


Abstract
Macrophages play an important role during the development and progression of atherosclerotic plaques. αvβ3 integrins are highly expressed by macrophages; thus, targeting αvβ3 may allow targeting of culprit macrophage-loaded atherosclerotic lesions in vivo. Methods: An αvβ3-targeted Arg-Gly-Asp (RGD) peptide was labeled with the cyanine 5.5 (Cy 5.5) dye and applied to image atherosclerotic plaques in apolipoprotein E–deficient mice. Results: The peptide–dye conjugate binds to αvβ3 integrin–positive RAW264.7 macrophages with high affinity. Competition experiments confirmed binding specificity of the probe. A significant fluorochrome accumulation in atherosclerotic plaques was demonstrated 24 h after injection by fluorescence reflectance imaging, which was blocked with high efficiency by competition with the unlabeled peptide. Conversely, the nonconjugated dye revealed only a minor fluorescence signal in the plaques. Fluorescence microscopy revealed colocalization of the probe with macrophages in the plaque of a mouse model for accelerated atherosclerosis, which was corroborated in human carotid artery specimens. In addition to macrophage-associated signals, binding of the probe to the neointima or elastica of the arteries was observed. Conclusion: RGD-Cy 5.5, combined with near-infrared optical imaging methods, allows the specific imaging of αvβ3-integrin expression on macrophages recruited to vascular lesions and may serve to estimate macrophage-bound inflammatory activity of atherosclerotic lesions.
Atherosclerosis is a chronic inflammatory disease affecting the arterial vessel wall (atherosclerotic plaques). Atherosclerotic vascular lesions might progress into unstable plaques, which are prone to rupture, potentially resulting in occlusion of the affected vessel, which causes myocardial infarction or stroke. To target rupture-prone plaques therapeutically, imaging of the vessel wall, and especially of components of atherosclerotic plaques, is of increasing importance. Luminography (e.g., angiography) cannot sufficiently detect these lesions, because the plaque composition, not the resulting degree of stenosis, critically determines the stability of the lesion (1). Plaque rupture might occur in the course of plaque-weakening processes such as degradation of the fibrous cap by matrix metalloproteinases (MMPs) being secreted by the macrophages (2). MMPs have already been used as molecular targets for successful preclinical molecular imaging of atherosclerotic plaques (3). Lipid-rich macrophages are one of the main components of the lipid core of the plaque and contribute to plaque weakening and rupture and, by virtue of their high levels of membrane-bound tissue factor, to the resulting thrombotic vessel occlusion as well (4). Furthermore, active and dynamic recruitment of blood-derived monocytes/macrophages proportionally correlates with plaque lesion size and symptom severity (5–8). Interestingly, monocyte-derived macrophages express high levels of integrins. Molecular imaging of integrins may, therefore, allow for noninvasive assessment of macrophage infiltration (9). Moreover, integrins constitute an important class of cell-adhesion receptors responsible not only for cell–matrix and cell–cell adhesion but also for inside–out and outside–in signal transduction. Integrins are involved in a variety of biologic processes such as angiogenesis, thrombosis, inflammation, osteoporosis, cancer, and atherosclerosis (10). One of the most relevant integrins, αvβ3, is expressed on almost all cells originating from the mesenchyme and on a variety of blood vessel cells (e.g., endothelial cells, smooth muscle cells [SMCs], fibroblasts, and platelets (11)) and binds to many different ligands, including several extracellular matrix proteins such as vitronectin, fibronectin, osteopontin, fibrinogen, and von Willebrand Factor (12,13). Specifically, the αvβ3 integrin is highly expressed in macrophages infiltrating atherosclerotic plaques and, along with membrane type-1 matrix metalloproteinases (MT-1 MMP), associates with the C terminus of the proteolytic active MMP-2 (14,15). This complex localizes MMP-associated proteolytic activity to the cell surface of, for example, macrophages, which may ultimately result in plaque destabilization.
The purpose of this study was to evaluate if the recently developed αvβ3-sensing, target-specific fluorophore Arg-Gly-Asp (RGD)–cyanine 5.5 (Cy 5.5) (16) binds to macrophages in atherosclerotic plaques and could thereby facilitate visualization of macrophage infiltration into plaques in a model of accelerated vascular lesion formation in apolipoprotein E–deficient (ApoE−/−) knock-out mice, using near-infrared fluorescence reflectance imaging (FRI).
MATERIALS AND METHODS
Labeling of Cyclo[Cys-Arg-Gly-Asp-Cys]-Gly-Lys
Peptides were labeled as described elsewhere (16). Briefly, a cyclic RGD peptide was labeled with Cy 5.5 dye by the stirring of both educts in the dark, followed by high-performance liquid chromotography purification. As reported previously, the identity of the labeled product was confirmed by high-resolution electrospray mass spectrometry (16). Before intravenous application, the integrity of the tracer was checked using photometry and fluorometry (U3310 UV/VIS spectrophotometer and F-4500 fluorescence spectrometer, respectively; Hitachi).
Cell Lines
The mouse monocytic macrophagelike cell line RAW264.7 (TIB-71; American Type Culture Collection), human melanoma cells M21 (16), and human adenocarcinoma cell line MCF-7 (HTB-22; American Type Culture Collection) were cultured in RPMI 1640 (Invitrogen Corp.) supplemented with 10% fetal calf serum, penicillin, and streptomycin. M21 and MCF-7 cells were grown routinely in a monolayer culture at 37°C in a 5% CO2 humidified air atmosphere. Murine peritoneal RAW264.7 cells were grown to 80% confluence at 37°C in a humidified atmosphere containing 5% CO2 and 95% air, washed twice with fresh medium, recovered by scraping, and suspended in fresh medium at a density of 106/mL. Cell viability was checked via trypan blue (Sigma-Aldrich Chemie GmbH).
Fluorescence-Activated Cell Sorter (FACS) Analysis by Flow Cytometry
Cells were washed twice with phosphate-buffered saline (PBS) without Ca2+ and Mg2+ and harvested in 5 mL of ice-cold Versen buffer (13.7 mM sodium chloride [NaCl], 10 mM ethylenediaminetetraacetic acid, 2.6 mM potassium chloride, 8.1 mM sodium phosphate, and 1.4 mM potassium dihydrogen phosphate, pH 7.2). Aliquots of 2.5 × 105 cells were blocked with mouse IgG in PBS/bovine serum albumin (BSA; 0.1%) for 15 min at 4°C, washed twice with PBS with Ca2+ and Mg2+, and resuspended in 150 μL of FACS binding buffer (cation buffer: 150 mM NaCl, 10 mM magnesium chloride, 10 mM calcium chloride, and 5 mM manganese chloride, pH 7.2, diluted to a concentration of 1:20 in PBS/0.1% BSA). The phycoerythrin-labeled murine monoclonal antibody 23C6-PE (3 μg/mL) (anti-αvβ3 complex [CD51/CD61]; Pharmingen) was added for 45 min at 4°C. Subsequently, cells were washed, resuspended in PBS/0.1% BSA, and analyzed on a Becton Dickinson FACSCalibur (BD Biosciences).
In Vitro Integrin αvβ3 Ligand-Binding Studies
Cells were seeded in 24-well plates. Confluent cells were washed twice with PBS and overlayed with 150 μL of binding buffer (cation buffer diluted to a concentration of 1:5 in 1% PBS). Afterward, 2 nmol RGD-Cy 5.5 was added into each well, incubated for 1 h at 37°C, washed twice with PBS, and covered with binding buffer. For predosing experiments, free RGD peptide (20 nmol) was added to each well for 45 min at 37°C. Vital cells were examined using a fluorescence microscope (TE 2000-S; Nikon) equipped with a mercury vapor lamp, 620/700-nm (excitation/emission) filters (AHF Analysentechnik AG), and a DXM1200F camera (Nikon).
Animal Model
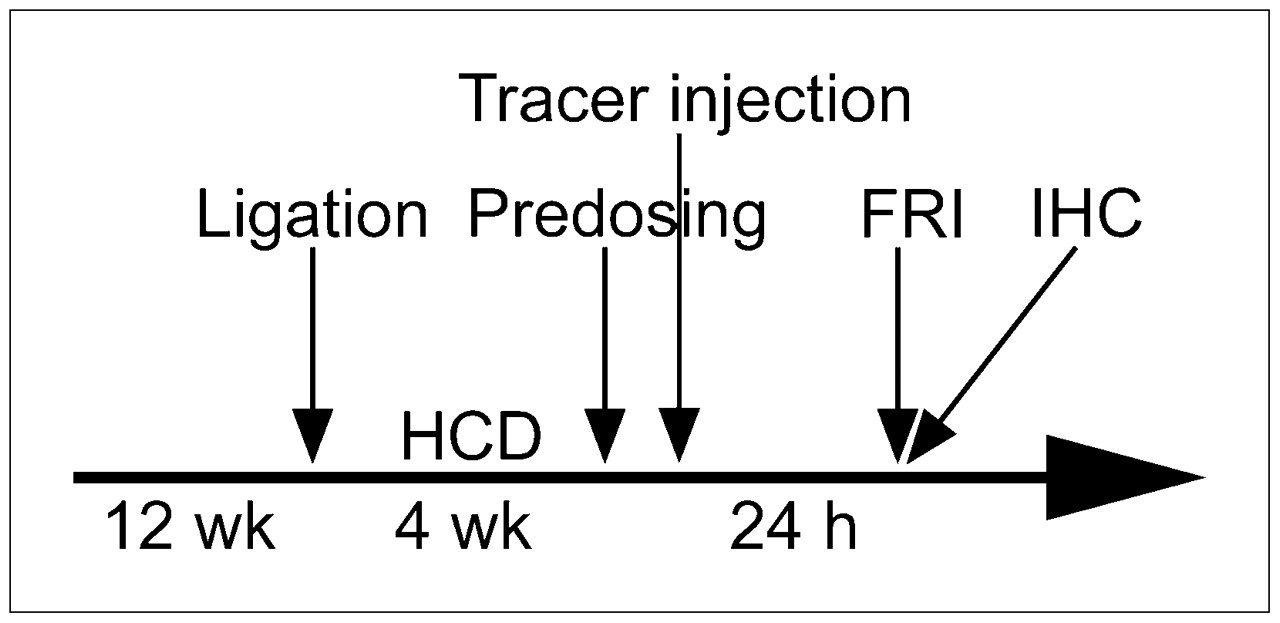
ApoE−/− mice (CL 57/BL6; age, 12 wk; weight, 20–28 g) (Charles River) were anesthetized by an intraperitoneal injection of ketamine/xylazine (125/12.5 mg per kilogram of body weight), and the left common carotid artery was ligated using 5-0 silk (Ethicon GmbH) as described elsewhere (17). The animals were then fed a high-cholesterol diet (15% cocoa fat, 1.0% cholesterol, and 0.5% sodium cholate; Altromin GmbH & Co. KG) for 4 wk (Fig. 1). For near-infrared FRI, the Cy 5.5–labeled ligand (2 nmol/animal; n = 3) was injected via the tail vein. For tracer competition studies, mice were dosed with unlabeled peptide (20 nmol; n = 3) before tracer injection. To clarify unspecific biodistribution effects, nonmodified Cy 5.5 (2 nmol; n = 3) was injected into a cohort of animals. For fluorescence imaging, animals were euthanized 24 h after tracer injection and then perfused in situ with 0.9% NaCl (w/v) for 3 min (Fig. 1). The chest was opened to expose the heart (the aortic arch and the carotid arteries). Finally, the aortic arch and the carotid arteries were dissected from the heart and surrounding fat tissue. Macroscopic images were captured in situ with a Coolpix 4500 camera (Nikon) mounted on a Nikon binocular. Fluorescence reflectance images of supraaortic vessels were acquired in situ and after resection of the aortic arch including both carotid arteries. Subsequently, the dissected tissue was embedded in Tissue-Tek O.C.T. (Sakura), snap-frozen in liquid nitrogen, and serially sectioned (10 μm). Animal use was randomized to correct for interindividual lesion heterogeneity. The protocol was approved by the Institutional Animal Care Committee.

Timeline illustrating ligation, feeding, imaging, and postimaging examinations. HCD = high cholesterol diet; IHC: immunohistochemistry.
Immunohistochemistry
Mouse-Derived Tissues.
Air-dried frozen sections were fixed in ice-cold acetone and blocked by incubation with 1% PBS/BSA for 30 min to prevent unspecific binding. Tissue sections were incubated with a monoclonal rat anti-mouse CD68 primary antibody (concentration, 1:100) (Clone FA-11; AbD Serotec) for 60 min at room temperature. A goat anti-rat Cy 3 antibody (Dianova) was applied as the secondary antibody (concentration, 1:100). Subsequently, slides were incubated with RGD-Cy 5.5 peptide for 60 min at room temperature before nuclear staining was performed using 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) (Sigma-Aldrich Chemie GmbH). Separate RGD-Cy 5.5 incubation was necessary, because fixation reduced RGD-Cy 5.5–derived fluorescence of the sections. Untreated sections exhibited RGD-Cy 5.5–derived fluorescence (data not shown). Tissue sections were captured with an Eclipse 50i microscope (Nikon) equipped with the DS 2MBWc digital camera (Nikon) and appropriate excitation/emission filters (AHF Analysentechnik AG).
Human-Derived Tissues.
With the approval of the local ethics committee, human tissues were obtained during routine carotid artery endarterectomies. Specimens were stored in RNAlater solution (Ambion) and subsequently embedded in Tissue-Tek O.C.T. (Sakura). Air-dried frozen sections (10 μm) were fixed in ice-cold acetone. Unspecific binding was blocked using 1% PBS/BSA. Tissue sections were incubated for 60 min at room temperature using a monoclonal mouse anti-human CD68 primary antibody (concentration, 1:100) (Clone KP-1; DAKO Cytomation) and a goat anti-mouse Cy 3–labeled secondary antibody (concentration, 1:100) (Dianova) for 60 min at room temperature. Subsequently, slides were processed as described above.
FRI
Near-infrared FRI was performed using the In-Vivo FX Imaging System (Kodak Molecular Imaging Systems), equipped with a 150-W halogen illuminator with Cy 5.5 bandpass excitation (625 ± 18 nm) and emission filters (700 ± 17.5 nm). Fluorescence signals were captured with a 4-million-pixel cooled charge-coupled device camera equipped with a 10× zoom lens. Images were captured 24 h after probe injection, with an acquisition time of 60 s and identical window settings (e.g., f-stop, field of view). Fluorescence images were coregistered with the anatomic white-light images using MI 4.0 software (Kodak). Equal-sized regions of interest (≥1,500 pixels for each location) were placed within the highly visible plaque-like lesion, the adjacent carotid artery that was free of lesions, and the background (i.e., beside the animal). Contrast-to-noise ratios (CNRs) were determined as follows:
Statistical Analysis
All data are presented as mean values ± SEM. Statistical analysis of CNRs was conducted using 1-way ANOVA (GraphPad Instat, version 3.06; GraphPad Software Inc.) followed by post hoc analysis (Bonferroni). A P value of less than or equal to 0.05 was considered significant.
RESULTS
Peptide Labeling
After the cyclo[Cys-Arg-Gly-Asp-Cys]-Gly-Lys (RGD) peptide was labeled with the fluorophore Cy 5.5, a purity greater than or equal to 95% was achieved by reversed-phase high-performance liquid chromatography, with a retention time of 28.5 min. Yields of the RGD-Cy 5.5 conjugate were typically 75%−90% as calculated with ε675 = 250,000 (mol/L)−1cm−1 from the absorption spectrum measured with the PBS solution. The excitation and emission spectra of the targeted probe remained unchanged, compared with the nonmodified fluorophore showing an excitation peak at 675 nm and emission maximum at 694 nm.
FACS and Microscopic In Vitro Binding Studies
Flow cytometric and fluorescence microscopic binding studies were performed to determine the capacity of the cells to synthesize the αvβ3 integrin as well as the binding affinity of the tracer molecule for the target. FACS analysis revealed high αvβ3 expression levels in RAW264.7 and M21 cells, whereas αvβ3 integrin expression was virtually nonexistent in MCF-7 cells (data not shown). These results were confirmed by the cell assay in vitro. Cy 5.5–labeled RGD peptide distinctly bound to integrin-positive RAW and M21 cells (Supplemental Fig. 1; supplemental materials are available online only at http://jnm.snmjournals.org), whereas αvβ3-negative MCF-7 cells revealed no detectable level of cell-associated fluorescence. Incubation in advance with a 10-fold excess of the unlabeled peptide (140 μM) drastically reduced binding of the labeled tracer, proving specificity of the probe (Supplemental Fig. 1). Negligible signals were detected for all cell lines when they were incubated with the nonmodified Cy 5.5 dye (up to 140 μM; data not shown).
Near-Infrared FRI
Carotid artery ligation resulted in the formation of macrophage-rich vascular lesions proximal to the ligation in ApoE−/− mice (n = 9). RGD-Cy 5.5 strongly accumulated in the lesions (Fig. 2). RGD-Cy 5.5–derived signals were significantly stronger than with nonmodified Cy 5.5 and predosing experiments as visualized by high-resolution ex vivo FRI (Fig. 3). Autofluorescence of lesions was in the same range as background fluorescence derived from other tissues (data not shown). Thus, target-specific tracer binding can be assumed. Unspecific biodistribution effects of the unmodified Cy 5.5 seem, on the contrary, insignificant.
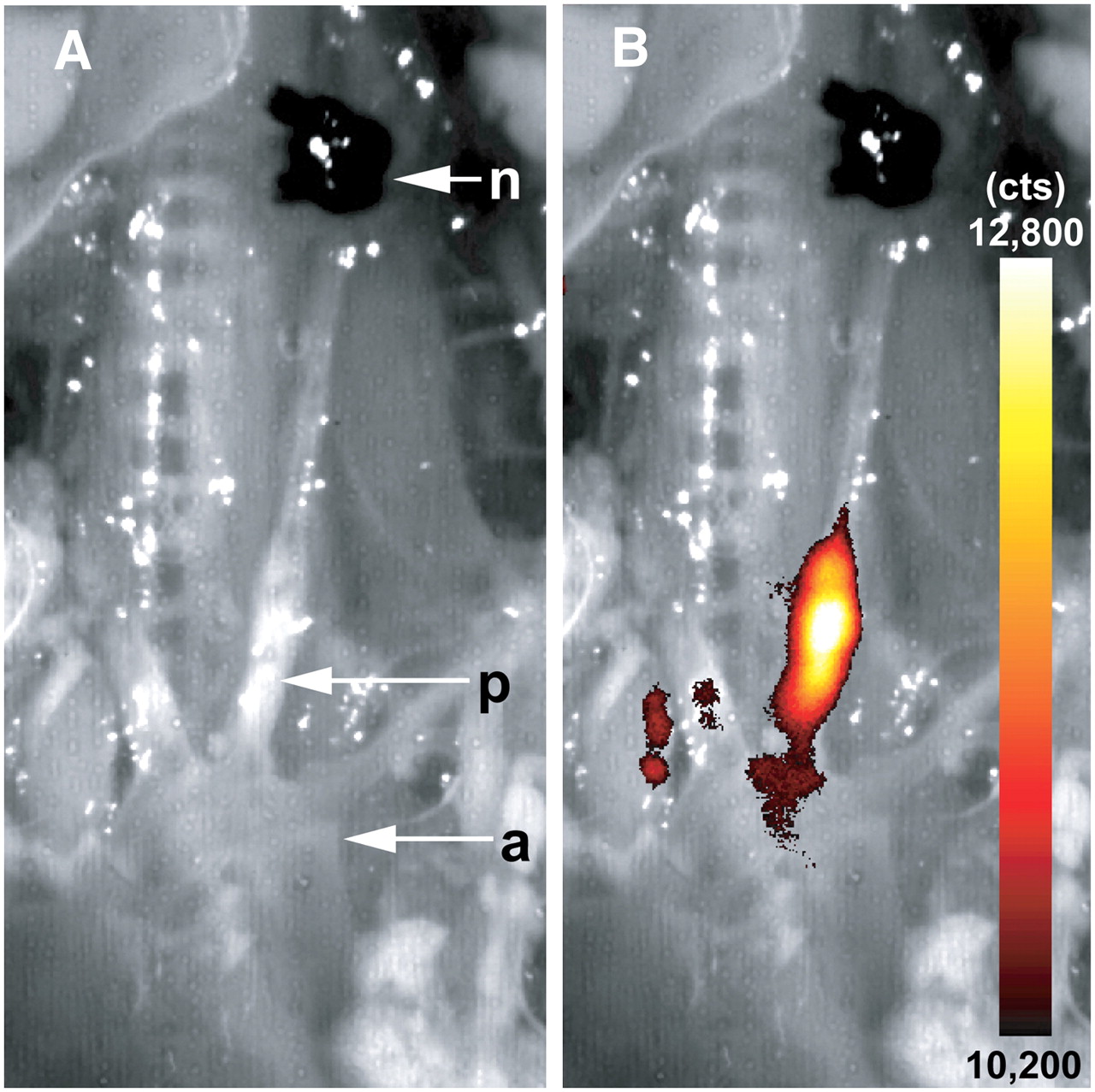
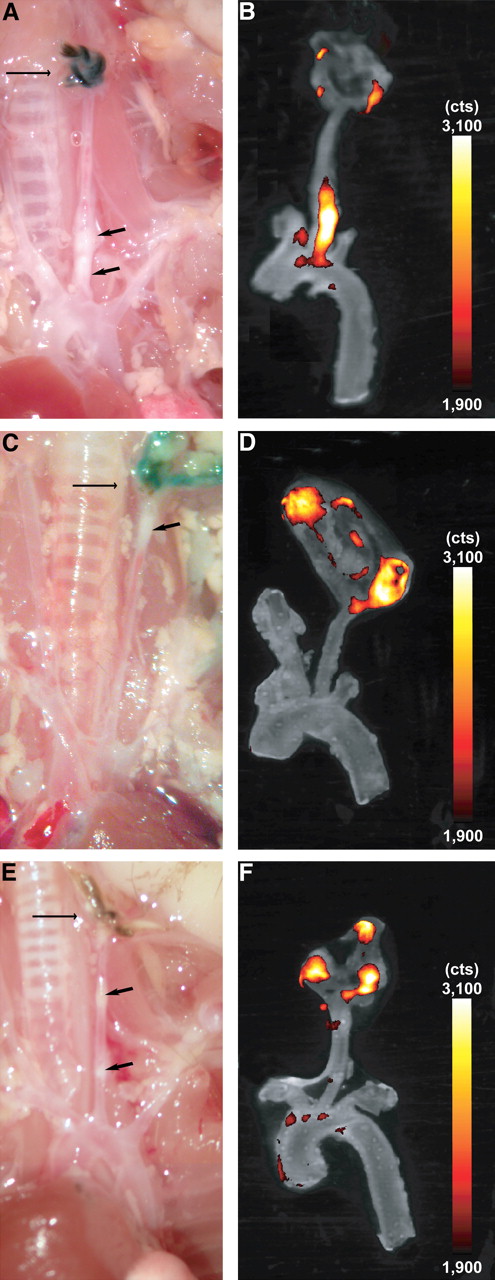
In situ fluorescence reflectance imaging overview of supraaortic vessels. FRI data were evaluated 24 h after intravenous injection of RGD-Cy 5.5 probe. Lesion formation (p) is clearly visible in bright field anatomic image (A). Fluorescence signals, given as photon counts per second (cts), emerged in area of lesion formation (B; merge of A with FRI data) proximal of ligation node (n). a = aortic arch.
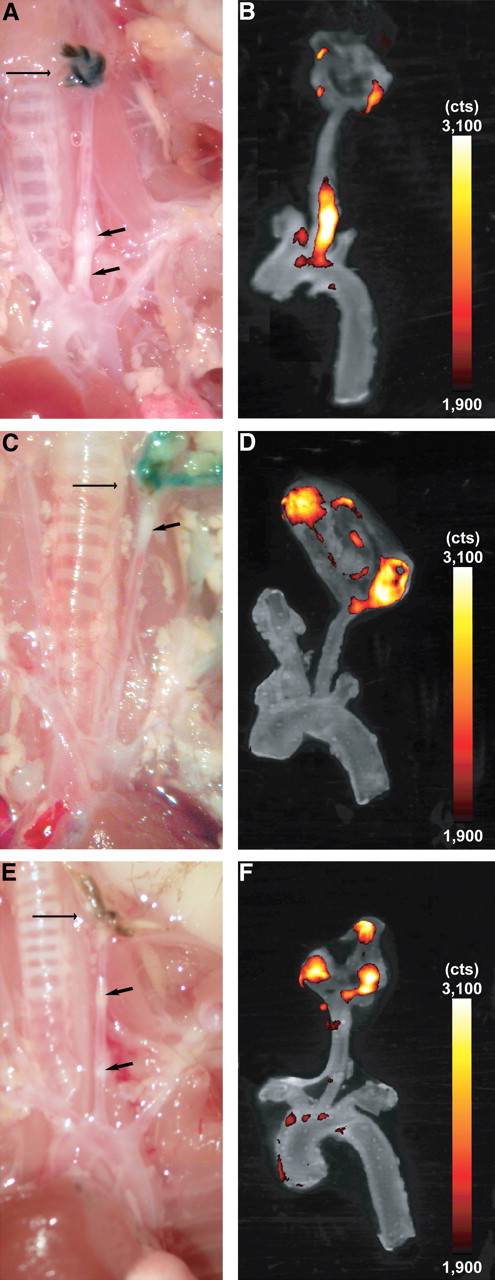
FRI of resected carotid artery in different experimental groups. ApoE−/− mice were injected with either RGD-Cy 5.5 (A and B), free RGD peptide before tracer injection (C and D), or free Cy 5.5 (E and F). Arrows on macroscopic images (A, C, and E) indicate vascular lesions (short arrow) and ligation (long arrow). (B, D, and F) Overlay of color-encoded near-infrared fluorescence images of resected arteries and corresponding white-light images. Fluorescence intensity is given in photon counts per second (cts). Strong fluorescence signal was observed at plaques of left carotid artery using Cy 5.5–labeled RGD peptide (B), which corresponds well with visible atherosclerotic lesion (A). Dosing in advance reveals artifical fluorescence signal at site of surgery but no relevant fluorescence within plaque (D). Free Cy 5.5 resulted in weak signal in plaque (F). For CNR calculation, images were processed and windowed identically.
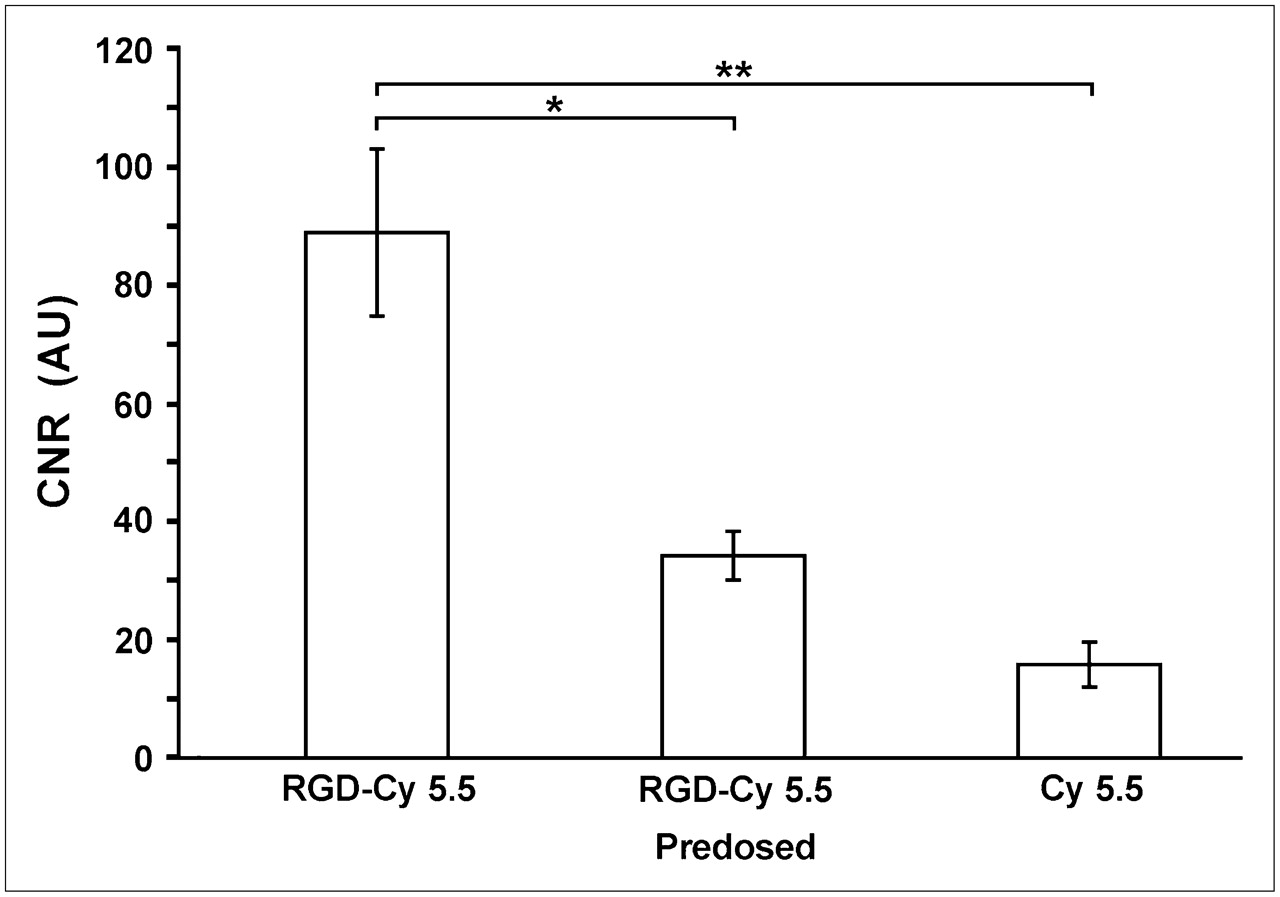
Semiquantitative assessment demonstrated the highest CNRs in RGD-Cy 5.5–injected mice (CNR, 88.8 ± 14.1, n = 3; Fig. 4), whereas predosing experiments (n = 3) efficiently decreased the CNR (34.2 ± 4.1). Free Cy 5.5 dye resulted in less than 20% of the RGD-Cy 5.5–mediated CNR (CNR, 15.8 ± 3.8).

Semiquantitative FRI results for all experimental groups. Calculated CNRs were 88.8 (±14.1 [SEM]) for RGD-Cy 5.5–injected animals, which could be significantly reduced by advance dosing with free peptide (CNR, 34.2 ± 4.1). Free Cy 5.5 revealed only weak fluorescence signals (CNR, 15.8 ± 3.8). *P < 0.05; **P < 0.01. AU = arbitrary units.
Immunohistochemistry
To determine whether RGD-Cy 5.5 signals in vascular lesions were associated with macrophage infiltration, immunohistochemical stainings for the macrophage-specific antigen CD68 were performed (Supplemental Fig. 2). The RGD peptide–associated signal in the neointima colocalized with the macrophage-associated CD68 staining to a significant extent (Supplemental Fig. 2B). As expected, RGD-Cy 5.5 also stained the vascular SMCs and elastin-rich fibers (Supplemental Fig. 2D). Moreover, double stainings of human atherosclerotic plaques revealed distinct colocalization of αvβ3 and CD68 (Fig. 5D).
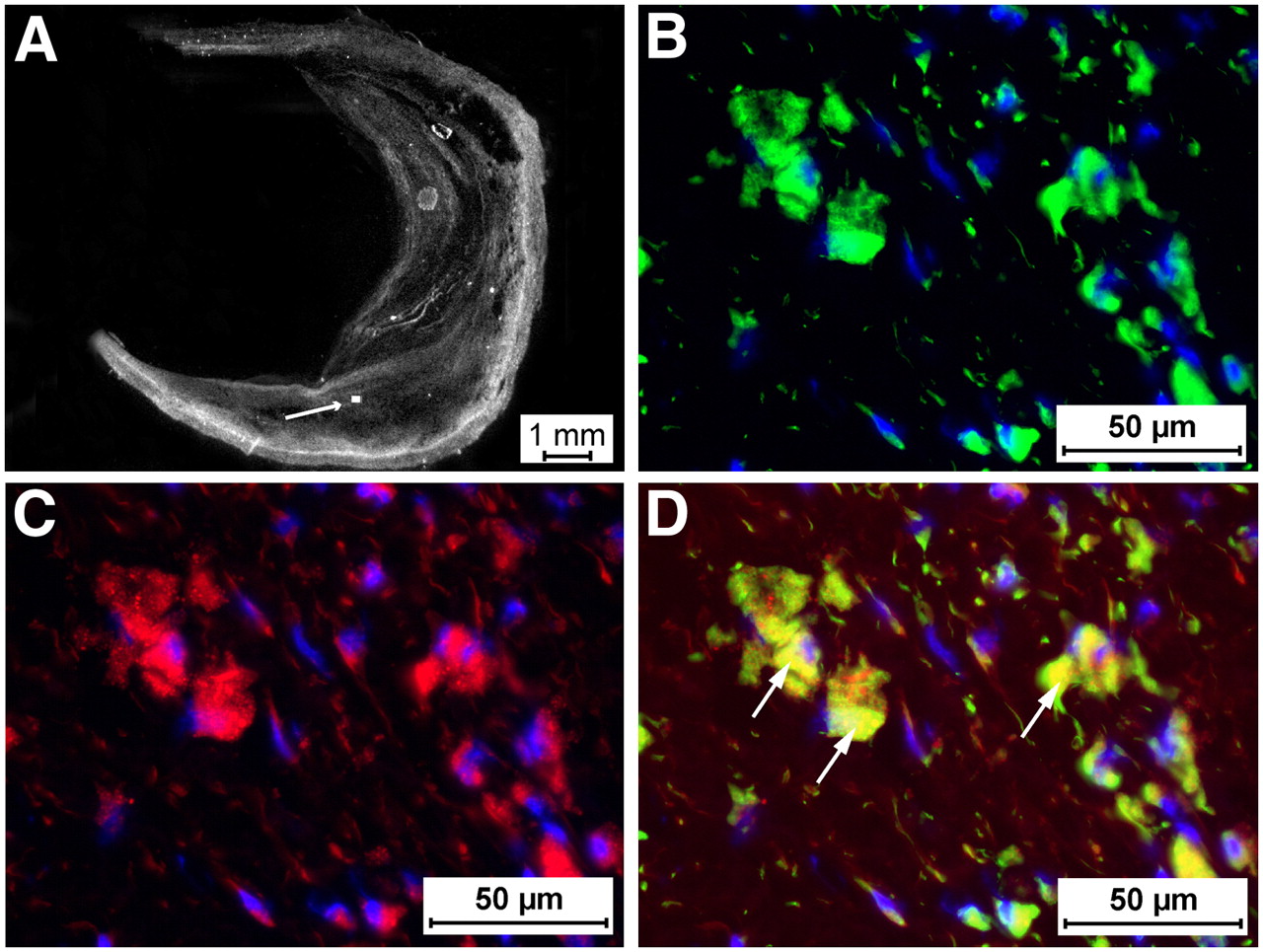
Immunohistochemistry of human atherosclerotic plaques. Human carotid endarterectomy specimens (A, white light) were stained for CD68 (B and D; green signal) and RGD-Cy 5.5 (C and D; red signal). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; B, C, and D; blue signal). (D) Merged image of macrophages (arrows) within plaque demonstrated CD68 and RGD-Cy 5.5 double staining, suggesting that macrophage infiltration may potentially be imaged in humans using this approach. Box and arrow indicate the enlargement shown in B–D. Scale bars are indicated.
DISCUSSION
Progression and destabilization of atherosclerotic plaques with consecutive rupture is a critical process potentially resulting in life-threatening cardiovascular events such as myocardial infarction or stroke. Cellular processes such as monocyte-derived macrophage infiltration and enhanced MMP activation drive plaque instability (4,18). Although numerous studies explained the potential of targeting αvβ3 to image myocardial angiogenesis (e.g., of the healing myocardium after infarction (19)), we hypothesized that αvβ3 may be a suitable target to visualize atherosclerotic plaques because it is highly expressed on macrophages (9) and binds MMP-2 (15), a key molecule for plaque destabilization. Fluorochrome labeling of RGD peptides (12,13,20) and tumor targeting with optical probes using planar reflectance and 3-dimensional tomographic imaging methods have been shown to be feasible (16,21). The RGD tripeptid sequence is an integrin-shared adhesive motif (22) with distinct different affinities to Arg-Gly-Asp–dependent integrins such as αvβ3, α5β1, or αIIbβ3 and led to the concept that different integrins distinguish varieties in the conformation and sequential environment of various RGD sites (13,23–25). Cyclization of the RGD peptide also increased binding with vitronectin receptors (i.e., αvβ3) dramatically but had no effect on the affinity to, for example, fibronectin receptors such as α5β1 (13,23,26,27). In this study, a Cy 5.5–labeled cyclic RGD peptide specifically binding αvβ3 integrins (20) was used to detect resident macrophages in vascular lesions in ApoE−/− mice. Because cyanine dyes are in part Food and Drug Administration approved (e.g., indocyanine green) or successfully applied in clinical studies, which is also true for RGD peptides, the toxicity of the construct for clinical applications should be negligible. Furthermore, cyclic RGD peptides are known to possess inhibitory concentrations of 50% ranging between 7 and 40 nmol/L for the inhibition of vitronectin to immobilized αvβ3 (28). A cystin moiety for cyclization was chosen because of the reported enhanced targeting efficiency of such peptides (29,30). In vitro, the probe tightly bound to viable murine RAW264.7 macrophages and αvβ3 integrin–positive M21 melanoma cells (31,32), suggesting a high αvβ3 receptor expression in macrophages. Binding of the RGD peptide to RAW macrophages was significantly reduced by dosing with the free cyclic peptide in advance, supporting target specificity of the probe.
On the basis of these data, RGD-Cy 5.5 was used for near-infrared FRI of macrophage-rich vascular lesions in the ligated left common carotid artery of ApoE−/− mice. This model differs from others in that it does not require mechanical trauma and widespread endothelial denudation to induce SMC proliferation. Notwithstanding, the carotid artery ligation model cannot exactly mimic the human pathophysiology. However, it emulates certain aspects of human vascular lesion formation, that is, macrophage and protease accumulation (33). Our results highlighted the capability of the Cy 5.5–labeled RGD peptide to specifically image αvβ3 expression in those vascular lesions because the fluorescence signal could be efficiently blocked in competition experiments. Nontargeted cyanine dyes did not show any relevant fluorochrome retention in the lesions. In addition, fluorescence signals were detected at the site of surgery (Fig. 3) distal to the arterial ligation. This may well reflect postsurgical inflammatory processes. However, the CNRs of the RGD-Cy 5.5 signals within the lesion were unambiguously higher than CNRs from tracer competition experiments or when unlabeled cyanine dye was used. Thus, simple nonspecific biodistribution effects of the cyanine dye can be excluded. Immunohistochemistry confirmed that FRI signals were due to accumulation of the tracer in αvβ3-expressing macrophages. We used a specific antibody against the 110-kDa transmembrane glycoprotein CD68, specific for monocytes and tissue macrophages (34), and restricted staining to macrophages within the neointima. RGD-Cy 5.5 not only strongly stained macrophages but also revealed a weaker signal associated with elastin-rich fibers of the medium and vascular SMCs underlying the atherosclerotic plaque. These results were also congruent to the findings of Antonov et al. (9), who demonstrated that αvβ3 integrin receptors, compared with endothelial cells, SMCs, and tissue macrophages, were highly upregulated in atheroma macrophages of early lipid (fatty streak) and advanced atherosclerotic lesions. However, endothelial cells and SMCs are known to express the vitronectin receptor on their surface (11) and to bind proteolytically active MMP-2, among other ligands. Assuming a migratory phenotype (15), RGD-Cy 5.5 binding to these cells is expected and may potentially dilute the signal-to-noise ratio of the macrophages but may actually add to the sensitivity of lesion detection in vascular imaging approaches. Immunohistochemistry on the human carotid endarterectomy specimen confirmed high expression levels of the target in human atherosclerotic plaques and colocalization with CD68-positive macrophages. Although other cell types and extracellular matrix components did exhibit some binding of the probe, the macrophage-bound RGD-Cy 5.5 signal was much stronger and readily distinguishable, suggesting that αvβ3 targeting may be a viable concept for translation from bench to bedside.
Furthermore, this tracer might be applicable in angiogenesis imaging for risk stratification of patients after myocardial infarction, because αvβ3 is barely detectable in endothelial cells in the quiescent state but shows high expression in vascular endothelial cells during angiogenesis (35,36).
In general, optical imaging is an attractive concept for the detection of molecular signatures in vivo because of its exquisite sensitivity, which is comparable to scintigraphic imaging techniques (37). In the near-infrared range, good tissue penetration and low background signal allow fluorochromes to be detected, even in the pico- to femtomolar range. Potential clinical applications of this optical imaging technology for cardiovascular diseases are conceivable. More recently, imaging techniques such as optical coherence tomography have successfully been explored clinically for characterization of coronary artery plaques using a catheter-based imaging approach (38). The RGD-Cy 5.5 tracer tested in this work might be usable in combination with optical coherence tomography to identify plaque rupture, fibrous cap erosion, intracoronary thrombi, and thin cap fibroatheroma, because with planar fluorescence imaging techniques such as FRI a sufficient correlation between fluorescence signal and anatomic fluorescence origin is hard to establish. Thus, thorax opening was necessary in our approach. However, optical coherence tomography offers exquisite anatomic resolution in the micromolar range and might allow in vivo microscopic resolution of structural signatures related to plaque destabilization. This technique may provide the chance to specifically determine plaque dignity significantly earlier than with the available technologies, which are capable of imaging only plaque morphology at best. From a technical point of view, near-infrared imaging technology can be miniaturized just like optical coherence tomography so that a clinical translation could be straightforward. Indeed, quantitative real-time catheter-based fluorescence imaging has most recently been described for laparoscopic and colonoscopic applications in mice so that the grounds for translation of this technology for intravascular applications are prepared (39). Thus, near-infrared imaging combined with a target-specific probe as shown in this study may potentially further enhance the diagnostic potential of invasive coronary angiography. Moreover, incorporation of FRI techniques into ultrasoundlike, handheld devices has been proposed, opening avenues to apply near-infrared imaging for characterization of superficial artery segments (e.g., carotid artery) or intraprocedural applications of the technology (40).
CONCLUSION
This study demonstrates that macrophage-associated αvβ3 integrin expression can be visualized using FRI approaches. This imaging technique is, therefore, a versatile tool not only for experimental small-animal molecular imaging but also potentially for noninvasive target detection, for example, for clinical plaque imaging in future imaging applications.
Acknowledgments
We thank Susanne Greese, Wiebke Gottschlich, Dr. Julia Heselhaus, Irmgard Hoppe, and Ingrid Otto-Valk for their technical assistance and Bodo Levkau for providing the macrophage cell line. This work was supported in part by the Deutsche Forschungsgesellschaft (BR 1653/2-1, SFB 656 project A1, A4, Z2), Interdisciplinary Center for Clinical Research (FG3, The1/068/04, ZPG 4b), and European Community (IP 6th framework, Molecular Imaging and Network of Excellence DiMI).
Footnotes
-
↵* Contributed equally to this work.
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication March 8, 2008.
- Accepted for publication July 2, 2008.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- 64Cu-Labeled Divalent Cystine Knot Peptide for Imaging Carotid Atherosclerotic Plaques
- In Vivo Evaluation of Angiogenic Activity and Its Correlation with Efficacy of Indirect Revascularization Surgery in Pediatric Moyamoya Disease
- PET/CT Imaging of Integrin {alpha}v{beta}3 Expression in Human Carotid Atherosclerosis
- Claudin-4-targeted optical imaging detects pancreatic cancer and its precursor lesions
- Integrin-Targeted Imaging of Inflammation in Vascular Remodeling
- Evaluation of {alpha}v{beta}3 Integrin-Targeted Positron Emission Tomography Tracer 18F-Galacto-RGD for Imaging of Vascular Inflammation in Atherosclerotic Mice
- Optical and Multimodality Molecular Imaging: Insights Into Atherosclerosis