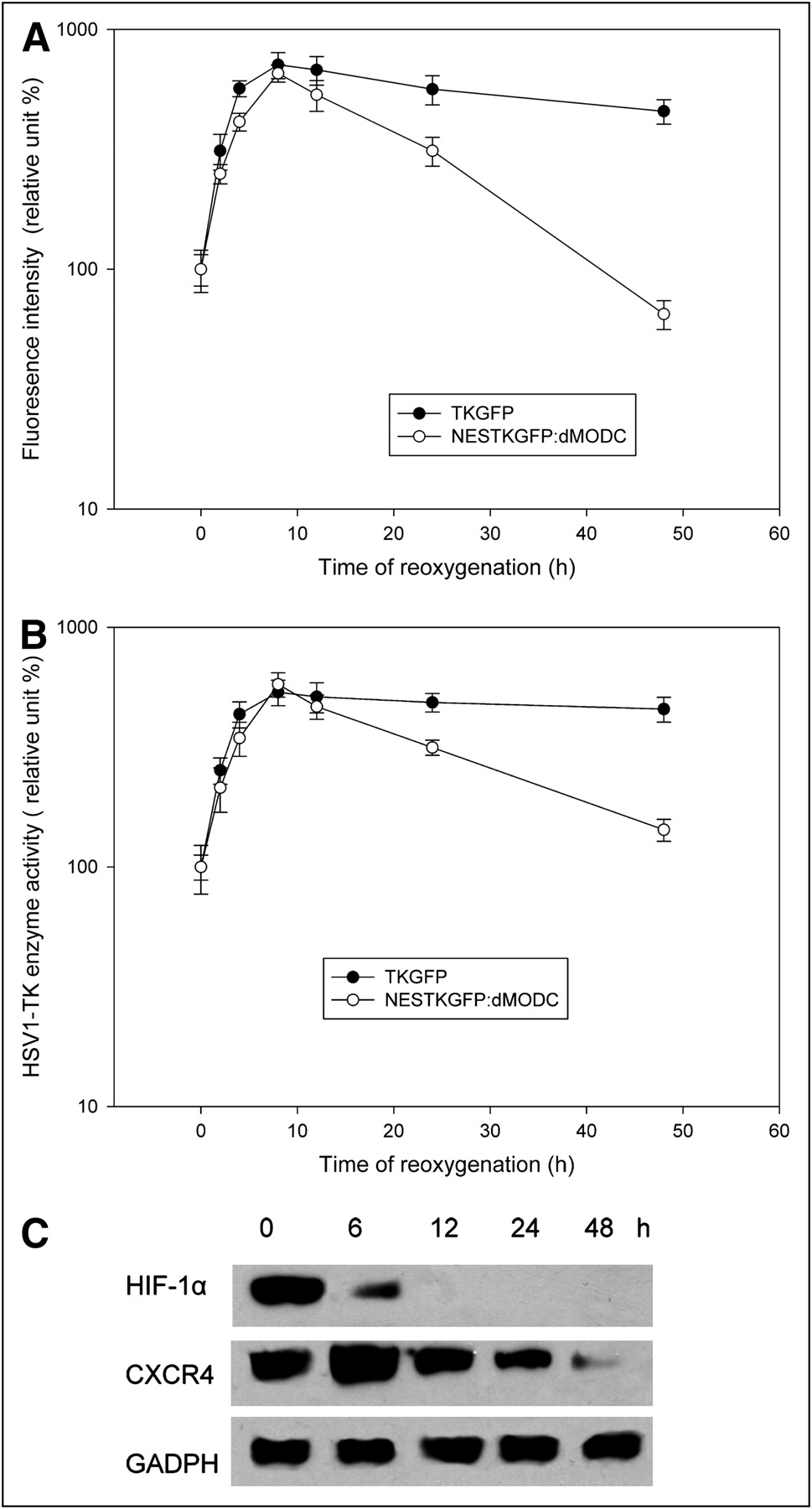
To compare the temporal resolution of TKGFP and NESTKGFP:dMODC, TKGFP and NESTKGFP:dMODC reporter cells, respectively, were subjected to hypoxic conditions (0.1% O2) for 20 h and then reoxygenated in an incubator with 95% air and 5% CO2 for different times. The hypoxia treatment without reoxygenation, as compared with that observed under aerobic conditions, enhanced HSV1-tk enzyme activity and fluorescence intensity by a factor of greater than 100 in the TKGFP and NESTKGFP:dMODC reporter cells (Figs. 5A and 5B). After 8 h of reoxygenation, sharp and high peaks of HSV1-tk enzyme activity and fluorescence intensity appeared in the TKGFP and NESTKGFP:dMODC reporter cells, after which the HSV1-tk enzyme activity and fluorescence intensity decreased significantly in the NESTKGFP:dMODC reporter cells but not in the TKGFP reporter cells. The similar effect also appeared in the protein expression of CXCR4 (an HIF-1 target gene) (Fig. 5C). Furthermore, the HIF-1α Western blot analysis also confirmed that HIF-1α was unstable under the reoxygenation condition. These data indicated that NESTKGFP:dMODC as a reporter has better temporal resolution than does TKGFP and is able to clearly demonstrate the dynamics of HIF-1 transcription activity mediated by hypoxia and reoxygenation.

Hypoxia- and reoxygenation-mediated dynamics of HIF-1 transcription activity in vitro. FACS (A), HSV1-tk enzyme activity assay (B), and Western blot analysis (C) of HIF-1α and CXCR4. TKGFP and NESTKGFP:dMODC reporter cells were subjected to hypoxia treatment (0.1% O2) for 20 h and then reoxygenated in incubator with 95% air and 5% CO2 for different times. Hypoxia-responsive TKGFP and NESTKGFP:dMODC expression in living NG4TL4 cells was determined by FACS and HSV1-tk enzyme activity assay. Protein expression of HIF-1α and its target gene, CXCR4, during hypoxia and reoxygenation was measured by Western blot analysis. GADPH = glyceraldehyde-3-phosphate dehydrogenase.
Hypoxia- and Reoxygenation-Mediated Dynamics of HIF-1 Transcription Activity In Vivo
To observe the HIF-1 transcription activity in a solid tumor in living mice, the first animal imaging study was performed to observe the baseline TKGFP and NESTKGFP:dMODC expression levels. 18F-FHBG accumulation and fluorescence intensity were low and similar in both the TKGFP and NESTKGFP:dMODC reporter tumors (P > 0.47; Fig. 6; Table 1). Furthermore, HIF-1α immunohistochemistry confirmed no detectable expression of HIF-1α in both the TKGFP and NESTKGFP:dMODC reporter tumors (Supplemental Fig. 4). The results from the second animal imaging study revealed a substantially high 18F-FHBG accumulation and fluorescence intensity in both the TKGFP and the NESTKGFP:dMODC reporter tumors after hypoxia treatment. High HIF-1α expression was also observed in the reporter tumors. The third animal imaging study demonstrated that 18F-FHBG accumulation and fluorescence intensity in the NESTKGFP:dMODC reporter tumor with reoxgenation had decreased significantly (P < 0.0001). However, no significant changes in 18F-FHBG accumulation and fluorescence intensity in the TKGFP reporter tumor with reoxgenation were observed (P > 0.38). In addition, immunohistochemistry studies verified the degradation of the HIF-1α induced in the reporter tumors with reoxygenation. These results showed a rapid decrease in the HIF-1 transcriptional activity mediated by acute hypoxia in the tumor tissue; further, they indicated that NESTKGFP:dMODC as a reporter has better temporal resolution than does TKGFP for monitoring the dynamics of HIF-1 transcription activity.

In vivo small-animal PET and optical imaging of HIF-1 transcription activity mediated by hypoxia and reoxygenation. Axial PET images (upper panel) and in vivo optical images (lower panel) of HIF-1–mediated TKGFP or NESTKGFP:dMODC expression in TKGFP (right side) or NESTKGFP:dMODC (left side) reporter tumor in forelimbs of mouse under aerobic, hypoxic, and reoxygenation conditions. In this study, 6 mice underwent PET and in vivo optical imaging.
Quantitative Data Obtained from Sequential Small-Animal PET and In Vivo Optical Imaging of Reporter Tumors
DISCUSSION
The TKGFP dual-reporter gene combined with optical and nuclear imaging modalities promotes molecular–genetic imaging and is more suitable for application in translation medicine. This combination approach has the advantage of speed and easier validation of approaches in cells or small animals and, in turn, in humans. However, the potential problems of cytotoxicity and poor temporal resolution of TKGFP may limit its use for such purposes. Previous studies have suggested that TKGFP is highly stable in mammalian cells because of the high protein stability of GFP and HSV1-tk (12,15). Our previous studies have shown that HSV1-tk is highly stable in mammalian cells (15,16); this may be due to the absence of potential proteolytic cleavage sites (pro, glu, and ser/thr–rich motif) and the presence of stabilizing residues in the N-terminal protein sequence. In contrast to HSV1-tk, GFP is a single-chain polypeptide of 238 amino acids. Most of these amino acids are found in the form of antiparallel β sheets, forming a compact barrellike structure. The compact structure makes GFP stable under a variety of conditions, including protease treatment (12). To investigate the characteristics of TKGFP, bioinformatics analysis was performed to study the TKGFP amino acid sequence. The results showed no potential proteolytic cleavage sites, and the half-life of the TKGFP was estimated to be about 30 h based on the N-end rule. Our protein stability studies also proved that TKGFP was highly stable in mammalian cells; therefore, this reporter system has low temporal resolution for monitoring the temporal dynamics of endogenous signal transduction activity or transgene expression. Furthermore, our data indicated that TKGFP as a reporter controlled by a constitutive promoter is cytotoxic because of the nuclear tropism and high stability of the protein. These problems render the TKGFP reporter system unsuitable for monitoring short-time-scale dynamic processes such as kinetic gene expression controlled by inducible promoters or a less stable protein with a more rapid turnover; thus, this system cannot be applied in translational research.
To construct a more responsive TKGFP reporter system, destabilized TKGFP was produced by inserting the leucine-rich NES sequence at the N terminus and fusing the degradation domain of MODC at the C terminus, as described previously (16,32). The insertion of the leucine-rich NES sequence at the N terminus of TKGFP inhibits the nuclear tropism of TKGFP and promotes the reporter protein export from cell nuclei to cytoplasm. These results correspond with the published results of other research groups (32). In mammalian cells, most intracellular proteins are degraded in the cytoplasm by ubiquitin-mediated proteolysis. Although TKGFP with the leucine-rich NES sequence inserted at the N terminus does not show nuclear tropism, NESTKGFP remains highly stable in the cytoplasm. Therefore, we fused the pro, glu, and ser/thr domain of MODC in NESTKGFP to further improve its high protein stability in cells. Our results demonstrated that the protein and enzyme stability of NESTKGFP:dMODC decreased in the living cells. However, the protein and enzyme stability of TKGFP:dMODC was constant during the observation period. The nuclear tropism of TKGFP:dMODC may inhibit its degradation in the cytoplasm. This involves 2 steps in the ubiquitin-mediated pathway: the addition of a chain of ubiquitin molecules to an internal lysine side chain of the target protein and the proteolysis of the ubiquitinated protein by proteasome, a large, cylindric multisubunit complex. The numerous proteasomes present in the cell cytosol proteolytically cleave ubiquitin-tagged proteins in an adenosine triphosphate–dependent process, yielding peptides and intact ubiquitin molecules (35). Our proteasome inhibition assay confirmed that the degradation of NESTKGFP:dMODC in the cells was via the ubiquitin-mediated pathway, because the rapid turnover of NESTKGFP:dMODC was inhibited by the proteasome inhibitor MG-132.
Hypoxia-responsive elements linked with luciferase reporter genes combined with optical imaging systems have been used for real-time observation of HIF-1 signal transduction activity in living subjects (36–38). However, these approaches are limited by depth penetration and the nature of tomography and cannot monitor the spatial heterogeneity of HIF-1 signal transduction activity. Furthermore, these approaches cannot be easily generalized to human applications (39). In contrast to optical reporter systems, the PET reporter system, HSV1-tk, is applicable with fully 3-dimensional tomography to monitor or quantify the intensity of transgene or endogenous gene expression (33,34). However, this reporter system has poor temporal resolution for monitoring short-time-scale dynamic processes such as kinetic gene expression controlled by inducible promoters or by a less stable protein with a more rapid turnover in vitro and in vivo (15,16). In this study, the novel mutant TKGFP (NESTKGFP:dMODC) was used to observe the temporal dynamics of HIF-1 signal transduction activity mediated by hypoxia and reoxygenation in vitro and in vivo. Our results showed that both TKGFP and NESTKGFP:dMODC as a reporter can represent the HIF-1 transcriptional activity induced by different hypoxic conditions. Reduced oxygen concentrations induced an inverse concentration-dependent activation of HIF-1 transcription and upregulation of TKGPF and NESTKGFP:dMODC expression in the NG4TL4 reporter cells. However, in comparison with NESTKGFP:dMODC, TKGFP as a reporter cannot be used for real-time monitoring of the dynamics of reoxygenation-mediated HIF-1 transcription activity in vitro and in vivo because of the poor temporal resolution of TKGFP. In a previous study, the TKGFP reporter system had been applied to study the dynamics and spatial heterogeneity of HIF-1-specific transcription activity in C6 rat glioma cells. The study involved repetitive noninvasive PET of hypoxia-induced HIF-1 transcription activity in tumors in living mice (31). However, assessment in real time seemed to be difficult because the reporter proteins were stable in cells. The TKGFP reporter system may not be suitable for monitoring the dynamic changes in HIF-1 signal transduction activity mediated by cyclic hypoxia in vitro and in vivo, because some TKGFP may have remained in the cells after reoxygenation. Our results proved this hypothesis; further, our observations confirmed that NESTKGFP:dMODC as a reporter was more suitable for observing the dynamic changes in HIF-1 signal transduction activity in vitro and in vivo.
In summary, we successfully constructed a novel mutant TKGFP (NESTKGFP:dMODC) fusion reporter gene with high temporal resolution and low cytotoxicity together with optical imaging and PET for use in real-time monitoring of the temporal dynamics and spatial heterogeneity of HIF-1 signal transduction activity mediated by hypoxia and reoxygenation in vitro and in vivo. This reporter gene imaging system can also be used to noninvasively observe other interesting endogenous signal transduction activity or transgene expression. Because of the better temporal resolution of NESTKGFP:dMODC, it is suitable for monitoring short-time-scale dynamic processes such as kinetic gene expression controlled by inducible promoters or by a less stable protein with a more rapid turnover. In contrast to other real-time reporter systems, the ability to perform translational research from a cell culture setting to preclinical animal models and, in turn, to clinical applications is a unique and powerful feature of this reporter system. Therefore, it provides a useful platform in translational molecular–genetic imaging studies.
CONCLUSION
TKGFP as a reporter has high protein stability and cytotoxicity in living mammalian cells. Its poor temporal resolution limits its application in the in vitro and in vivo monitoring of short-time-scale dynamic processes such as kinetic gene expression controlled by inducible promoters or by a less stable protein with a more rapid turnover. TKGFP with reduced protein stability and cytotoxicity in living mammalian cells was produced by inserting the leucine-rich NES sequence at the N terminus and fusing the degradation domain of MODC at the C terminus. NESTKGFP:dMODC as a transcription reporter combined with optical imaging and PET allows the direct monitoring of transcription induction and easy determination of its association with other biochemical changes in vitro and in vivo.
Acknowledgments
We thank the Molecular and Genetic Imaging Core/NRPGM, Taiwan, for technical support and May Lee for editorial assistance. This study was supported by grant 97-2314-B-039-001-MY2 from the National Science Council, Taipei, Taiwan, and grant CMU96-110 from China Medical University, Taichung, Taiwan.
Footnotes
-
COPYRIGHT © 2009 by the Society of Nuclear Medicine, Inc.
References
- Received for publication December 16, 2008.
- Accepted for publication August 25, 2009.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}