


Abstract
Optical techniques, such as bioluminescence and fluorescence, are emerging as powerful new modalities for molecular imaging in disease and therapy. Combining innovative molecular biology and chemistry, researchers have developed optical methods for imaging a variety of cellular and molecular processes in vivo, including protein interactions, protein degradation, and protease activity. Whereas optical imaging has been used primarily for research in small-animal models, there are several areas in which optical molecular imaging will translate to clinical medicine. In this review, we summarize recent advances in optical techniques for molecular imaging and the potential impact for clinical medicine.
Recent advances in genomics and proteomics have identified a large number of molecules and signaling pathways that potentially promote or limit diseases such as cancer, atherosclerosis, and infectious disease. A key challenge for basic scientists and clinicians is defining which pathways, among many, control disease initiation and progression in intact animals or patients, rather than in reductionist systems. Ongoing discoveries of potential target molecules in disease also drive pharmaceutical and biotechnology companies to find leading agents that selectively regulate these molecular pathways in vitro and in vivo. Optical techniques, including bioluminescence and fluorescence, are emerging as key technologies to meet these challenges and advance molecular imaging in preclinical research and patient care. This review will discuss basic methods in optical imaging for preclinical and clinical applications and highlight recent advances for in vivo imaging of specific molecular processes and signaling pathways.
OPTICAL IMAGING TECHNOLOGIES FOR PRECLINICAL AND CLINICAL APPLICATIONS
The focus on optical imaging techniques for molecular imaging is driven in large part by the sensitivity for imaging optical contrast agents and reporter molecules in vivo. The lower limits of detection for optical imaging may reach picomolar or even femtomolar concentrations of an optical reporter or contrast agent. Combined with the minimal background of techniques such as bioluminescence imaging and fluorescence imaging in the near-infrared spectrum, the signal-to-background ratio for detecting specific molecular signals equals or exceeds that which can be achieved with other molecular imaging modalities.
A key challenge for optical imaging probes and instrumentation, particularly those targeted toward eventual clinical applications, is overcoming attenuation and scattering of light by tissues. For light in the visible spectrum, absorption by hemoglobin and other molecules may reduce optical signals by approximately 10-fold per centimeter of tissue (1). To image fluorescence in deeper tissues, investigators have developed strategies for imaging near-infrared fluorescence (NIRF) with emission wavelengths between 650 and 900 nm. At these wavelengths, absorption of light by hemoglobin, lipids, and water is lowest, and tissue autofluorescence also is greatly reduced. As a result, the sensitivity for NIRF imaging agents is greatly enhanced, potentially allowing for tomographic optical imaging signals to be detected at depths of 7–14 cm (2).
Differential absorption of light by tissues also produces images that are weighted toward optical reporters and probes that are located closer to the surface of a subject. While this limitation is being overcome with 3-dimensional imaging and analysis techniques such as fluorescence molecular tomography (FMT) (3), optical techniques typically allow relative quantification of imaging signals, rather than absolute quantification possible with PET. Despite these challenges, optical techniques have growing roles in molecular imaging research and clinical translation.
Bioluminescence Imaging
Bioluminescence imaging is commonly used for preclinical cellular and molecular imaging in small animals. Bioluminescence refers to light produced by the enzymatic reaction of a luciferase enzyme with its substrate (Fig. 1). Firefly (Photinus pyralis) luciferase is the most frequently used luciferase for molecular imaging. This enzyme oxidizes its substrate, luciferin, in a reaction that requires oxygen and adenosine triphosphate (ATP), emitting light with a broad emission spectrum and a peak at ≈560 nm. Because of tissue attenuation, red and far-red emissions from firefly luciferase are detected preferentially for imaging in small animals. Luciferin distributes throughout an animal rapidly after intraperitoneal injection and passes across blood–tissue barriers including the brain and placenta. Light from firefly luciferase peaks ≈10–12 min after injection of luciferin and decreases slowly over 60 min (4), providing a broad time window for acquiring images. The combination of enzymatic amplification of signals from luciferase and the almost negligible background bioluminescence in vivo makes bioluminescence imaging with firefly luciferase a highly sensitive method for small-animal molecular imaging.
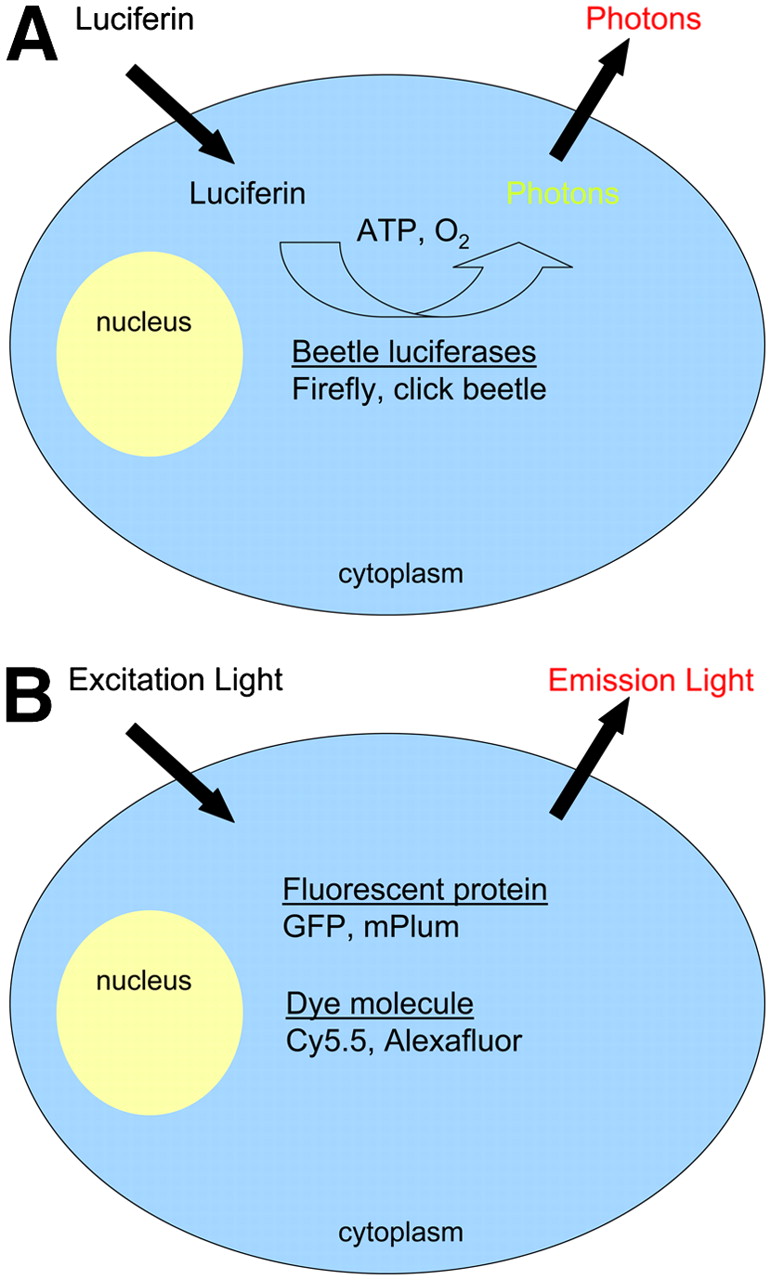
Schematic representation of bioluminescence and fluorescence imaging reporters. (A) Beetle luciferase enzymes (firefly, click beetle) expressed in cytoplasm of engineered cells catalyze production of light photons from the substrate luciferin in the presence of oxygen and ATP. Gaussia and Renilla luciferases use a different substrate, coelenterazine, and are independent of ATP. In its native form, Gaussia luciferase is secreted from cells and reacts with substrate in the extracellular space. (B) Fluorescence imaging requires light of appropriate wavelength to excite the reporter molecule (fluorescent protein or dye molecule), resulting in emission of light with a defined emission spectrum for each reporter protein or dye.
There are a variety of luciferase enzymes from other organisms that possess unique spectral characteristics and substrate requirements. Luciferases from a click beetle, Pyrophorus plagiophthalamus, have been optimized to produce green–orange (544 nm) or red (611 nm) light after oxidizing luciferin. Despite the potential advantages of 611-nm emission for in vivo imaging, firefly luciferase remains the preferred enzyme for bioluminescence molecular imaging because its reaction with luciferin produces more light. Renilla (sea pansy) and Gaussia (marine copepod) luciferases react with a different substrate, coelenterazine, to produce blue light with peak emission at approximately 480 nm. These enzymes are ATP-independent and are smaller than firefly luciferase, which is advantageous for producing genetic fusions with other proteins (5). The blue emission wavelength of these enzymes, limited biodistribution and rapid kinetics of coelenterazine in small animals, and higher background have restricted in vivo applications for molecular imaging (6–8). Generating red-shifted variants of these marine luciferases, such as has been accomplished recently for Renilla luciferase, will greatly improve their uses in small-animal imaging (9).
Lux operons from bacteria, such as Photorhabdus luminescens, emit blue light that has been used to image bacterial pathogenesis. The lux operon encodes all proteins required for bioluminescence, so bacteria engineered to express the lux operon do not require an exogenous substrate to produce light. Unfortunately, attempts to incorporate this suite of genes into mammalian cells have been unsuccessful. The availability of multiple luciferase enzymes allows 2 different biologic processes to be monitored in vivo, using appropriate spectral filters and substrates (10). Although bioluminescence imaging is unlikely to be used in humans, this technique is a powerful tool to interrogate mechanisms of disease and accelerate drug development in preclinical models.
Fluorescence Imaging
In fluorescence imaging, an external light of appropriate wavelength is used to excite a target fluorescent molecule, followed almost immediately by release of longer-wavelength, lower-energy light for imaging. Targets for fluorescence imaging may be endogenous molecules (such as collagen or hemoglobin), fluorescent proteins (green fluorescent protein [GFP] and related molecules), or optical contrast agents with fluorescent molecules. Although each of these sources of fluorescence can be used for cellular and molecular imaging in small-animal models, optical contrast agents have the potential to extend from preclinical research to patient care.
In vivo fluorescence imaging encompasses a wide range of resolution and imaging depths, including subcellular analysis at <400 μm with intravital microscopy and whole-animal imaging with 1- to 3-mm spatial resolution at <10 cm with FMT (11). In addition, fiber-optic endoscopes with confocal or 2-photon laser fluorescence imaging will facilitate clinical use of fluorescence molecular imaging with targeted optical contrast agents (12). Manufacturers also have developed filter sets and algorithms for unmixing spectra of several different fluorescent signals, resulting in greater sensitivity for detecting multiple fluorescent reporters in vivo (13).
Fluorescence imaging at depths greater than a few millimeters requires NIRF probes and dyes. There is an ever-growing list of fluorophores for NIRF imaging in small animals, including Cy5.5, the Alexa dye series, indocyanine green, and quantum dots. Whereas potential toxicity may limit applications of Cy5.5 or quantum dots in patients, gaining Food and Drug Administration approval for clinical use of a NIRF dye will be a key step toward using fluorescence imaging for routine clinical applications. In addition, there are ongoing challenges to develop contrast agents targeted to specific molecules, especially agents in which fluorescence is quenched until released by a specific enzyme or defined target.
Quantification of Optical Imaging Data
Optical imaging data typically are quantified by region-of-interest (ROI) analysis, measuring photon flux for bioluminescence and absorption units or efficiency for fluorescence. Quantification is complicated by variable attenuation of light in different organs and tissues. For example, less light will be detected from the same number of molecules of luciferase in the liver as compared with the lung (14). For 2-dimensional images, optical signals within a given ROI may represent the summation of overlapping anatomic structures. Tomographic optical imaging techniques, such as FMT, partially overcome these limitations and improve the accuracy of data analysis (11). Although current optical imaging technologies provide relative quantification of signals, a key strength of optical imaging for preclinical studies is the ability to measure inhibition and dissociation constants in vivo (15–17). As a result, both bioluminescence and fluorescence imaging have been used to assay pharmacodynamics of therapeutic agents in vivo, establishing dosing parameters to guide initial clinical trials and eventual clinical applications.
Imaging Signaling Pathways and Molecules
Strategies based on protein-assisted complementation of luciferases have proven to be particularly versatile for imaging protein interactions and signaling pathways. Each luciferase enzyme can be divided into 2 inactive or minimally active fragments that do not spontaneously associate (16,18–20). When these separate fragments of luciferase are fused to interacting proteins, association between the proteins of interest brings luciferase fragments into close proximity, reconstituting enzymatic activity. Therefore, bioluminescence can be used to monitor association and dissociation of 2 proteins during intracellular signaling or in response to therapy.
Renilla, firefly, and Gaussia luciferase complementation has been used to detect constitutive, reversible, or drug-mediated protein association and dissociation in cells and mice (16,18,21–23). The ability to monitor specific protein interactions and their disruption in response to therapeutic compounds provides a facile system to screen for inhibitors of protein interactions, which is a current focus of drug development.
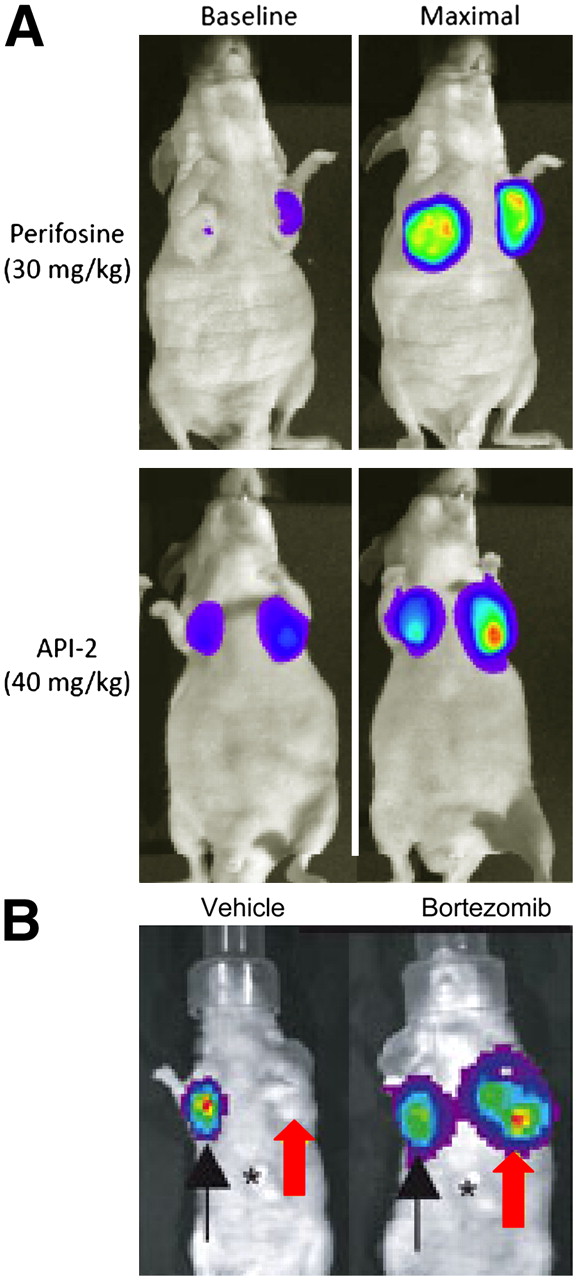
Luciferase complementation also has been used to produce novel sensors of biologic pathways. Paulmurugan and Gambhir used Renilla luciferase complementation to develop a sensor for estrogen signaling and effects of agonists, antagonists, and partial agonists of the estrogen receptor (24). Rehemtulla and coworkers developed a firefly luciferase complementation reporter for kinase activity of the oncogene AKT, which could quantify activation and pharmacologic inhibition of this kinase in cultured cells and mice (Fig. 2A) (17). These reporters for key signaling pathways promise to accelerate drug discovery and development, which will improve therapy for cancer and other diseases.

Imaging reporters for protein function and stability. (A) In vivo detection of AKT kinase activity. Mice were implanted with cells stably expressing an imaging reporter that is inhibited by AKT kinase activity. Under baseline conditions, tumors show low bioluminescence. Bioluminescence increases markedly in mice treated with perifosine or API-2, inhibitors of AKT kinase activity. (B) Imaging proteasome function. HeLa cells stably expressing control firefly luciferase (black arrow), the proteasome luciferase reporter (red arrow), or empty vector with no luciferase (asterisk) were implanted into nude mice. Bioluminescence from proteasome reporter was undetectable in mice treated with vehicle control, whereas inhibiting proteasome with the chemotherapeutic drug bortezomib stabilized the reporter and produced bioluminescence comparable to firefly luciferase alone.
Imaging Protein Stability
Degradation of proteins mediated by the ubiquitin–proteasome system regulates key cellular processes, including cell cycle progression, inflammation, and neurodegenerative diseases, and the proteasome is a promising target for cancer therapy. Recent studies show the power of molecular imaging to analyze the proteasome and stability of target proteins. Luker et al. generated a bioluminescent reporter that allowed in vivo monitoring of proteasome function and inhibition in response to pharmacologic agents (Fig. 2B) (15). Notably, this reporter revealed differences in proteasome function in response to acute versus chronic inhibition with the chemotherapeutic drug bortezomib.
Other studies have imaged the stability of proteins that are degraded by the proteasome, using reporters in which a protein of interest is fused to firefly luciferase. Kaelin and coworkers generated reporters for stability of p27, a negative regulator of the mammalian cell cycle, and Hif-1α, a transcription factor that is stabilized under hypoxic conditions (25,26). Similarly, Gross et al. produced a luciferase reporter to image stability of IκBα, a negative regulator of transcription factor NF-κB (27). For each reporter molecule, the level and stability of the reporter protein paralleled those of the respective endogenous proteins in cultured cells and mouse models, allowing effects of physiologic stimuli and therapeutic agents to be monitored with bioluminescence imaging. In addition to studies in transfected cells and tumor xenografts, Kaelin's group also made a genetically-engineered mouse expressing the Hif-1α-luciferase reporter and used this animal to image unique effects of normoxia and hypoxia on the kidneys. These studies emphasize the power of molecular imaging to monitor levels of specific proteins in vivo.
In Vivo Detection of Protease Activity
Because proteases are central to diseases, including cancer and atherosclerosis, there is intense interest in imaging protease activity. Weissleder and coworkers have developed and validated a large number of fluorescent NIRF probes for intracellular and extracellular proteases, such as cathepsins, matrix metalloproteinases, HIV protease, and urokinase plasminogen activator (28). In general, these protease sensors are designed as quenched NIRF probes that contain consensus recognition and cleavage sites for the protease of interest. The intact, quenched probe has a very low level of fluorescence, but protease activity cleaves the imaging substrate and produces substantially increased fluorescence for imaging. These fluorescent probes are powerful tools for spatial and temporal analyses of molecular mechanisms of disease in vivo. For example, Grimm et al. used microarray analysis to determine that cathepsin cysteine proteases are upregulated in a mouse model of lung adenocarcinoma (29). Using a fluorescent probe for cathepsins and 3-dimensional imaging with FMT, these researchers were able to identify tumors as small as 1 mm in vivo and coregister the fluorescent signals with tumor masses identified by microCT.
Jiang et al. developed protease reporter molecules based on a fluorophore (Cy5) linked to a polycationic cell-permeant peptide, specific protease cleavage site, and polyanionic peptide (30). Protease activity at the surface of tumor cells cleaves the reporter peptide and allows the permeant peptide to carry the fluorophore into adjacent cells. This strategy allowed in vivo detection of matrix metalloproteinase-2 activity in tumor xenografts, although the 2-fold contrast enhancement is modest. Though this appears to be a general method for imaging protease activity, further optimization is needed to enhance the imaging signal.
Strategies to Improve Detection of Tumor Margins and Metastatic Foci
Another goal in molecular imaging is to improve detection of small foci of primary or metastatic cancer and distinguish boundaries between malignant and normal tissue. Kobayashi and colleagues developed a 2-step activation strategy for sensitive, specific detection of peritoneal metastases in mouse models (31). Tumors initially were pretargeted with a biotinylated antibody to epidermal growth factor receptor. After allowing the primary antibody to clear, the pretargeted antibody was revealed with a neutravidin-conjugated fluorescent secondary antibody, allowing detection of submillimeter tumor foci in the peritoneal cavity. Although not tested formally, the pretargeting strategy with activated fluorescence should be a general platform for fluorescence molecular imaging of a wide variety of cell-surface molecules.
A complementary strategy for delineating malignant cells has been developed for glioma, a cancer in which tumors commonly recur at the surgical margin. Veiseh et al. synthesized a bioconjugate of chlorotoxin and the NIRF dye Cy5.5 that preferentially binds to glioma cells in a process dependent on matrix metalloproteinase-2 (32). When injected intravenously, this agent crossed the blood–brain barrier and bound to malignant, but not normal, cells in the brain. Tumor foci composed of a few hundred cells were detectable in vivo. Because chlorotoxin has been administered safely to patients with glioma, this optical imaging agent appears to have great potential for clinical translation.
CONCLUSION
Optical molecular imaging techniques have become essential tools for studying small-animal models, providing unique insights into disease pathogenesis, drug development, and effects of therapy. In this way, optical imaging already is making a substantial impact on basic and translation medical research (33). Fluorescence imaging is entering initial clinical testing in areas such as breast imaging and endoscopy. For example, diffuse optical spectroscopy of hemoglobin and deoxyhemoglobin in breast tumors shows promise as a biomarker for effective neoadjuvant chemotherapy in cancer patients (34). As molecular contrast agents translate from preclinical studies to patients, potential applications of optical molecular imaging in clinical care are expected to expand dramatically. Although there will continue to be challenges with interrogating fluorescence in deep tissues, diagnostic tests in which the detection instrument can be positioned near the fluorescent probe—such as endoscopy, intraoperative scanning, and breast imaging—are likely to flourish. Therefore, optical fluorescence imaging will be a key part of advancing molecular imaging into direct applications in patients.
Acknowledgments
The authors thank Limin Zhang and Al Rehemtulla for Figure 2A. Work in the authors' laboratory is supported in part by grant BCTR0503550 from the Susan Komen Foundation, a grant from the Sidney Kimmel Foundation, and NIH R24CA083099 grant for the University of Michigan Small Animal Imaging Resource.
Footnotes
-
COPYRIGHT © 2008 by the Society of Nuclear Medicine, Inc.
References
- Received for publication August 1, 2007.
- Accepted for publication October 22, 2007.





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- In vitro and in vivo NIR Fluorescence Lifetime Imaging with a time-gated SPAD camera
- Aggregation-induced emission dots assisted non-invasive fluorescence hysterography in near-infrared IIb window
- Tumor-Specific Uptake of Fluorescent Bevacizumab-IRDye800CW Microdosing in Patients with Primary Breast Cancer: A Phase I Feasibility Study
- Detection of Micrometastases Using SPECT/Fluorescence Dual-Modality Imaging in a CEA-Expressing Tumor Model
- Targeted Dual-Modality Imaging in Renal Cell Carcinoma: An Ex Vivo Kidney Perfusion Study
- Targeting Acidity in Pancreatic Adenocarcinoma: Multispectral Optoacoustic Tomography Detects pH-Low Insertion Peptide Probes In Vivo
- Near-infrared fluorescence molecular imaging of amyloid beta species and monitoring therapy in animal models of Alzheimer's disease
- SPECT- and Fluorescence Image-Guided Surgery Using a Dual-Labeled Carcinoembryonic Antigen-Targeting Antibody
- Noninvasive Molecular Imaging Using Reporter Genes
- Genetically Engineered Oncolytic Newcastle Disease Virus Effectively Induces Sustained Remission of Malignant Pleural Mesothelioma
- Generation of a highly inducible Gal4->Fluc universal reporter mouse for in vivo bioluminescence imaging
- Optical Imaging: Skin Cancer Imaging