


Abstract
Bystander and low–dose-rate effects influence the dose–response relationship in a manner not predicted by current dosimetric methodologies. Radiation-induced bystander effects refer to biologic responses in cells that are not traversed by an ionizing radiation track and, thus, not subject to direct energy deposition; that is, the responses occur in nonirradiated cells. Low–dose-rate hypersensitivity effects have been documented as a reduction in the survival of cells irradiated at dose rates of 0.1–1.0 Gy/h, with total doses ranging from 1.5 to 5 Gy. For humans undergoing external radiotherapy, evidence of bystander events has been observed in the form of abscopal effects, wherein irradiation of one portion of the anatomy affects a portion outside the radiation field, whereas low–dose-rate hypersensitivity has not been described. In this report, the historical literature is briefly reviewed, key experiments are summarized, and current understanding of the factors thought to be involved in the bystander and low–dose-rate effects is conveyed. The mechanisms associated with these events are still being investigated, and questions remain on their impact in radionuclide therapy. Although current findings do not yet sufficiently justify changing traditional dose estimates used to predict the outcomes of radionuclide therapy, it is important to appreciate the potential importance of these effects and to begin revising methods to reflect the emerging empiric and mechanistic knowledge.
This paper reports on the proceedings of a continuing medical education session sponsored by the MIRD Committee at the 53rd annual meeting of the SNM. The objectives of the paper are to review briefly the historical literature, to summarize key experiments, and to describe the current understanding of mechanisms thought to be involved in bystander and low–dose-rate (LDR) hypersensitivity effects.
Radiation-induced bystander effects are biologic responses in cells that were not traversed by an ionizing radiation track and, thus, not subject to direct energy deposition; that is, the responses occur in nonirradiated cells. These bystander effects take place in the neighbors of irradiated cells or in other nonirradiated cells that have received secreted signals from irradiated cells. As such, bystander effects are somehow communicated from an irradiated cell to a nonirradiated bystander cell via cell-to-cell gap junctions (1) or by the secretion or shedding of soluble factors (2). The precise nature of factors that mediate the bystander effect is unknown, but reactive oxygen and nitrogen species and various cytokines have been implicated (3–6). Radiation-induced bystander effects have been extensively documented in several recent reviews (7–10), which have described both detrimental (e.g., DNA strand cleavage, chromosomal damage, and cytotoxicity (11)) and potentially beneficial (e.g., radioprotection (12)) bystander effects.
Although the bystander effect is widely considered a new concept, reports that biologic entities may be inactivated equally by ionizations within the entity or in the surrounding medium have existed since the 1940s (13), and clastogenic factors in plasma from radiotherapy patients were first observed in the 1950s (14,15). The current high level of interest in bystander effects was sparked by Nagasawa and Little in 1992 (16). Using a very low fluence α-particle microbeam, they found that more cells had sister chromatid exchanges than were predicted by calculations of cell traversal probability. Since then, studies have demonstrated that bystander effects induced by high linear energy transfer (LET)—but not those induced by low LET—are dependent on cellular interaction and functioning gap junctions (13,17).
The bystander effect has been shown to be contingent on the dose delivered to the targeted cells. Figure 1 shows that when 10% of the cells in a dish are irradiated with an exact number of α-particles (high LET, with average LET equaling 90 keV/μm), survival of the nonhit cells in the dish is reduced, with the decline in survival depending on the number of hits experienced by the traversed cells (18). Three-dimensional cell culture systems have also been used to investigate the impact of various fractions of irradiated cells on cell cluster survival (19–21). Bystander events are observed when survival of all cells making up the cluster is less than that predicted from the fraction of irradiated cells (20). Such studies, in vitro, have shown bystander effects for both β-particle and Auger-electron emitters. Bystander effects have recently been demonstrated, in vivo, for Auger-electron emitters (22).

Cell survival of nonhit cells when 10% of cells in dish are hit with defined number of α-particles. Because results are normalized to plating efficiency, survival below 100% for nonhit cells reflects bystander effect. In this study, effect appears to be dose-dependent; α-particles were aimed at centroid of nucleus of each targeted cell. (Reprinted with permission from the Health Physics Society, the National Council on Radiation Protection and Measurements, and Dr. Eric J. Hall (13)).
EVIDENCE FOR BYSTANDER EFFECTS
In Humans
The preponderant support in the clinical literature for bystander-like effects may be found in outcomes of external radiotherapy treatment. There is evidence of reactions occurring outside the defined zone of radiation absorption. First described over 50 y ago by Mole (23), who called these out-of-field events “abscopal effects,” these distal/nontargeted effects have been documented sporadically since then. It should be stressed that abscopal effects are not bystander effects in the traditional sense (24) but refer to radiation responses in areas separate from the irradiated tissue and are presumably mediated by secreted soluble factors.
In both radiotherapy patients and external-beam–irradiated animal models (i.e., low LET), most reports on abscopal effects refer to antitumor consequences outside the radiation field (25–35). In the clinical setting, these include regression of hepatocellular carcinomas after radiotherapy to treat a tumor at the base of the spine (25) and histologic changes in metastatic lymph nodes in some women treated for breast cancer (28). Similar results have been observed in animal models. For example, Law and Mole (36) described an abscopal effect on the thymus after applying 5 Gy of irradiation to the posterior half of the weanling rat. The outcome in this case was reduction in weight and DNA content, consistent with the decrease seen after whole-body exposure. Camphausen et al. (26) irradiated the non–tumor-bearing legs of C57BL/6 (wild-type p53) and B6.129S2-Trp53tm1Tyj (p53 null) mice and observed significant growth delays of distally implanted Lewis lung carcinoma and T241 fibrosarcoma cells. The authors found a dose-dependent inhibition of tumor growth, with five 10-Gy fractions leading to a greater inhibition than twelve 2-Gy fractions. This study provides some mechanistic insights for the observations and implicates p53 as a key mediator of the radiation-induced abscopal effect. In addition, these data suggest that pathways downstream from p53 are important in eliciting this response.
With Unencapsulated Radionuclides
The assessment of possible bystander effects is more difficult when they are due to the decay of internally administered radionuclides than when they are due to external irradiation. This difficulty exists because many of the radionuclides used for therapy emit photons as well as α-particles and electrons, with ranges much greater than the mammalian cell diameter (∼10 μm). Only a few reports have examined the bystander effect of unencapsulated therapeutic radionuclides. The low-energy, Auger-electron emitters 123I and 125I, the β-emitter 131I, and the α-particle emitter 211At have been investigated (22,37–39).
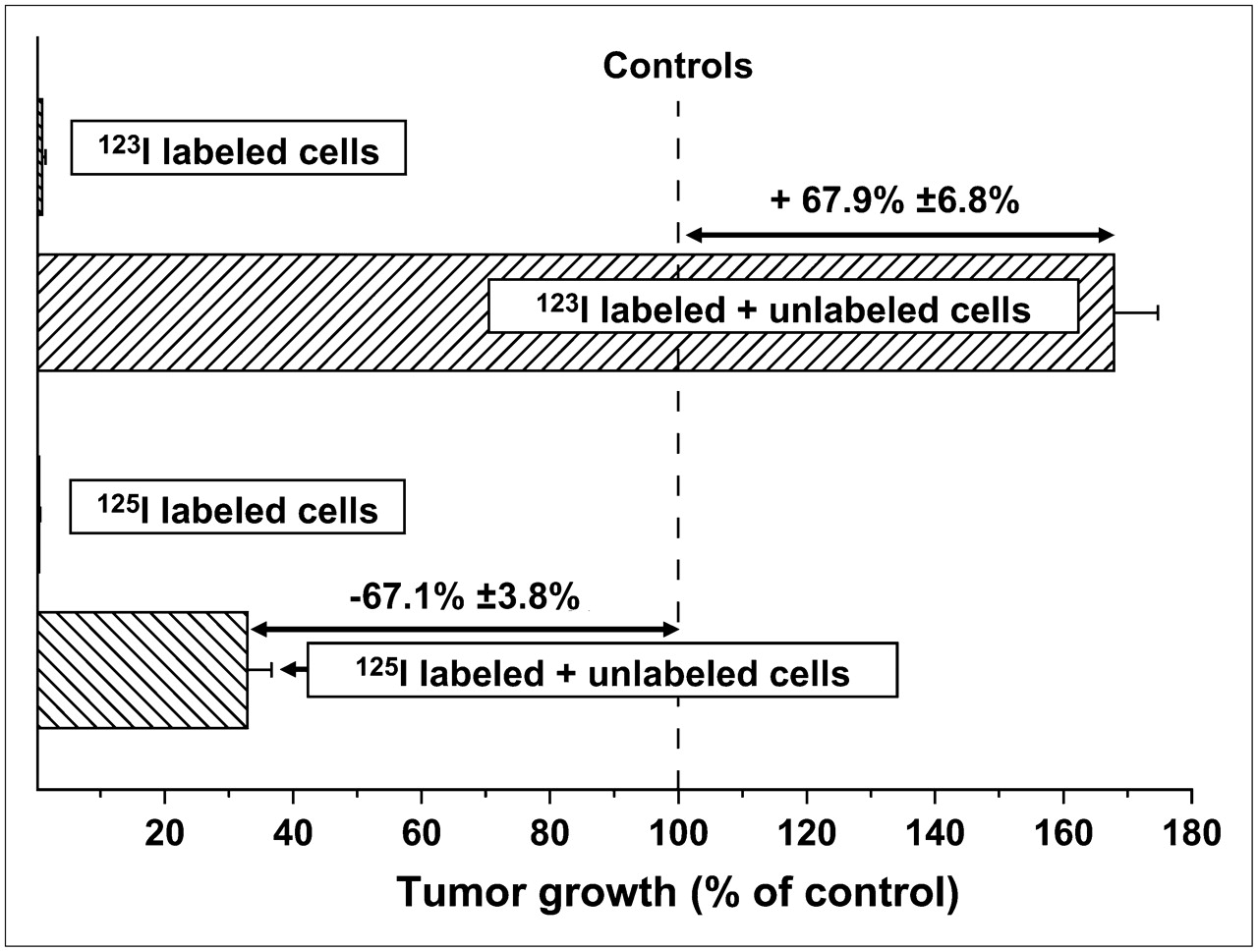
Because of their unique physical decay characteristics, which are functionally equivalent to high-LET-like radiation (40,41), and the virtual absence of cross-fire irradiation of adjacent cells, both 123I (half-life, 13.3 h) and 125I (half-life, 60.5 d) have been used to examine the bystander effect in vitro and in vivo (22,39). The studies performed in vivo demonstrated that the growth of subcutaneously implanted human tumor cells was influenced by the presence of 125I- or 123I-labeled cells that had been mixed with unlabeled tumor cells in the tumor inoculum. Despite the fact that the electron spectra of both radionuclides are identical, injection of a mixture of unlabeled and 125I-labeled cells in mice inhibited the growth of unlabeled, unirradiated cells (22), whereas a mixture with 123I-labeled cells enhanced the growth of unlabeled cells (Fig. 2) (39). Similar inhibitory (125I) and stimulatory (123I) bystander effects were also observed when the radiolabeled cells were incubated in vitro with unlabeled cells (39). Previously, it had been reported that cell survival in a 3-dimensional tissue culture model was similarly compromised when cells were cocultured with 125I-labeled cells (37). These data indicate that both the inhibition (125I) and the stimulation (123I) of tumor growth are a consequence of bystander effects initiated within and generated by the radioiodine-labeled cells.
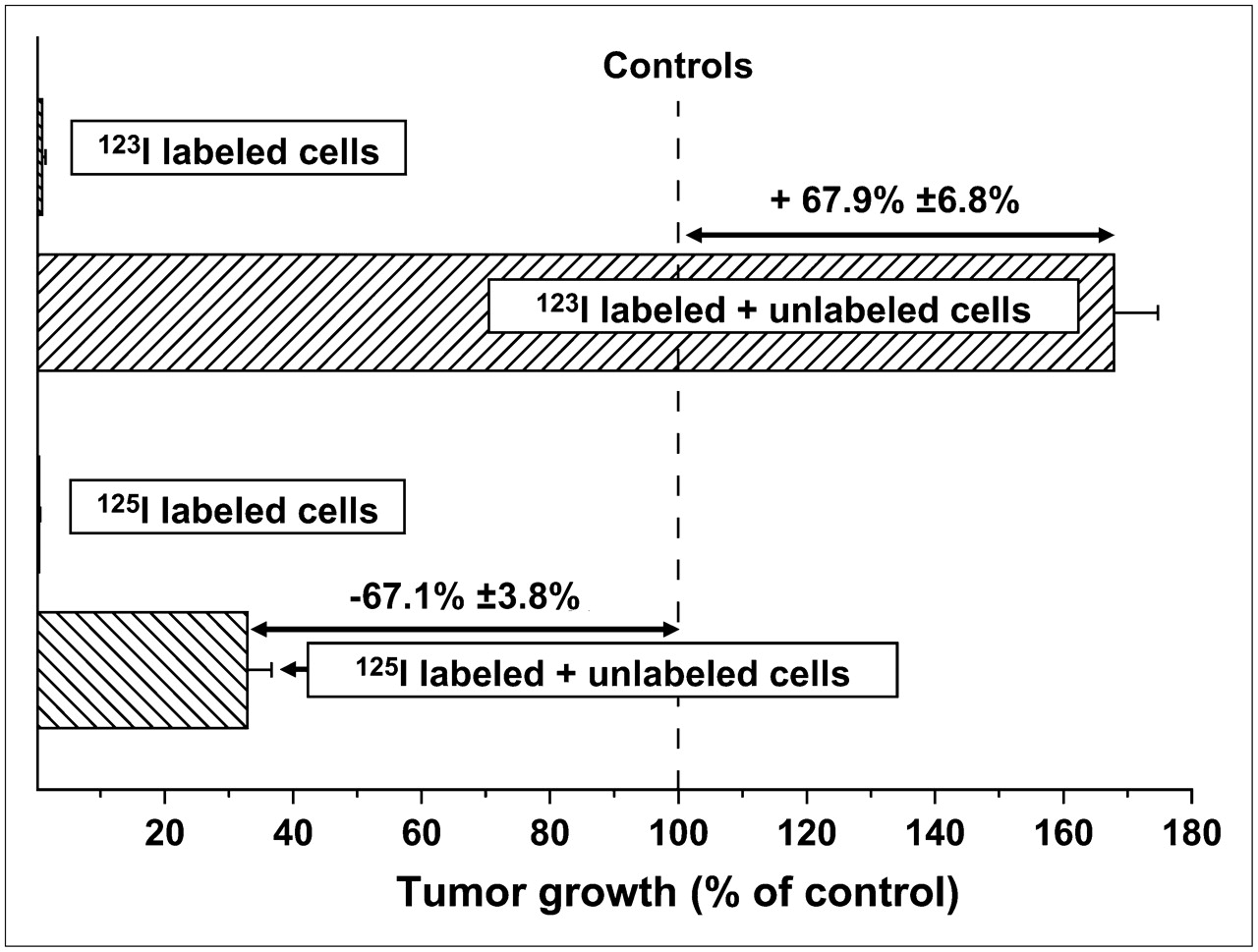
Inhibitory and stimulatory effects of mixtures of unlabeled and 125I- or 123I-IUdR–labeled cells, respectively, on subcutaneous tumor growth in vivo.
In both the animal-model and the cell-culture protocols, irradiated and unirradiated cells exist near one another and are bathed in whatever factor or factors are secreted. Several papers have discussed the involvement of oxidative metabolites and gap-junction intercellular communication in radiation-induced bystander effects (1,15,42,43). In particular, the transfer of genetic instability from irradiated cells to neighboring, unirradiated cells has been emphasized (42–44). Some investigators have also identified secreted factors, including transforming growth factor–β1 (3) and interleukin (IL)-8 (5), that mediate bystander effects in vitro but do not require the existence of oxidative metabolites or gap junctions (5). In a recent study, Boyd et al. (38), examining the ability of medium from cultures in which cells were prelabeled with 123I, 131I, or 211At to inhibit cell proliferation in vitro (clonogenic survival), demonstrated effective cell killing for all 3 therapeutic radionuclides. Similarly, in a preliminary attempt to identify the soluble factors participating in the inhibitory and stimulatory bystander effects, Kishikawa et al. tested the supernatants from 123I-labeled cells in culture for their ability to stimulate tumor-cell proliferation and observed an increase in cell growth (39). These findings suggest that one or more signaling pathways are activated by the decay of such therapeutic radionuclides in the radiolabeled cells and that this, in turn, leads to the secretion of one or more factors that are then transferred to the unirradiated bystander cells, with the consequent induction of an inhibitory (125I) or a stimulatory (123I) event.
The bystander effect induced by radioactive decay introduces a new concept that affects our views on risk assessment and therapeutic efficacy after the administration of therapeutic radiopharmaceuticals to patients. Traditionally, dose is estimated by averaging the radiation dose to cells within a tissue, organ, or tumor mass from radioactive atoms on or within the cells (self-dose) and the radiation dose from radionuclides in or on other cells or in the extracellular fluids (cross-dose). Such absorbed dose estimates have played an important role in determining the amount of diagnostic or therapeutic radioactivity to be administered to patients. When a bystander effect is factored in, the actual radiobiologic response will be greater or less than that predicted by dosimetric estimates alone. Current findings (22,37–39) do not yet sufficiently justify changing traditional dose estimates in the prediction of radiotherapeutic effects. Many questions remain. Is the in vivo bystander effect restricted to the highly specific damage to DNA by ionization secondary to Auger-electron cascades, or is it also seen when such radionuclides deposit their energies within the cell cytoplasm or membrane? Will this phenomenon, in general, occur in vivo when the bystander cells also receive a nonlethal dose consequent to long-range charged-particle traversal (cross-fire) or photon irradiation? Is the bystander effect the same in various normal cell lineages? Is the phenomenon different in tumor cells and among various types of tumors? Does the dose received by the irradiated cells affect the extent and nature of the bystander effect? Will the dose rate make a difference? And finally, will the cells undergoing a bystander effect induce a bystander effect in neighboring cells (a domino effect)? Additional studies are required to address these questions.
DOSE-RATE EFFECTS
LDR irradiation, or irradiation at 0.1–1.0 Gy/h, with total doses ranging from 1.5 to 5.0 Gy, has been reported to affect multiple biologic processes that are important determinants of cell survival in both tumors and normal tissues (45). LDR irradiation in the range 0.01–0.1 Gy/h has been less extensively studied but has recently become an area of active investigation because of data suggesting that such very LDR irradiation may enhance the radiosensitivity of tumors to subsequent larger doses of acute high–dose-rate (HDR) irradiation (45).
Effects on Tumor Cells
Numerous studies have assessed the relative effect of dose rate on tumors, with conflicting results that may be dependent, in part, on tumor type and experimental model or design (45). The existence of an inverse dose-rate effect was first reported by Mitchell et al. (67,68), and these findings were initially attributed to incomplete repair of DNA damage and arrest of cells in the radiosensitive, G2 phase of the cell cycle. Since then, there have been multiple reports of an inverse dose-rate effect in a variety of tumor types, with some studies demonstrating the absence of an association between G2 arrest and radiosensitivity and implicating other mechanisms of action.
The issue of LDR hypersensitivity is particularly relevant to systemically targeted radiation therapy using radionuclides. Several animal studies have addressed the question of the relative efficacy of radioimmunotherapy compared with conventional HDR radiation therapy (48,49). These studies concluded that the low-dose/LDR irradiation associated with radioimmunotherapy can be more effective than HDR irradiation in some tumor models. The consequences were especially profound in a murine syngeneic B-cell lymphoma model in which dose-equivalent tumor-specific 131I-labeled antibody, nonspecific isotope-matched 131I-labeled antibody, LDR γ-irradiation, and fractionated HDR tumor irradiation were compared (50).
Effects on Normal Tissues
Although the literature reflects mixed findings, most data suggest that normal tissues are able to repair LDR-induced DNA damage more effectively than acute HDR-induced DNA damage. DNA repair occurs primarily during p53-dependent, delayed cell cycle progression, with the relative radiosensitivity of normal tissues contingent, in part, on the tissue type. For example, LDR total-body irradiation preferentially spares nonhematopoietic tissues (51), and toxicity data from radioimmunotherapy trials have demonstrated that normal tissues (such as the kidney and gastrointestinal mucosa) have a higher tolerance to LDR than HDR irradiation (52,53). Many studies also suggest that there is a decreased risk of genomic instability and mutation with decreasing dose rate (54).
Low-dose/LDR irradiation has been reported to induce radioresistance in some normal cells (previously referred to as hormesis or radioadaption) or to increase immunocompetence. Observations in preclinical models suggest an effect of low-dose/LDR irradiation on immune responses; for example, protection of mice against Friend murine leukemia virus (55–57); delayed onset of thymic lymphoma, acute myelogenous leukemia, and some transplanted solid tumors (58–60); decreased pulmonary (61,62) and lung and nodal metastases (63); and suppressed local tumor growth (62,64). In contrast, other studies have found total-body irradiation to be associated with increased tumor progression (65,66). These discrepancies may be due, in part, to differences in tumor type, tumor burden, dose rate, and total dose studied.
Key Present-Day Experiments
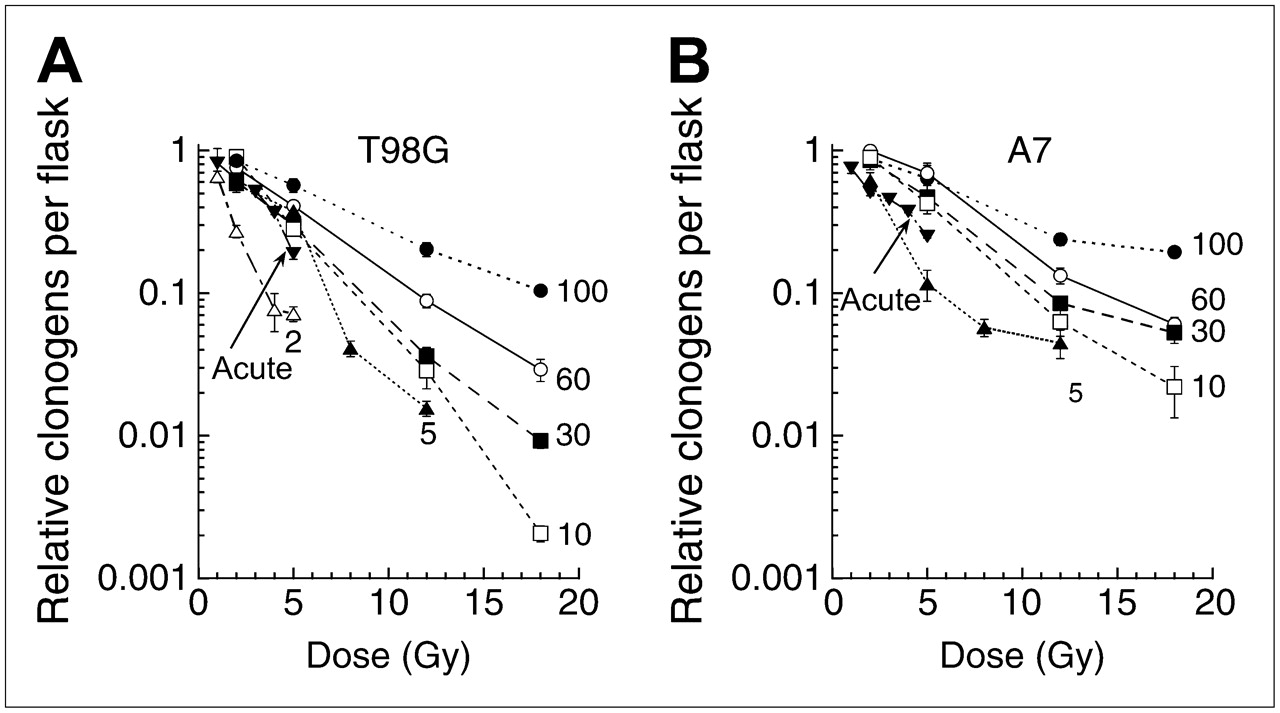
LDR irradiation at 3 cGy/h for 24–72 h was found to sensitize U251 glioma cells, in vitro, to HDR irradiation at 1.1 Gy/min (2–10 Gy) (67). A similar outcome occurred in vivo in U251 xenografts with 5 cGy/h LDR combined with fractionated HDR irradiation (67). It has been suggested that the response to LDR irradiation correlates with the susceptibility to induced hypersensitivity by low acute doses (<0.05 Gy) (68) and that a minimum dose-rate effect may occur at an LDR of 0.06–0.6 Gy/h (19). The studies from Joiner's laboratory (46,47) illustrate the inverse dose-rate effect and the effectiveness of low-dose hypersensitivity when radiation is delivered continuously to proliferating cell populations at dose rates below 1 Gy/h. In these experiments, total cytotoxicity was assessed as the sum of 2 factors. First, radiation-induced reduction in cell replication during extended irradiation periods was accounted for by measuring cell number relative to sham-irradiated samples, to give a relative cell yield. Second, a high-precision, colony-forming assay was used to measure the surviving fraction of cells in irradiated samples relative to sham-irradiated samples. Multiplying the relative cell yield by the surviving fraction gives relative clonogens per flask—the best measure of the overall lethal effect of a radiation exposure on a dynamic cell population in vitro—and parallels the concept of “relative clonogens per tumor” (69). To illustrate, Figure 3 shows the response of 2 glioma cell lines to continuous irradiation at less than 100 cGy/h, which is the dose-rate level at which permanent iodine or palladium implants typically are used. As the dose rate is reduced further, the cells become increasingly responsive. X-ray exposures were 3 times more effective per unit dose at a dose rate of 5 cGy/h than at a dose rate of 100 cGy/h, and in the T98G cell line (Fig. 3A), the lowest dose rate tested (2 cGy/h) was 5 times more effective per unit dose than a dose rate of 100 cGy/h and was even more effective than acute exposures. These studies showed that G2 accumulation is not the explanation for the increased radiosensitivity at lower dose rates. Rather, the opposite occurs: progression of cells into mitosis increases as the dose rate is reduced. This is consistent with the mechanism underlying the increased response of cells to small acute doses, as elucidated by Marples et al. (70), in which the “early” G2 arrest, which allows DNA repair to take place in irradiated G2 cells, occurs only if sufficient DNA damage (equivalent to a single dose of approximately 0.2 Gy) has been produced.

Survival curves after exposure of asynchronously growing T98G (A) and A7 (B) human glioma cells to LDR 60Co γ-radiation. Numbers represent dose rate in cGy/h. Number of relative clonogens per flask was calculated by multiplying surviving fraction by relative yield. Each data point is plotted as mean ± SEM. Acute dose rate was 33 Gy/h.
Understanding the mechanism of action of LDR hypersensitivity is important for determining the optimal way to combine LDR irradiation with conventional HDR radiation therapy. This area of investigation is active, and relevant findings are summarized in the next section.
Current Understanding of Mechanisms
LDR hypersensitivity is mediated, at least in part, by cell cycle redistribution, with accumulation of cells at the G2/M interface; effects on sublethal and potentially lethal damage repair; selective effects on tumor vasculature; reoxygenation during protracted exposure; potentiated induction of apoptosis; and effects on gene expression. The relative contribution of these potential mechanisms of action of LDR irradiation may vary as a function of tumor type, microenvironment, and experimental model and conditions. Data have supported and refuted some of the proposed LDR mechanisms of action.
The literature contains conflicting data regarding the role of G2/M arrest on susceptibility to LDR irradiation (45), but recent studies, such as those performed on prostate cancer cell lines, provide compelling data demonstrating that radiation-associated perturbations in cell cycle distribution are not the primary determinants of tumor cell killing by LDR irradiation (71). Whereas the mechanisms of action of inverse dose-rate effects are still somewhat unclear, recent reports suggest that low-dose/LDR irradiation decreases activation of ataxia–telangiectasia, mutated (ATM) and induces histone-H2A, family member X (H2AX) downstream (72,73). These observations are consistent with the hypothesis that sensing of DNA damage by ATM, decreases with LDR irradiation, resulting in decreased activation of the early DNA damage response (73). Furthermore, decreased induction of H2AX with LDR is not dependent on cell cycle (73). The observation that poly (ADP-ribose) polymerase-1 inhibition increases sensitization of tumor cells to low-dose irradiation (74) further substantiates the role of the ATM pathway in low-dose/LDR effects. Although decreased activation of this pathway by LDR would suggest that inefficient detection of DNA double-strand breaks plays an important role in the efficacy of LDR, not all studies have demonstrated a correlation between dose rate, double-strand breaks, and survival (75). The frequency of cells with micronuclei has also been reported to be inversely related to dose rate and directly related to total dose (76).
Multiple studies, primarily with lymphohematopoietic malignant cell lines and xenograft tumors, have demonstrated a relationship between inverse dose-rate effects, susceptibility to apoptosis (77–79), and downregulation of B-cell lymphoma 2 (80,81). Conversely, LDR hypersensitivity can be abrogated by priming with HDR irradiation, which has been reported to cause a relative decrease in apoptosis, p53, and bax expression (82).
Dose-rate effects on gene expression may be important determinants of LDR consequences. Gene array analyses have revealed 2 major clusters of radiation-induced genes. Effects on genes involved with cell cycle regulation in ML-1 cells were found to be independent of dose rate, whereas those affecting apoptotic potential and apoptotic pathways were dependent on dose rate (83). In this cell line, decreased dose rate was associated with an alteration in expression of apoptotic genes that was protective, with a linear induction of p53-regulated genes (e.g., regulators of cell cycle progression) (83).
LDR total-body irradiation is used to decrease toxicity to normal tissue and has been reported to have greater effects on tumor than expected for the delivered dose, suggesting an immunity-enhancing aspect of total-body irradiation. Indeed, total-body irradiation has been found to influence cytokine secretion and effector cells in multiple ways (84–86) (e.g., increased interferon-γ and IL-2, increased IL-2R expression on T cells, T-cell activation, decreased serum corticosteroids, decreased levels of immunosuppressor T cells (87–91), macrophage and natural killer cell activation (92), increased expression of adhesion molecules resulting in enhanced leukocyte trafficking (93,94), and enhanced responses to tumor antigens (95)). Low-dose/LDR-induced natural killer cell activity (96,97) may be related to low-dose/LDR-increased IL-12 and IL-18 secretion. A comprehensive review by Liu further discusses dose-rate effects on antigen-presenting cells and T lymphocytes (98) in terms of mechanisms of action and activation pathways.
Radioadaption in nonimmune cells (99–101) occurs via nonimmune mechanisms. Data that are largely indirect suggest that low-dose/LDR irradiation affects the efficiency of DNA repair (102) and that DNA-dependent protein kinase activity may play a role in these events (103,104). Protein kinase C–mediated signaling (105), interferon-γ (106,107), and induction of antioxidant enzymes (108,109) may also contribute to the process of radioadaption. Some of the most provocative data come from studies by Kipnis et al. (110–112) reporting that low-dose total-body irradiation or total lymphoid irradiation increased neuronal survival and enhanced recovery after optic nerve or spinal cord injury and that these effects were secondary to radiation-induced type 1 helper T-cell–derived neuroprotection.
SUMMARY
Traditionally, doses are estimated by averaging the radiation dose to cells within a tissue, organ, or tumor mass from radioactive atoms on or within the cells (self-dose) and from radionuclides in or on other cells or in the extracellular fluids (cross-dose). Such absorbed dose estimates have played an important role in determining the amount of radioactivity to be administered to patients in diagnostic and therapeutic procedures. Bystander and LDR effects influence the dose–response relationship in a manner that cannot be predicted by current dosimetric methodologies. Further studies are needed to elucidate more completely the mechanism of action of both bystander and LDR effects on tumors, normal tissues, and immune responses at the molecular level and to explain how and why bystander and low-dose/LDR effects depend on cell type, dose rate, total dose, radiation quality, endpoint measured, time, and microenvironment. Nevertheless, both bystander and low-dose/LDR events are currently relevant and presumably operative in systemically targeted radionuclide therapies.
In some clinical situations, bystander effects may lessen the impact of nonuniformity in spatial distribution of the absorbed dose. A particularly promising area for future investigation is the use of LDR irradiation as a component of combined-modality therapy to increase tumor cell killing. This kind of irradiation can be used to “prime” tumor cells, enhancing their sensitivity to subsequent HDR irradiation. Studies are needed to determine which tumor types are most sensitive to LDR as a means of increasing sensitivity to HDR irradiation. It is also conceivable that LDR irradiation might be useful when combined with sensitizers, chemotherapy, or biologic response modifiers. Another possible clinical application of LDR is upregulation of antitumor immune responses, with potential utility as a component of vaccine strategies and radioadaption of normal tissues. Mechanistic and optimization studies are needed to exploit fully the therapeutic potential of both bystander and LDR effects to optimally enhance the therapeutic index of radionuclide therapies. In conclusion, although current findings do not yet sufficiently justify changing traditional dose estimates used to predict the outcomes of radionuclide therapy, it is important to anticipate these effects and to begin revising methods to reflect the emerging empiric and mechanistic knowledge.
GLOSSARY
Abscopal effects: Radiation responses seen in areas separate from the irradiated tissue and presumably mediated by secreted soluble factors. Most of these effects refer to antitumor effects.
Acute myelogenous leukemia: A cancer of the white blood cells that progresses quickly and is characterized by the growth of too many immature white blood cells in the blood and bone marrow.
α-particle: A particle consisting of 2 protons and 2 neutrons—produced during α-decay—that is identical to the nucleus of a helium atom.
Ataxia–telangiectasia, mutated: A gene that is activated during DNA repair. Mutations in this gene lead to a disease (ataxia–telangiectasia) that is characterized by hypersensitivity to ionizing radiation.
Auger electron: Monoenergetic electrons emitted from a decaying atom as a result of inner electron shell vacancies.
Bax (Bcl-2 associated X protein): A gene or gene product that promotes apoptosis.
Bcl-2 (B cell lymphoma 2): A gene or gene product that inhibits apoptosis.
β-particle: An elementary particle emitted from a nucleus during radioactive decay. A β-particle is identical to an electron and is emitted from the nucleus with a probability distribution (spectrum) of possible energies.
Bystander effect: Biologic responses to radiation in cells that have not been traversed by an ionizing radiation track and, thus, are not subject to direct energy deposition events; that is, the responses occur in nonirradiated cells that are near irradiated cells.
Clastogenic factors: Agents that cause breaks in chromosomes.
Double-strand break: A break in both strands of DNA. This kind of damage is typically difficult to repair and often leads to cell death.
G2: The period of the cell cycle that represents the gap between the completion of DNA synthesis and the beginning of mitosis.
G2/M: The period of the cell cycle that represents the transition phase between G2 and mitosis (M).
Gy (gray): A unit of absorbed dose equal to 1 J/kg.
HDR: A dose rate typically greater than 50–60 Gy/h.
Histone-H2A, family member X: A protein involved in the recognition or repair of DNA double-strand breaks.
Hormesis: In radiotherapy, a dose–response phenomenon characterized by a beneficial, resistance-inducing effect at a low dose but a deleterious effect at a high dose. This effect has been attributed to “priming” or increased activation of repair and immune protective mechanisms after exposure to a low dose of radiation.
IL-2R: A receptor for IL-2 involved in mediating T-cell activation and the body's immune response to certain cancers.
IL-8: A proinflammatory cytokine that is released by several cell types (e.g., monocytes, macrophages, T cells, endothelial cells, and tumor cells) in response to an inflammatory stimulus.
IL-12: A cytokine that is involved in the differentiation of naive T cells into CD4+ helper T cells. This function is important in resistance against pathogens.
IL-18: A cytokine that shares biologic activities with IL-12.
IL-18 is synthesized as a propeptide that is cleaved to create the active cytokine.
Interferon-γ: A cytokine that is secreted by T lymphocytes and natural killer cells. Interferon-γ has antiviral, immunoregulatory, and antitumor properties.
LDR: A dose rate between 0.1 and 1 Gy/h.
LDR effects: Irradiation at 0.1–1.0 Gy/h, with total doses ranging from 1.5 to 5.0 Gy. This level of irradiation has been reported to affect multiple biologic processes that are important determinants of cell survival in both tumors and normal tissues.
LDR hypersensitivity effects: The observation that in certain tumors and cells LDR irradiation leads to reduced survival.
Linear energy transfer: The average amount of energy lost by a particle to the surrounding medium per unit distance traveled.
MIRD: Medical Internal Radionuclide Dosimetry, standard methods, models, assumptions, and mathematic schema for assessing internal radiation doses from administered radiopharmaceuticals.
Natural killer cell: A type of white cell that is specialized for killing cells recognized by the immune system as foreign.
P53: a gene that codes for a protein that regulates the cell cycle and hence functions as a tumor suppressor. The name is due to its molecular mass: it is in the 53-kDa fraction of cell proteins (P).
Poly(ADP-ribose) polymerase-1: A DNA-binding protein that recognizes DNA strand breaks and is presumed to play a role in DNA repair.
Protein kinase C: A type of enzyme that phosphorylates other proteins that are used to activate or inactivate various cellular processes.
Radioimmunotherapy: Irradiation of cancer cells by therapeutic radionuclides that are delivered using antibodies against antigens exclusively or predominantly expressed on tumor cells.
Total-body irradiation: External radiotherapy of the whole body, often used in preparation for bone marrow or stem cell transplantation.
Transforming growth factor–β1: A cytokine that controls many aspects of cellular function, including cellular proliferation, differentiation, migration, apoptosis, adhesion, angiogenesis, immune surveillance, and survival.
Type 1 helper T cells: CD4+ T cells that are a part of the immune system and are involved in activating and directing other immune cells.
Very LDR: A dose rate between 0.01 and 0.1 Gy/h.
Footnotes
-
↵* NOTE: FOR CE CREDIT, YOU CAN ACCESS THIS ACTIVITY THROUGH THE SNM WEB SITE (http://www.snm.org/ce_online) THROUGH OCTOBER 2008.
-
No potential conflict of interest relevant to this article was reported.
-
COPYRIGHT © 2007 by the Society of Nuclear Medicine, Inc.
References
- 1.↵
- 2.↵
- 3.↵
- 4.
- 5.↵
- 6.↵
- 7.↵
- 8.
- 9.
- 10.↵
- 11.↵
- 12.↵
- 13.↵
- 14.↵
- 15.↵
- 16.↵
- 17.↵
- 18.↵
- 19.↵
- 20.↵
- 21.↵
- 22.↵
- 23.↵
- 24.↵
- 25.↵
- 26.↵
- 27.
- 28.↵
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.↵
- 36.↵
- 37.↵
- 38.↵
- 39.↵
- 40.↵
- 41.↵
- 42.↵
- 43.↵
- 44.↵
- 45.↵
- 46.↵
- 47.↵
- 48.↵
- 49.↵
- 50.↵
- 51.↵
- 52.↵
- 53.↵
- 54.↵
- 55.↵
- 56.
- 57.↵
- 58.↵
- 59.
- 60.↵
- 61.↵
- 62.↵
- 63.↵
- 64.↵
- 65.↵
- 66.↵
- 67.↵
- 68.↵
- 69.↵
- 70.↵
- 71.↵
- 72.↵
- 73.↵
- 74.↵
- 75.↵
- 76.↵
- 77.↵
- 78.
- 79.↵
- 80.↵
- 81.↵
- 82.↵
- 83.↵
- 84.↵
- 85.
- 86.↵
- 87.↵
- 88.
- 89.
- 90.
- 91.↵
- 92.↵
- 93.↵
- 94.↵
- 95.↵
- 96.↵
- 97.↵
- 98.↵
- 99.↵
- 100.
- 101.↵
- 102.↵
- 103.↵
- 104.↵
- 105.↵
- 106.↵
- 107.↵
- 108.↵
- 109.↵
- 110.↵
- 111.
- 112.↵
- Received for publication February 23, 2007.
- Accepted for publication June 22, 2007.





{kind=link}
{kind=link}
{kind=link}