


Abstract
p-131I-iodo-l-phenylalanine (131I-IPA) is a tumor-specific amino acid derivative that demonstrated antiproliferative and tumoricidal effects on experimental gliomas. This study tested the efficacy of 131I-IPA combined with external beam photon radiotherapy as a new therapeutic approach against gliomas. Methods: Glioma cells derived from the rat F98 glioma or human Tx3868 or A1207 glioblastoma cell lines were stereotactically inoculated into the brains of Fischer 344 rats or RNU rats. Tumor formation was verified radiologically. On day 8, groups of glioma-bearing rats of each tumor model underwent whole-brain radiotherapy with 8 Gy, an intravenous administration of 131I-IPA (30 MBq), or combined treatment, aiming for a total of 12 rats per group. Another 12 animals were treated with physiologic saline and served as control. Results: Control rats had a combined median survival (±SD) of 21 ± 6 d. All revealed metabolically and histologically large tumor masses. Efficacy of radiotherapy alone or a monotherapy with 30 MBq of 131I-IPA was statistically insignificant on the syngeneic Fischer–F98 model (P ≥ 0.45 and P = 0.10, respectively). In contrast, a subset of long-term survivors (>120 d) was observed in RNU rats bearing Tx3868 and A1207 glioblastoma xenografts (18%−25% and 35%−45% for radiotherapy and 131I-IPA, respectively). Combined 131I-IPA and radiotherapy treatment significantly prolonged median survival for the syngeneic Fischer–F98 glioma model (P < 0.01) and human glioblastoma–bearing RNU rats alike (P < 0.05). On day 120 after monotherapy with 131I-IPA, 45% of the RNU rats were still alive, but after 8 Gy of photon radiotherapy only 18%−25% of the RNU and none of the Fischer rats survived. In comparison, 55%−75% survival rates were registered after combined treatment on day 120 for all animal models. Conclusion: These data convincingly demonstrated that systemic radionuclide therapy with 131I-IPA combined with external photon radiotherapy is a safe and highly effective treatment for experimental gliomas, which may merit a clinical trial to ascertain its potential in patients with gliomas. Because only a low 131I-IPA activity and low radiotherapy doses were applied, further optimizations including higher radiation doses and conventional fractionated radiotherapy are warranted.
- malignant gliomas
- targeted radionuclide therapy
- external photon radiotherapy
- combination treatment
- radiosensitivity
Despite advances in the current standard therapies—including surgery, radiotherapy, and chemotherapy—the prognosis for patients with malignant glioma remains poor (1,2). The diffuse infiltration of these neoplasms means that not even the smallest surgical instruments are fine enough to bypass the healthy neurons and resect only the tumor cells (3). Additionally, most malignant gliomas are highly resistant to external radiation or systemic chemotherapy (4–6). Therefore, overall survival (OS) generally ranges from a few months to about 1 y for glioblastoma multiforme, the most common malignant glioma (1–4). To overcome these dismal prospects, various experimental therapies have been administered, including gene therapy, antisense treatment, boron neutron capture, locoregional radioimmunotherapy, ligand–toxin conjugate administration, or 5-aminolevulinic acid photodynamic therapy; methods for sensitizing glioma cells to apoptosis induction; or methods aiming at different targets such as the coagulation system, to name only a few (7–13). Unfortunately, all these efforts have failed to appreciably increase the median OS of patients with gliomas.
The finding that malignant brain tumors accumulate amino acids more avidly than do healthy brains has led to the development of amino acid–based radiopharmaceuticals for detecting brain neoplasms using PET and SPECT (14–17). The iodinated amino acids p-123I-iodo-l-phenylalanine (123I-IPA) and p-124I-iodo-l-phenylalanine have been used clinically for this purpose (18–20). Both these phenylalanine derivatives actively cross the blood–brain barrier after intravenous administration and accumulate specifically in gliomas, presumably via the amino acid transporters l and ASC (system ASC amino acid transporter for neutral amino acids), which are overexpressed in malignant glioma cells (21–23). A specific accumulation and prolonged retention of the radiopharmaceuticals were demonstrated clinically for low-grade and high-grade gliomas alike. At the same time, the uptake of these agents in normal brain parenchyma and nonneoplastic lesions remained moderate (18–20). Therefore, we postulated that when administered systemically, l-phenylalanine that was conjugated to a α- or β-emitting radionuclide such as 211At or 131I could selectively deliver an effective radiation dose to malignant glioma cells. Consequently, we developed p-131I-iodo-l-phenylalanine (131I-IPA), which—when used as a single agent—was tumoricidal to glioma cells and improved survival of rats with cerebral C6 gliomas (24). Besides the therapeutic β-emission, 131I-IPA seems to act synergistically tumoricidal in connection with external irradiation (S. Samnick, unpublished data, 2005). The unique properties of 131I-IPA—properties that combine a specific and sustained tumor accumulation and an intrinsic cytostatic and radiosensitizing effect—make 131I-IPA highly attractive as a therapeutic probe against gliomas.
In this study, we have tested the therapeutic efficacy of systemic targeted radionuclide therapy with 131I-IPA combined with external beam radiotherapy on experimental gliomas.
MATERIALS AND METHODS
Cell Lines, Cell Cultures, and Implantation Solutions
The primary human A1207 glioblastoma cell line was provided by Dr. Reiner Class of Symbiotec GmbH, and the rat F98 glioma cell line was from the American Type Culture Collection. The human Tx3868 glioma cell line, derived from a primary human glioblastoma multiforme, was provided by Dr. Eckard Meese of the Institute of Human Genetics, University of Saarland.
A1207 and F98 cells were grown in Dulbecco's modified Eagle's medium (DMEM) (PAA) and Tx3868 cells in RPMI 1640 medium supplemented with glucose (4.5 g/L), 10% fetal bovine serum, l-glutamine (300 mg/L), and 1% penicillin (100 IU/mL)/streptomycin (100 μg/mL) (PAA). Cells were maintained in a humidified 5% CO2 incubator at 37°C and passaged routinely.
The implantation solutions were prepared by trypsinization of the subconfluent cell cultures using a solution of 0.05% trypsin in phosphate-buffered saline (PBS) containing 0.02% ethylenediaminetriacetic acid (PAA), followed by resuspension in fetal bovine serum–free DMEM (F98 and A1207) or PBS (Tx3868). A trypan blue exclusion test was performed on a hemocytometer to assess cell viability before implantation; the viability was greater than 99% in all cases.
Animals and Tumor Implantation
Male Fischer 344 rats and RNU rats (immunodeficient athymic Rowett nude rat), weighing 210–240 g and acquired from Charles River Laboratories, were used for tumor implantation. Throughout the experiments, the rats were maintained in the animal facility of the Saarland University Medical Center. The experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (revised 1996) (25) and in compliance with the German law on the protection of animals. All experiments were approved by the local district government of Saarpfalz-Kreis (AZ: K 110/180–07, 2006).
Our tumor cell–implantation protocol was formulated on the basis of the results of serial preliminary experiments performed to identify the implantation coordinates and cellular suspension characteristics that would lead to a valid, reproducible model with an optimal time frame from implantation to death in the absence of active treatment.
For tumor implantation, the animals were anesthetized by an intraperitoneal injection of ketamine (70 mg/kg of body weight) and xylazine (Rompun 2%; Bayer Health Care) at a dose of 20 mg/kg of body weight. The rats' eyes were coated with 5% dexpanthenol (Bayer Vital) to prevent keratitis. Animals then were placed in a stereotactic fixation device, and a midline incision was made in the scalp to expose the skull, after which the bregma was identified and exposed. A small hole was drilled into the skull, 1 mm anterior and 3 mm lateral to the right of the bregma. Then, A12078 human glioblastoma cells (105 cells in 5 μL of PBS), F98 rat glioma cells (104 cells in a volume of 5 μL of serum-free DMEM), or Tx3868 human glioma cells (105 cells in 5 μL of PBS) were inoculated stereotactically into that hole to a depth of 5 mm, using a microsyringe (SGE Micro volume syringe; Hamilton). After the syringe was slowly withdrawn to minimize backflow of the cell suspension, the craniectomy was resealed with bone wax and the scalp closed. Animals were allowed to recover from the procedure and were given food and water ad libitum afterward. They were observed daily for signs of raised intracranial pressure or for focal neurologic signs (hemiparesis, ataxia). Tumor formation was verified by MRI between day 8 and day 14 after tumor cell inoculation using a 1.5-T system (Magnetom Sonata; Siemens). To maximize the signal-to-noise ratio, animals were positioned in a dedicated wrist coil placed directly on the head. Coronal T2-weighted images and contrast-enhanced T1-weighted images were obtained in analogy with the imaging protocol described previously (24).
Synthesis of 131I-IPA
131I-IPA was prepared by isotopic radioiodination in the presence of Cu(II). Specifically, a solution of sodium 131I-iodide (370–1,000 MBq) in 0.05N NaOH (Amersham Healthcare) and 5 μL of aqueous Na2S2O5 (4.0 mg Na2S2O5/mL) was evaporated to dryness by passing a stream of nitrogen through a reaction vessel at 90°C, followed by the addition of 150 μL of p-iodo-l-phenylalanine solution (0.25 mg/mL 0.1N H3PO4), 20 μL of l-ascorbic acid (10 mg/mL of water), and 20 μL of aqueous Cu(II) sulphate (0.10 mol/L). The reaction vessel was heated for 60 min at 165°C and cooled, and the mixture was diluted with water (250 μL). 131I-IPA was separated from radioactive impurities and byproducts by high-performance liquid chromatography, using water:ethanol:acetic acid (89:10:1; v/v) as the mobile phase. The fraction containing 131I-IPA was collected into a sterile tube, buffered with 0.5 M PBS (pH 7.0; Braun), and sterile-filtered through a 0.22-μm sterile membrane (Millex GS; Millipore) to an isotonic and injectable radiopharmaceutical for in vivo investigations. 131I-IPA was obtained in 88% ± 10% radiochemical yield. The specific activity of 131I-IPA used for in vivo studies was 0.7–1.0 GBq/μmol.
Animal Treatments
For each of the 3 tumor models, 48 rats (confirmed by MRI as bearing gliomas) were randomized to receive 1 of 4 treatments starting at day 8 after tumor cell implantation, including the external radiotherapy, the 131I-IPA, the combined 131I-IPA and radiotherapy, and the untreated control groups. Each group consisted of 12 rats, aiming for a total of 144 rats for all animal models.
In a preliminary pilot evaluation in healthy rats, we found that the application of external radiation doses greater than 30 Gy resulted in acute brain damage (data not shown). For this reason, the evaluation of the radiotherapy on glioma xenografts on rats using radiation doses up to 60 Gy as clinically used in humans was not applicable in this study. The application of 8 Gy of radiotherapy as whole-brain irradiation does not damage the rat brain and resulted in a modest OS of rats with cerebral glioma (26). Therefore, whole-brain radiotherapy using an 8-Gy photon irradiation was chosen for the assessment of the efficiency of the combined treatment, compared with single treatments alone.
The radiotherapy monotherapy group received an external 8-Gy whole-brain photon irradiation as daily 4-Gy fractions on days 8 and 9 after tumor cell implantation. The irradiation was performed at a dose rate of 2 Gy/min under ambient conditions using a 6-MeV linear accelerator (Mevatron MXE; Siemens). The focus-isocenter distance was 100 cm. The remainder of the rat body was shielded from the direct beam using lead, so that the animals were exposed only to minor scattered radiation outside the brain.
The 131I-IPA monotherapy group was intravenously administered a total activity of 30 MBq of 131I-IPA via a tail vein in two 15-MBq portions within 7 d—that is, on days 8 and 15 after tumor cell inoculation. This regimen was chosen according to the kinetics of 131I-IPA in human glioma, as assessed previously in a mouse xenograft model and in a patient with this tumor (20,27).
The combined modality–treatment group received a 8 Gy of photon irradiation, followed by an intravenous administration of 131I-IPA in 2 portions as described above, starting at 24 h after the end of the radiotherapy.
The control group was treated with 250 μL of physiologic saline only, administered via a tail vein on day 8.
Animals were anesthetized by isoflurane inhalation during all treatments, including the saline.
Evaluation of Treatment Efficacy
Treatment efficacy for the radiotherapy monotherapy, 131I-IPA monotherapy, and combined modality–treatment groups, compared with that for the control animals, was evaluated on the basis of the median OS. Animals were observed until ethical sacrifice, until death from any other cause, or through day 180 after tumor cell inoculation. Animals that died before day 4 after treatment (there were 2 Fischer 344 rats with F98 glioma and 1 Tx3868- and 1 A1207-bearing RNU rat) were excluded from the analysis and substituted by new tumor-bearing rats. In all cases, 12 glioma-bearing rats were evaluated in each treatment arm.
Histopathologic and Metabolic Analyses
The brains of all sacrificed rats underwent histopathologic and metabolic analyses as described below, and the results of those analyses were compared with the results of an identical analysis of a representative brain specimen obtained from a control and sham animal of the corresponding rat model after autopsy.
For the histopathologic analysis, rat brains were taken after autopsy and fixed in 4% neutral-buffered formalin, cut in coronal slices of 2–3 mm, and embedded in paraffin wax. Sections were stained with hematoxylin and eosin and Verhoeff-van Gieson stain and examined according to previously published methods (24).
Regional brain metabolism was analyzed by bioluminescence as described previously (28). Briefly, the frozen brains were removed from the bone skull in a cold box at −20°C. Coronal sections (20 μm thick) were prepared in a cryostat at −20°C. Alternate sections were taken for imaging levels of adenosine triphosphate (ATP), glucose, and lactate and then heated to 95°C to inactivate endogenous enzymes. A solution was prepared containing all enzymes, coenzymes, and cofactors necessary for the substrate-specific bioluminescence reaction but without the substrate of interest (ATP, glucose, or lactate). This solution was frozen in a block, and 60-μm-thick sections were cut in a cryostat. A freeze-dried and heat-inactivated brain section was then covered with a 60-μm section of the frozen enzyme block, and this open sandwich was placed on a photographic film (Agfapan 100 ASA; Agfa) for the recording of bioluminescent light emitted by the brain sections after warming to room temperature. Exposure times were 0.5 min for ATP and 5 min for glucose and lactate. The bioluminescence-induced blackening of films was quantified by computer-assisted densitometry using image-analysis software (Image-Pro Plus; Media Cybernetics) and expressed as optical density (OD). The OD was shown to correlate linearly with substrate concentrations measured quantitatively in tissue extracts using conventional enzymatic-fluorometric techniques (28).
Statistical Analysis
The statistical significance of differences among experimental groups was determined by Student t test. Survival was described by Kaplan–Meier analysis. The threshold of statistical significance was set at P level of less than 0.05. Statistics and fitting of experimental curves were performed with the program SigmaStat (Jandel Scientific).
RESULTS
Implantation procedures were well tolerated by both RNU and Fischer 344 rats. Survival of untreated rats was inversely related to the number of glioma cells stereotactically inoculated into the brain (data not shown). In untreated animals, mean ± SD survival after tumor cell inoculation was 19 ± 4 d in Fischer 344 rats implanted with 1 × 104 F98 glioma cells, 20 ± 4 d in RNU rats implanted with 1 × 105 A1207 human glioblastoma cells, and 23 ± 5 d in RNU rats implanted with 1 × 105 Tx3868 human glioma cells (n = 12 each). The analysis of brain specimens during the optimization procedures revealed the presence of large tumor masses in rat brains at day 10 after cell inoculation. Histomorphologically, the F98 and Tx3868 gliomas were large, with necroses and numerous mitotic figures. A representative MR image of a Fischer rat brain at day 10 after inoculation of 104 F98 glioma cells and the corresponding brain slice confirming tumor are presented in Figure 1. Coronal hematoxylin and eosin slices of RNU rats bearing human A1207 and Tx3868 glioblastomas are shown in Figures 1D–1F.
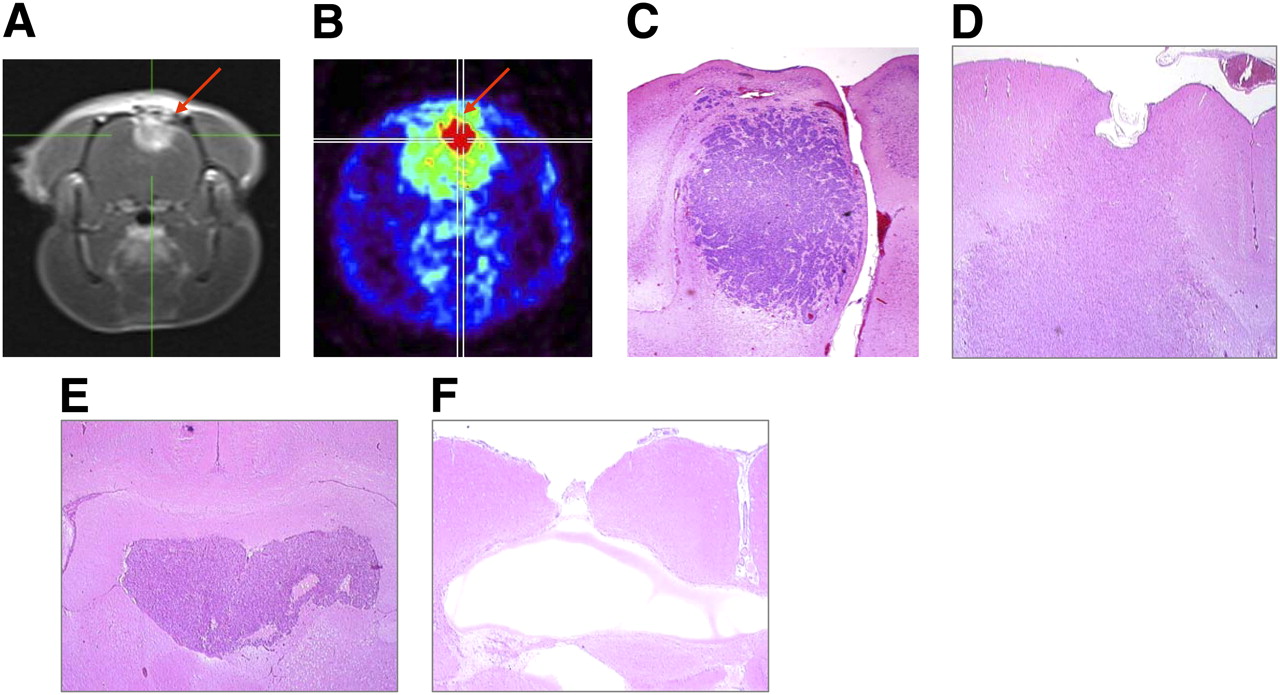
Coronal MRI (T1-weighted) scan of Fischer rat brain assessed at day 10 after inoculation of 104 F98 glioma cells, demonstrating F98 glioma (A, arrow) that was subsequently confirmed by small-animal PET after injection of 10 MBq of 18F-FDG (B, arrow). Corresponding histomorphologic analysis of isolated brain confirms tumor (C). Analog representative brain slice of A1207 glioma–bearing RNU rat scarified at day 10 after tumor implantation. At brain–tumor border, glioma cells grow diffusely into surrounding parenchyma. Gap at top of brain represents bone wax (D). (E) Illustration of brain slice of RNU–Tx3868 glioma model showing tumor spreads along ventricle and for most part sharp demarcation. (F) Brain from cured RNU rat with Tx3868 glioma sacrificed at day 120 after combined 131I-IPA treatment and radiotherapy. In addition to lesion defect, no residual tumor cells were detectable. (hematoxylin and eosin, ×20)
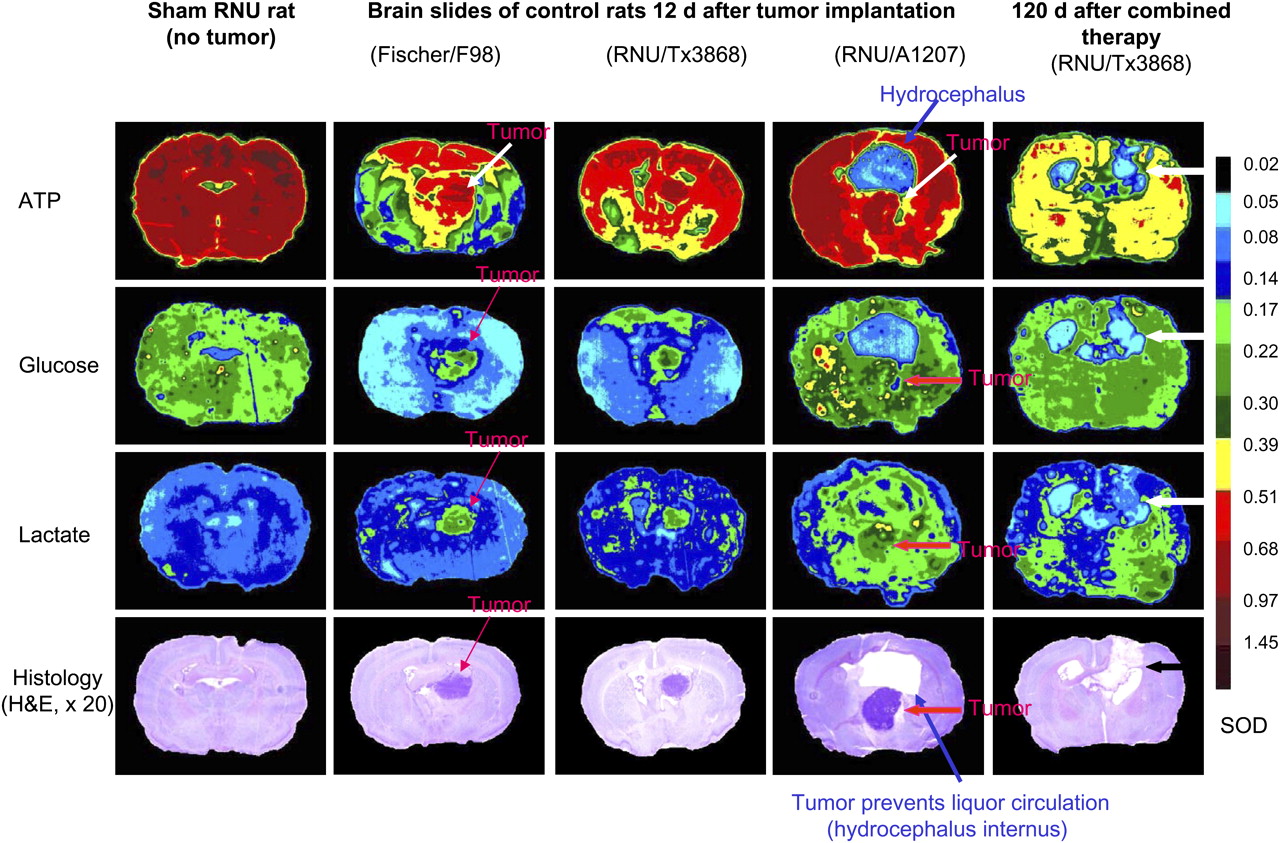
The F98 glioma and the human Tx3868 glioblastoma consistently maintained an extensive infiltrative proliferation, and the tumors often extended along the implantation tract all the way to the surface of the brain, with leptomeningeal spread. With respect to the behavior in vivo, the only notable difference among the F98, Tx3868, and A1207 glioma cell lines was a consistent tendency of A1207 glioblastoma to form more sharply circumscribed tumor masses and to develop more extensive edema in the surrounding brain. Metabolically, ATP, glucose, and lactate increased significantly (P < 0.05) in untreated tumor, as compared with the surrounding tissue, especially in the brain of Fischer–F98 and RNU–Tx3868 models (Fig. 2).

Representative examples of metabolic and histologic outcome in F98–Fischer rat and in Tx3868- and A1207-bearing RNU rats after combined treatment based on external radiotherapy with 8 Gy of photon irradiation and 131I-IPA (30 MBq). Arrows indicate tumor or hydrocephalus. A1207 glioblastoma was characterized by consistent tendency to form edema in surrounding brain. Shown is substance defect in cured RNU rat bearing Tx3868 glioma without evidence of tumor (black arrow). H&E = hematoxylin and eosin; SOD = scale of OD.
In the first experiment, we tested the efficacy of radiotherapy alone and of monotherapy with 30 MBq of 131I-IPA on syngeneic and xenografted orthotopic gliomas in rats. The application of 8 Gy of photon radiotherapy alone resulted in no significant (P ≥ 0.45) increase in survival for the Fischer–F98 glioma model, as compared with untreated control rats. In particular, no F98 glioma–bearing Fischer rat was alive on day 52 after the application of 8 Gy of photon irradiation as monotherapy. In the same time, the OS was significantly (P < 0.02) improved in RNU rats with intracerebral human A1207 and Tx3868 glioblastomas after radiotherapy. A comparison of both monotherapies showed that the systemic application of 30 MBq of 131I-IPA was more effective as a radiotherapy with 8 Gy in terms of survival on Fischer–F98, RNU–Tx3868, and RNU–A1207 glioma models. Three months after the monotherapies, the portion of RNU rats with human A1207 and Tx3868 glioblastoma still alive after 8 Gy of radiotherapy was a median of 32% ± 6%, compared with 55% ± 8% after radionuclide therapy with 30 MBq of 131I-IPA. No Fischer 344 rat bearing F98 gliomas was censored on day 52 after radiotherapy alone. From day 120 onward, survival time in response to the individual monotherapies decreased rapidly (Fig. 3).
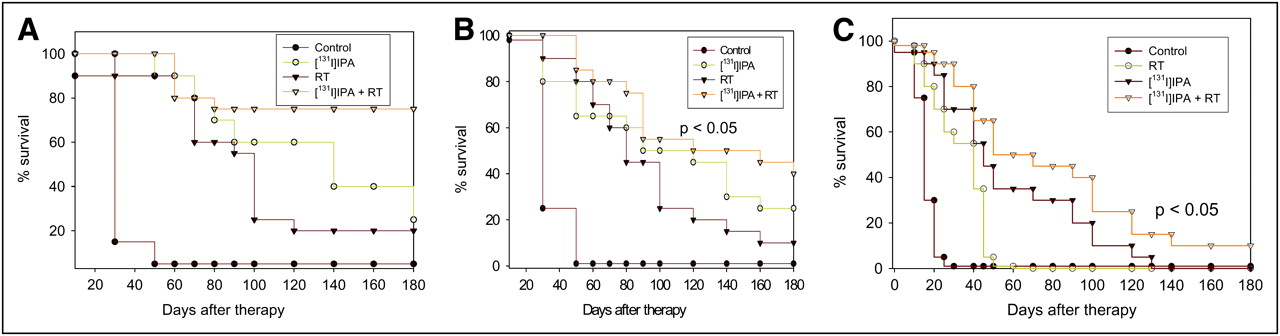
Kaplan–Meier survival curves for RNU rats with human Tx3868 (A) and A1207 (B) glioblastomas and for F98 glioma–bearing Fischer rats (C) subjected to physiologic saline (control) or to 8 Gy of photon radiotherapy, 131I-IPA (30 MBq), and combined radiotherapy and 131I-IPA treatment. n = 12 animals of each glioma model per treatment group for each treatment modality.
In the second experiment performed in parallel, we compared the efficiency of the single treatments alone with the combined 131I-IPA and radiotherapy treatment. Overall, the combined treatment based on radiotherapy and 131I-IPA was more effective in terms of local tumor control and prolonged median survival (P ≤ 0.01) for the F98 glioma–bearing Fischer rats and human Tx3868 and A1207 glioblastoma xenografts in RNU rats alike. In particular, the combined treatment enhanced the therapeutic efficacy of the radiotherapy on glioma-bearing rats by a factor of 1.3–4.0; the median survival of rats treated with 8 Gy of radiotherapy and 131I-IPA showed a statistical difference (P = 0.002), compared with rats treated with 8 Gy of radiation alone. The Kaplan–Meier survival curves of Fischer rats bearing F98 and of RNU rats with Tx3868 and A1207 gliomas and subjected to the indicated treatments, compared with untreated controls, are presented in Figure 3. Combined 131I-IPA treatment and radiotherapy was particularly effective on RNU rats with cerebral Tx3868 and A1207 glioblastomas. At day 120 after monotherapy with 131I-IPA, at least 45% of the RNU rats were still alive, but after 8 Gy of photon radiotherapy only 18%−25% of the RNU and none of the Fischer 344 rats survived. In comparison, 55%−75% survival rates were registered after combined 131I-IPA treatment and radiotherapy on day 120 for all animal models. Figure 1F shows an RNU rat with a human Tx3868 glioma after successful combined treatment. Histomorphologically, no tumor cells were left. Instead, there was a huge gap in the cortex. Surprisingly, neurologic deficits were not remarkable. Median survival times at day 120 after individual treatments are giving in Table 1.
Median (±SD) Survival at 120 Days after Radiotherapy, Radionuclide Therapy with 131I-IPA, and Combined Radiotherapy and 131I-IPA Treatment
Bioluminescent analysis not only revealed differences within the regional ATP and glucose metabolism in the brains of Fischer–F98 rats, compared with the RNU glioma models, but also compared the RNU–Tx3868 with RNU–A1207 glioma model. In all 3 animal models, the OD of all 3 studied analytes (ATP, glucose, and lactate) was decreased in treated versus control animals. The reduction from control levels was greatest in those animals given combination therapy, in whom the analytes had an OD significantly (P < 0.05) lower than that in the animals given 1 of the active monotherapies. In addition, substance losses were determined in treated rat slices after autopsy, signaling that the combination of 131I-IPA and radiotherapy is the most effective antitumor strategy (Fig. 2).
DISCUSSION
We have previously reported on the efficacy of radionuclide therapy with 131I-IPA on intracranial C6 glioma models (24). Although the rat C6 glioma model has been used for more than 30 y, the extent to which it recapitulates the characteristics encountered in human gliomas remains controversial (29,30). For this reason, we selected for this study 3 alternative in vivo glioma models in rats, all of which have been described to adequately mimic the behavior of human gliomas (26,31). An important challenge, however, remains to further increase dose effect at the tumor side after systemic application of 131I-IPA. Various recent studies have demonstrated that the greatest potential for systemic radionuclide therapy will not be as a monotherapy but as therapy integrated into an established treatment regimen such as external radiotherapy or chemotherapy (13,32–35). These fundamental findings and data from our preliminary investigation in primary human glioblastoma cell lines prompted us to test the therapeutic efficacy of radionuclide therapy with 131I-IPA in combination with external beam photon radiotherapy, which forms part of the current standard therapy protocols for gliomas. The rationale behind combining 131I-IPA with radiotherapy is that the 2 modalities together may deliver more radiation dose to the targeted tumor than either modality alone. In fact, various studies have demonstrated that if radiotherapy precedes systemic targeted radionuclide therapy, uptake of the radiolabeled probe significantly increases (35). If systemic radionuclide therapy precedes radiotherapy, the low-dose-rate irradiation via radionuclide therapy may potentiate tumoricidal effects of subsequent radiotherapy through mechanisms that put tumor cells in a more sensitive phase or through a phenomenon termed protracted-exposure radiosensitization (36).
The present investigation confirmed in 3 additional rat models earlier findings in the intracranial C6 glioma model (24) that systemic radionuclide therapy with 131I-IPA by itself improves OS relative to that seen in control rats. However, in these experiments, when given in the same regimen, 131I-IPA monotherapy was less effective in improving OS in the syngeneic Fischer 344–F98 glioma model than in the carcinogen-induced C6 glioma model in our previously published experiments (24). This difference in tumor response cannot be clearly explained so far. One cause could be the reported radioresistance of the F98 glioma line, compared with the C6 glioma line (26). By contrast, in control rats 131I-IPA alone was highly effective, compared with control treatment in Tx3868 and A1207 glioblastoma–bearing RNU rats, in extending OS in tumor-bearing rats.
The major finding in the present investigation is that the systemic radionuclide therapy with 131I-PA combined with external beam radiotherapy significantly (P < 0.01) prolonged median survival times and was more effective than any monotherapy alone in F98 glioma–bearing Fischer rats and in RNU rats with human glioblastomas alike. Kaplan–Meier analysis showed that in the syngeneic Fischer–F98 glioma model and in the xenotransplanted human A1207 or Tx3868 glioblastoma–RNU rat models, combined treatment improved median OS by factors of 5–10 over that seen in controls, over that seen in 131I-IPA monotherapy groups, and over that seen in radiotherapy-only groups. Metabolically, bioluminescent analysis not only revealed differences within the regional ATP and glucose metabolism in the brains of Fischer–F98 rats, compared with the RNU glioma models, but also compared the RNU–Tx3868 model with RNU–A1207 glioma model and relative to control and sham rats. The different metabolic properties of the 3 glioma models could explain the differences in treatment effects. In particular, the ATP and glucose metabolism and the presence of hydrocephalus correlate negatively with therapeutic outcome and survival rate of treated rats. The fact that the metabolic profile recovered to control might attest to the effectiveness of the combined treatment strategy. No toxic effects were observed clinically in treated animals. Moreover, in autopsies after 3 mo of follow-up the survivors in the combination-therapy group showed little if any recurrent tumor growth. This is, to the best of our knowledge, the best demonstration that a systemically (intravenously) applied radionuclide therapy by a single amino acid combined with external radiotherapy improves therapy efficiency and prolongs survival in malignant gliomas. Our finding of greater efficacy of 131I-IPA within a combination regimen than as monotherapy provides additional evidence that, ultimately, the greatest potential for systemic radionuclide therapy will not be as monotherapy but will be integrated into established multimodality regimes such as external beam radiotherapy, chemotherapy, or both.
A potential problem with systemically administrated radionuclide therapy consists of toxicity caused by the portion of the radiation dose that does not reach the target tumor cells but instead reaches the whole body and susceptible organs such as the bone marrow, liver, urogenital system, and thyroid gland. The present study did not histologically evaluate the effects of the active treatments outside the brain. However, in the long-term surviving animals in the 131I-IPA–alone and the 131I-IPA and radiotherapy groups, no clinical adverse events were observed that were suggestive of organ or tissue damage in those body compartments. Moreover, previous investigations of 131I-IPA monotherapy found no histomorphologic abnormalities in the liver, kidneys, and thyroid of treated animals, and no rat had clinical signs of malfunction of any organ (24). Additionally, prior biodistribution studies and radiation dose estimates based on studies in animals and humans revealed that 123I-IPA–131I-IPA is exclusively eliminated by urine, and the highest absorbed doses were determined in the urinary bladder wall and in the kidneys (18,21,37,38). Radioactivity accumulation in bone marrow, the urogenital system, and the thyroid gland is rather tolerably low. According to the ICRP 60, the effective dose resulting from 131I-IPA was 0.13 ± 0.07 mSv/MBq, indicating that an intravenous application of 131I-IPA should result in a radiation dose lower by a factor of 2–3 than that delivered by conventional radioimmunotherapy (39). The present study and these previous studies also show that intravenous 131I-IPA does not share the toxicity of conventional antiglioma chemotherapies.
One limitation of this study was that it tested only relatively low therapeutic doses. In a preliminary pilot evaluation in healthy rats, we found that external whole-brain photon irradiation with radiation doses greater than 30 Gy damaged healthy rat brains and reduced the median survival of the rats (data not shown). Therefore, the application of high-dose radiation up to 60 Gy, as clinically used in humans, was not applicable for this study that focused on rat brain. In contrast, whole-brain radiotherapy with 8 Gy alone did not lead to measurable brain damage and resulted in a modest increase in survival rate for rats with cerebral glioma (26). For this reason, whole-brain radiotherapy using 8 Gy of photon irradiation was chosen for a better analysis of the efficiency of the combined 131I-IPA and radiotherapy treatment, compared with single treatments alone. However, an increase of radiation doses might have a greater effect on local tumor control (40), and the impact of an 131I-IPA activity increase will be elucidated in ongoing studies by our group.
Despite application of relatively low radiotherapy doses, our data convincingly demonstrated that 131I-IPA combined with radiotherapy is more effective than single treatments alone and improved OS in rats with cerebral gliomas significantly. If also confirmed by standard fractionated radiotherapy, systemic radionuclide therapy with 131I-IPA combined with a conventional external radiotherapy up to 60 Gy should be considered a promising option for the treatment of human gliomas, encouraging validations in patients.
CONCLUSION
131I-IPA combined with external irradiation (radiotherapy) improves OS relative to the 2 treatment modalities as monotherapy in experimental gliomas. The combination of 131I-IPA with radiotherapy would be a logical treatment to bring 131I-IPA into clinical trials for human gliomas. The reasons for this are severalfold. First, radiation and chemotherapy are the gold standard for the treatment of human gliomas, and any novel therapy will be measured against them. Second, data of our study demonstrated that 131I-IPA enhanced the efficiency of radiotherapy. In the meantime, further efficacy optimization strategies should be pursued experimentally, including the application of radiotherapy in conventional fractionated regimens or use of methods aiming to increase target doses and maximize dose effects.
Acknowledgments
We express our deep appreciation to Soja Hoffmann and Peter Hidiruglo for animal care and to Georg Blass for technical assistance. This work was supported by a grant from the Dr. Mildred Scheel Stiftung für Krebsforschung (BN: 106940).
Footnotes
-
COPYRIGHT © 2009 by the Society of Nuclear Medicine, Inc.
References
- Received for publication May 25, 2009.
- Accepted for publication August 28, 2009.





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.