


Abstract
Monitoring of angiogenesis-relevant approaches with functional imaging and histomorphometric analyses is desirable to evaluate the biologic effects. In this study we wished to examine the complex effects of angiopoietin-2 (Ang-2) gene transfer in a rat hepatoma model. Methods: Using a bicistronic retroviral vector for Ang-2, Morris hepatoma (MH3924A) cell lines with Ang-2 expression were generated (Ang-2-MH3924A). In human umbilical vein endothelial cells (HUVECs) cocultured with Ang-2-MH3924A, the proliferative action with or without growth factors were determined. Furthermore, animal experiments were performed to measure effects on tumor growth and perfusion. Finally, tumors were examined by immunohistochemistry and DNA chip analysis. Results: Ang-2−expressing MH3924A enhanced basic fibroblast growth factor−mediated endothelial cell proliferation. Perfusion, as measured by H215O PET, was increased in genetically modified tumors. Consistent with the increased perfusion, micro- and macrovascularization were increased. However, tumor growth was similar to wild-type MH3924A (WT-MH3924A). Proliferating cell nuclear antigen and TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling) staining revealed an increased number of positive cells, indicating a compensation of increased proliferation by enhanced apoptosis. DNA chip analysis showed an induction of angiogenesis-promoting genes, including crucial vascular growth factor receptors, as well as genes related to extracellular matrix (ECM), apoptosis, signal transduction, and oxidative stress. Conclusion: Our results suggest that Ang-2 expression increases perfusion or vascularization, especially in interaction with the vascular growth factor system, without affecting tumor growth. Simultaneous, enhanced expression of genes for ECM, apoptosis, and signal transduction indicates Ang-2's versatile role in angiogenesis including its destabilizing function on ECM and endothelium.
The process of tumor angiogenesis depends on a large number of different angiogenic factors, such as vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), interleukin-8, the angiopoietin (Ang)/Tie-2 system, and several others. These factors need to be produced either by cancer cells or by surrounding or infiltrating host tissue (1).
Besides the VEGF-system, the Ang/Tie-2 pathway is gaining strength as an interesting target for disrupting tumor angiogenesis. Angiopoietins are a family of extracellular ligands that specifically recognize and bind to the extracellular domain of Tie-2 on endothelial cells. Whereas Ang-1 leads to a phosporylation of Tie-2, Ang-2's primary effect is an antagonism toward Ang-1 on Tie-2, resulting in an inhibition of the Tie-2 signal (2). Ang-2's classically implicated role in angiogenesis is a disruption of the stabilizing and endothelial cell interaction mediating signal by the Tie-2-pathway (2,3), which results in destabilization of existing vessels. The gained induction of plasticity in the vessel environment is an essential step for the initiation of angiogenesis (2,4). Ang-2 is suggested to partly facilitate these functions by an induction of extracellular matrix (ECM) proteins such as matrix metalloproteinases (5). However, several studies using Ang-2−overexpressing tumor models reported divergent results on tumor growth and vascularization (6,7). Ang-2's heterogeneous effects seem to be dependent on the environment and its interactions with other angiogenic factors—first and foremost, VEGF. Ang-2 is able to enhance endothelial cell proliferation, in the presence of growth factors such as VEGF or basic FGF (bFGF), but may also induce apoptosis, in the absence of growth factors. Ang-2, therefore, seems to take part in both the destabilizing signals on vessels and the VEGF-induced angiogenesis. Because Ang-2 seems to be endothelial specific, and selectively implicated only in diseases associated with angiogenesis such as cancer (4,5), Ang-2 is a promising target in antiangiogenic tumor therapy.
PET using H215O provides a quantitative measure of tissue perfusion and, therefore, may be used to monitor physiologic effects caused by antiangiogenic approaches. We used PET measurements because absolute quantification is possible and the use of a completely and freely diffusible molecule (water) with an assumed 100% extraction from circulation present advantages in comparison with Doppler or laser-pulsed methods. Furthermore, recent data present evidence that 68Ga-DOTA-albumin (DOTA is 1,4,7,10-tetraazacyclododecane-N,N′,N′′,N′′′-tetraacetic acid) PET can be used as a blood-pool marker in vivo (8). In the present study, functional data were combined with histomorphometric investigations of apoptosis, proliferation, vascularization, oxidative stress, and inflammation, as well as gene array analysis of the expression pattern in cells and tumors to elucidate the complex role of Ang-2 in hepatoma.
MATERIALS AND METHODS
Cell Culture and Generation of Recombinant Cells
Rat Morris hepatoma (MH3924A) cells were cultured in RPMI 1640 medium (Gibco), supplemented with 292 mg/L glutamine, 100,000 IE/L penicillin, 100 mg/L streptomycin, and 20% fetal calf serum. Human umbilical vein endothelial cells (HUVECs) were cultured in Medium 199 (Life Technologies), supplemented with 292 mg/L glutamine, 100,000 IE/L penicillin, 100 mg/L streptomycin, and 10% fetal calf serum.
Ang-2 Gene Transfer and Northern Blots
For transfer of the Ang-2 gene, we used a bicistronic retroviral vector consisting of the human Ang-2 gene (obtained from George D. Yancopoulos (2)) and the neomycin resistance gene cloned downstream of the elongation factor-1α promoter. To ensure simultaneous expression of the genes coding for the Ang-2 and the neomycin resistance and stabilization of the messenger RNA (mRNA), a synthetic intron and an internal ribosomal entry site from encephalomyocarditis virus was inserted between the genes (9).
For transient packaging of the retroviral particles, a lipofection of the transient packaging cell line BOSC23 and infection was done as described (9). The cells were treated with 0.8 mg/mL neomycin until resistant cell lines were established. These were further incubated in neomycin-containing medium.
Total RNA was isolated using Trizol reagent (Roche) and mRNA expression was assessed by Northern blots, using a 32P-labeled 950−base-pair probe delivered by Nsi I/Pvu II digestion of the Ang-2−carrying vector pSKhtl2b.
Measurement of Cell Proliferation
HUVECs were isolated from umbilical cords as described (10). To determine the effects of Ang-2 on endothelial cell proliferation, HUVECs were cocultured in 6-well plates with wild-type (WT)-MH3924A or Ang-2-MH3924A, using culture inlets with a pore size of 3 μm (Nunc) as a separate chamber for the MH cell lines. Before the coculturing, WT- and Ang-2-MH3924A cells were adapted to supplemented Medium 199. The setting was performed in the absence of growth factors or in the presence of 1 ng/mL bFGF. After coculture for 72 h, proliferation was determined using a Coulter Counter.
Animal Studies
All animal studies were performed in compliance with the national laws relating to the conduct of animal experimentation. The in vitro doubling time of WT- (19.7 h) and Ang-2-MH3924A (18.1 h) cells was similar. After subcutaneous inoculation of WT or Ang-2 cells (2 × 106 cells/animal) into the thigh of ACI or RNU (immunodeficient nude) rats (Charles Rivers Laboratories), tumor growth was measured using calipers. PET studies of tumor perfusion were done using animals weighing 200−250 g. Perfusion and blood pool were examined in tumors with diameters between 10 and 13 mm after an intravenous bolus injection of 70−150 MBq H215O in 0.3 mL and 4−16 MBq 68Ga-labeled albumin [68Ga-DOTA-albumin] (same animals and session; the albumin PET scan was performed after the activity of H215O decayed). All PET studies were done as dynamic measurements (20 × 3 s, 6 × 10 s, 4 × 15 s, 6 × 30 s for H215O and 12 × 10 s, 3 × 20 s, 4 × 30 s, 10 × 60 s for 68Ga-DOTA-albumin) acquired in 2-dimensional mode using a matrix of 256 × 256 on an ECAT HR+ (Siemens/CTI) scanner (pixel size, 2.277 × 2.777 × 2.425 mm; transaxial resolution, 4.3 mm). A transmission scan was done for 10 min before tracer administration with 3 rotating germanium pin sources to obtain cross-sections for attenuation correction. After iterative reconstruction with the space-alternating generalized expectation maximization method ([SAGE] 0 subsets, 20 iterations) applying median root prior correction, the dynamic PET data were evaluated as described earlier using the PMOD software package (10). Time–activity curves were created using volumes of interest (VOIs). A VOI consists of several regions of interest over the target area. Perfusion studies with H215O were evaluated using a 1-tissue-compartment model with the mean value of the VOI data obtained from the heart as the input function as previously described (10). This procedure has an estimated recovery error of 10%, which can be neglected for the purposes of this study. The 68Ga-DOTA-albumin uptake was expressed as the mean standardized uptake value between 10 and 15 min after injection when the time–activity curve reaches a plateau (8).
Tissue Preparation and Immunohistochemistry
To obtain qualitative and quantitative data on tissue properties, which may influence tumor perfusion, immunohistochemical analyses of proliferation, apoptosis, necrosis, immune reactions, and vessel density were performed. Rat tumors were fixed with IHC zinc fixative (BD Biosciences) or 4% paraformaldehyde (and routinely embedded in paraffin) or shock-frozen in liquid nitrogen−cooled isopentane and kept at −70°C in a freezer until used for immunohistochemical investigations. Immunohistochemistry of the tumors was routinely performed according to procedures as described earlier (10). We used the monoclonal antibodies mouse antirat-CD31 (1:500; BD Pharmingen) (to detect endothelial cells), mouse anti-α-actin (1:100; Roche) (to identify smooth muscle cell−containing vessels), mouse antiproliferating cell nuclear antigen ([PCNA] 1:500; DakoCytomation) (for proliferating cells), mouse antirat CD11b (1:500; BD Pharmingen), and mouse antihuman-macrophage migration inhibitory factor ([MIF] 1:50; R&D Systems GmbH) (to mark inflammatory macrophages). As a negative control, the primary antibody was omitted or control immunoglobulin was applied. Apoptosis was measured on 4% paraformaldehyde-fixed paraffin-embedded tumor cross-sections by TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling) technique using a commercially available kit (Oncor). Immunoreactive cells in tumor cross-sections were recorded by a video camera (Olympus HCC-3600 P high gain) and quantified using a computer-assisted image analysis system developed by our group (VIBAM 0.0-VFG1-frame grabber) as previously described (10).
Gene Arrays
Aliqots of the WT- and Ang-2-MH3924A tumors were stored in Trizol to extract RNA according to the manufacturer's specifications. RNA was purified using the RNeasy kit (Quiagen Inc.) as described (10). Gene chip expression arrays were performed using The Rat Genome U34 Set (Affimetrix Inc.). The RNAs of Ang-2-MH3924A (n = 6) and WT-MH3924A (n = 7) tumors were pooled and evaluated on separate gene chips as suggested by the manufacturer. After normalizing the values to the β-actin expression, ratios of modified versus WT tumors were calculated. Because very low signal intensity results in an increased error rate, only genes above a cutoff value of 0.5% of the β-actin signal were used for further analysis. The ratios of Ang-2- versus WT-MH3924A tumors were calculated and values above 2 and below 0.5 were interpreted as significant. The whole experiment was repeated revealing a high correlation of the signals in both experiments (r = 0.98 [tumor]; r = 0.99 [cells]).
Data Analysis and Statistics
Results are presented as mean ± SEM. Statistical comparisons between 2 groups were performed by the Mann–Whitney U test or by the unpaired Student t test using the SIGMASTAT program (Jandel Scientific). P ≤ 0.05 was considered statistically significant.
RESULTS
Enhanced Proliferation of HUVECs Cocultured with Ang-2-MH3924A in Presence of bFGF
The successful transfer of the Ang-2 gene into MH3924A and the overexpression of the gene were determined by Northern blotting (not shown).
Coculture experiments were performed either in the absence of growth factors or in the presence of 1 ng/mL bFGF, to determine Ang-2's ability to enhance the proliferative effect of bFGF. In HUVECs cultured without MH3924A, the addition of 1 ng/mL of bFGF had no effect on the proliferation compared with HUVECs cultured without growth factor (Fig. 1). Furthermore, in HUVECs cocultured with WT-MH3924A, 1 ng/mL bFGF was not sufficient to induce an increase of proliferation. In contrast, stimulation of HUVECs with 1 ng/mL bFGF cocultured with Ang-2-MH3924A resulted in a significant increase in proliferation of HUVECs (Fig. 1). In the absence of bFGF, proliferation of HUVECs was unaltered, independent of coculture with WT- or Ang-2-MH3924A, or cultured alone, showing that this proliferative effect is not directly mediated by Ang-2.

Proliferation of HUVECs with or without (w/o) bFGF (1 ng/mL) cocultured with WT-MH3924A or Ang-2-MH3924A. Stimulation with low concentration of bFGF is not sufficient to induce proliferation of HUVECs. Note that Ang-2-MH3924A only induces proliferation in HUVEC in presence of bFGF. Values are mean ± SEM (n = 3 in duplicate; P = 0.003, Ang-2-MH3924A with vs. without bFGF; P = 0.004 vs. WT-MH3924A with bFGF).
Tumor Growth, Perfusion, Vascularization, and Vascular Permeability in WT- and Ang-2-MH3924A Tumors: PET and Immunohistomorphometry
For the evaluation of tumor growth, RNU rats were used to exclude immunologic influences on tumor proliferation. At day 38 after inoculation in RNU rats in vivo, volume-based tumor growth of Ang-2-MH3924A was comparable with that of WT-MH3924A (Fig. 2). Similar results were obtained with ACI rats (data not shown).
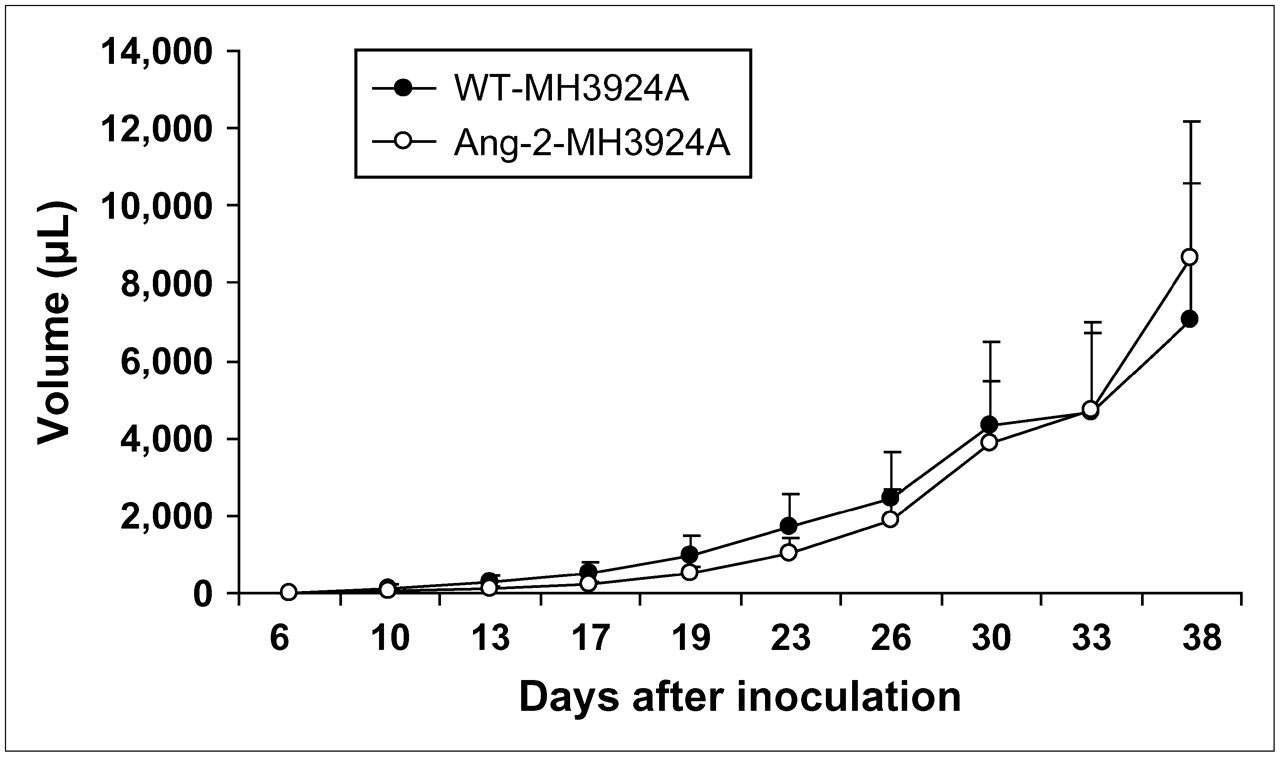
Tumor growth in RNU rats after transplantation of WT-MH3924A or Ang-2-MH3924A until 38 d after inoculation. Values are mean ± SEM (n = 8 for each group).
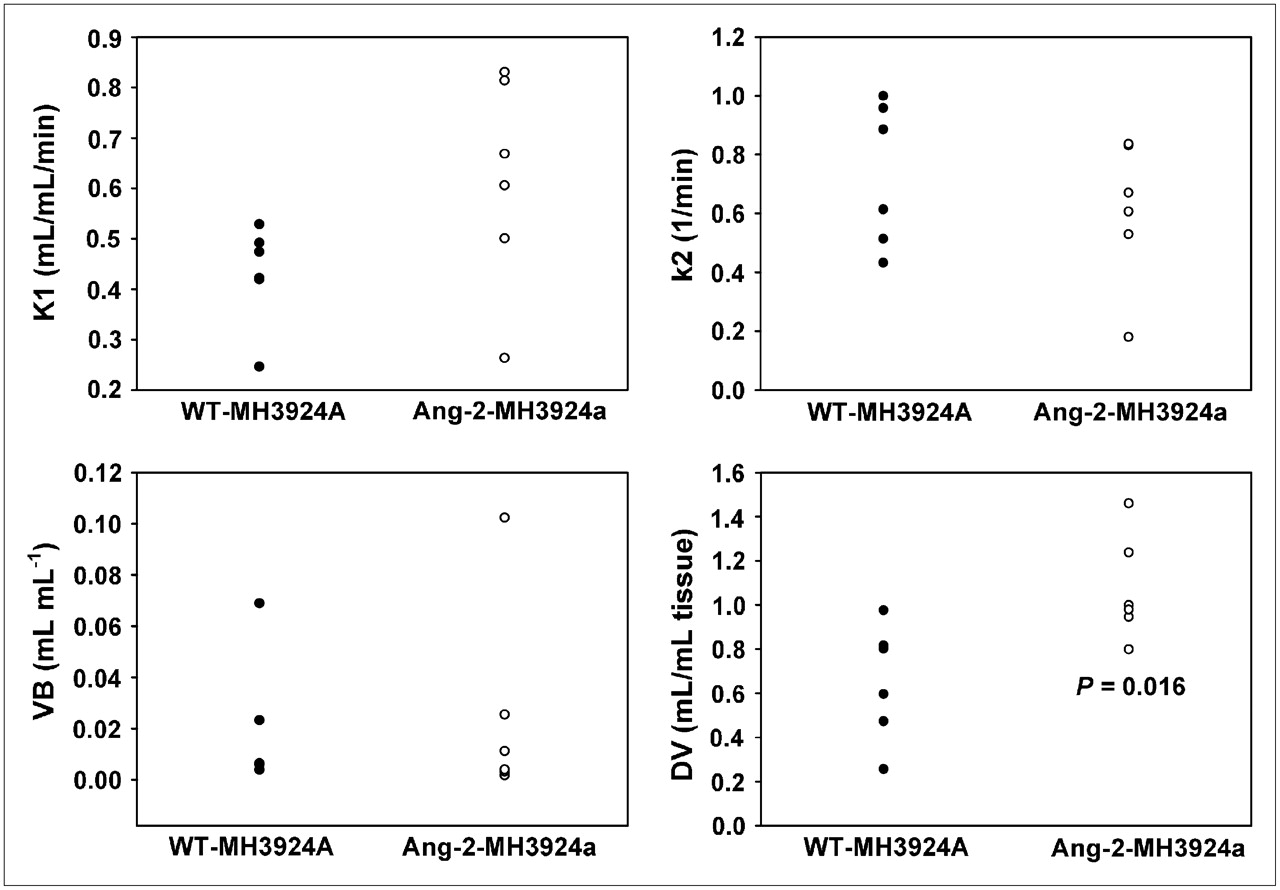
Dynamic PET measurements with H215O were performed at similar tumor sizes (10- to 13-mm diameter). Pharmacokinetic analysis of H215O PET data revealed that tumor perfusion, represented as K1 (in mL × mL tissue−1 × min−1), showed a mean increase of 42% (P = 0.08) in Ang-2-MH3924A (0.61 ± 0.08) in comparison with WT-MH3924A (0.43 ± 0.04). However, the flow of tracer from tumor tissue back to the blood vessels, represented as k2, was unchanged (P = 0.39) in Ang-2-MH3924A (0.61 ± 0.09) compared with WT-MH3924A (0.73 ± 0.09). Additionally, the fractional volume of distribution (DV) significantly increased in Ang-2-MH3924A (1.07 ± 0.09; P = 0.016) compared with WT-MH3924A (0.65 ± 0.11), whereas the vascular fraction (VB) was not significantly different between wild-type (0.02 ± 0.01) and genetically modified (0.02 ± 0.01) tumors (Figs. 3 and 4).
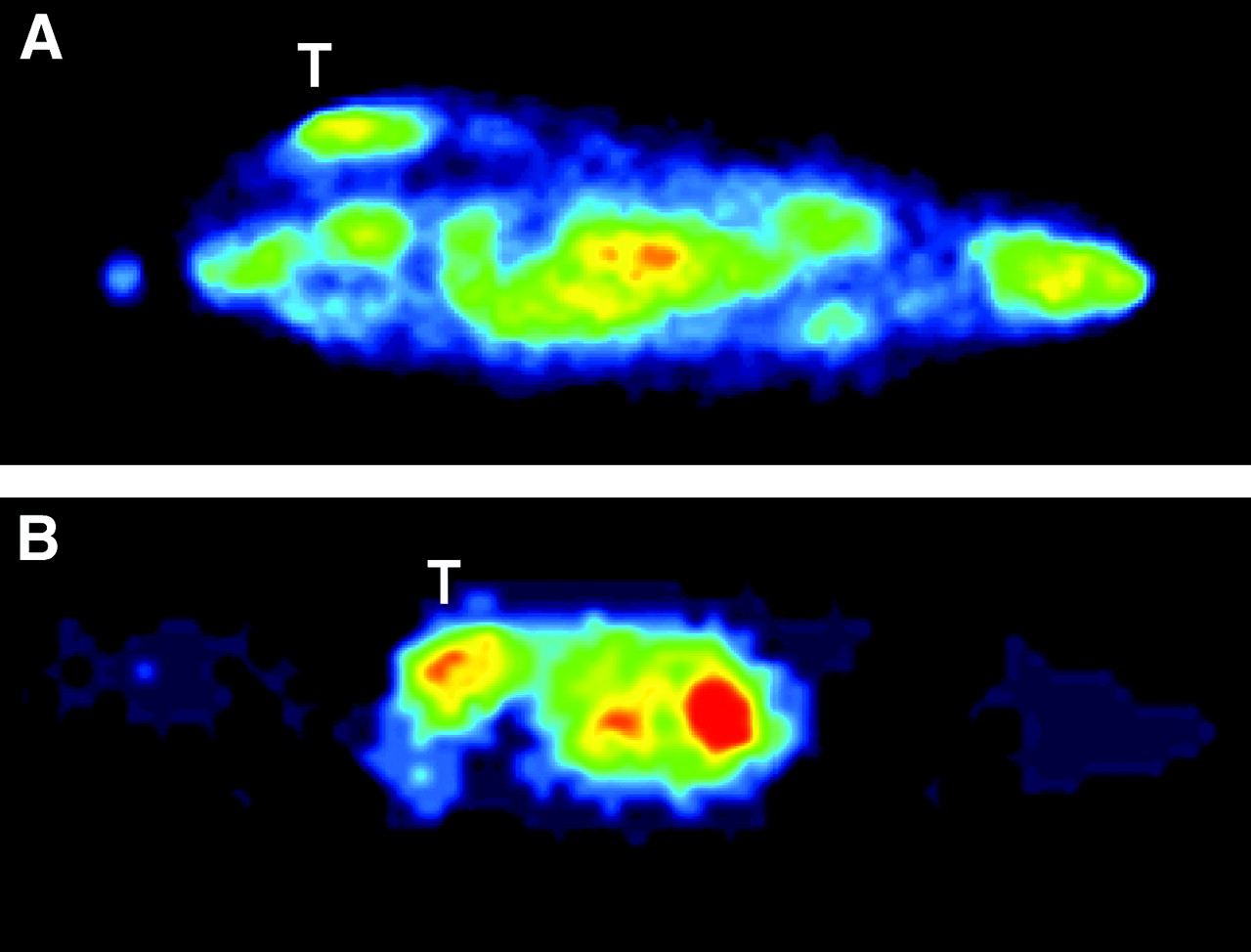
H215O PET images of rats with WT- (A) and angiopoietin-2−expressing hepatoma. T = tumor.

H215O PET. Tumor tissue perfusion (K1, k2, DV, and VB values) measured in WT-MH3924A (n = 6) and Ang-2-MH3924A (n = 6) tumors of ACI rats. Ang-2−overexpressing tumors demonstrate increased K1 (A), which was not significant with P = 0.06, whereas k2 was not changed in Ang-2-MH3924A (B). Fractional volume of distribution (DV) significantly increased in Ang-2-MH3924A (C), whereas vascular fraction (VB) remained unchanged (D).
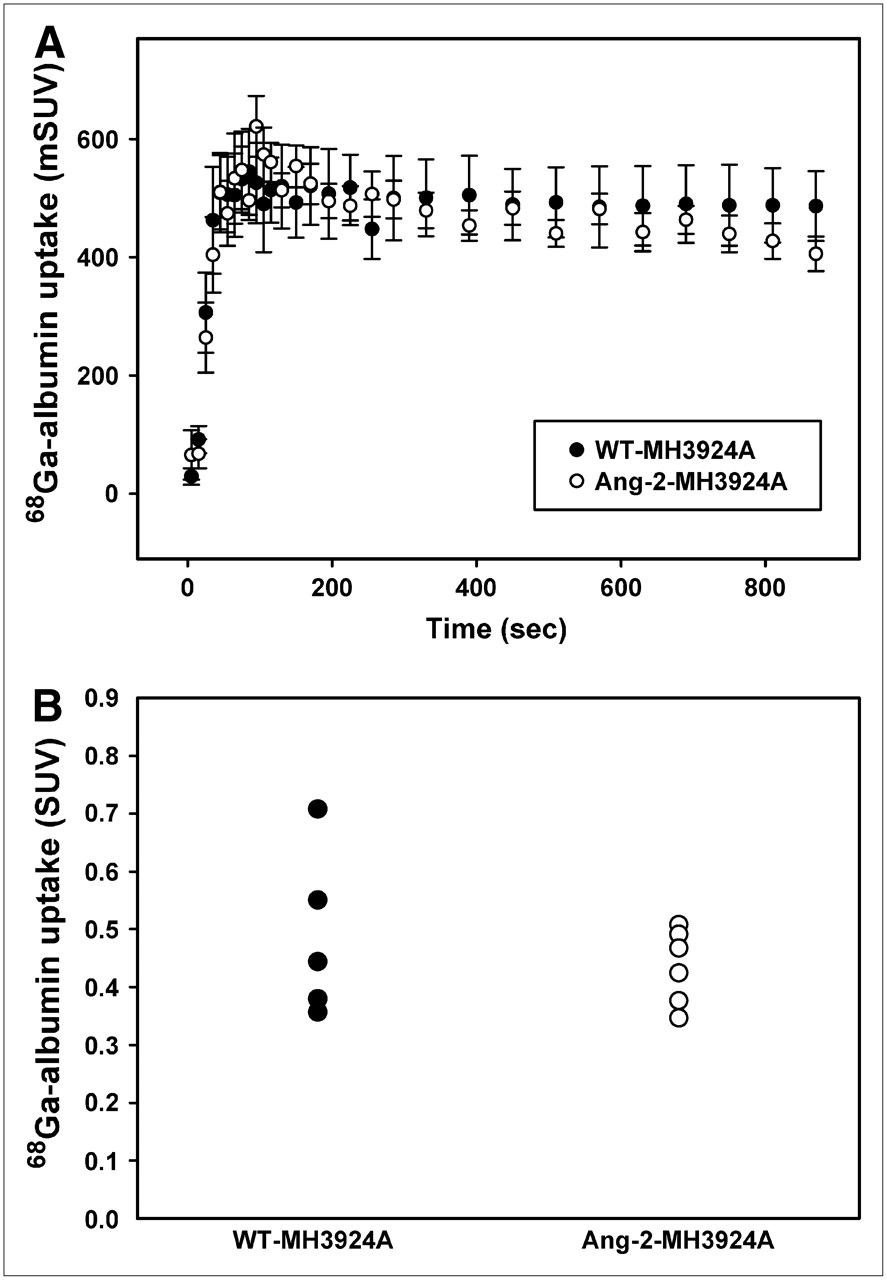
Because Ang-2 is commonly associated with immature, destabilized vessels, we performed PET measurements using 68Ga-DOTA-albumin to determine possible differences in blood pool between the Ang-2-MH3924A and WT-MH3924A tumors. No changes in albumin uptake were detected between the 2 groups (Fig. 5).
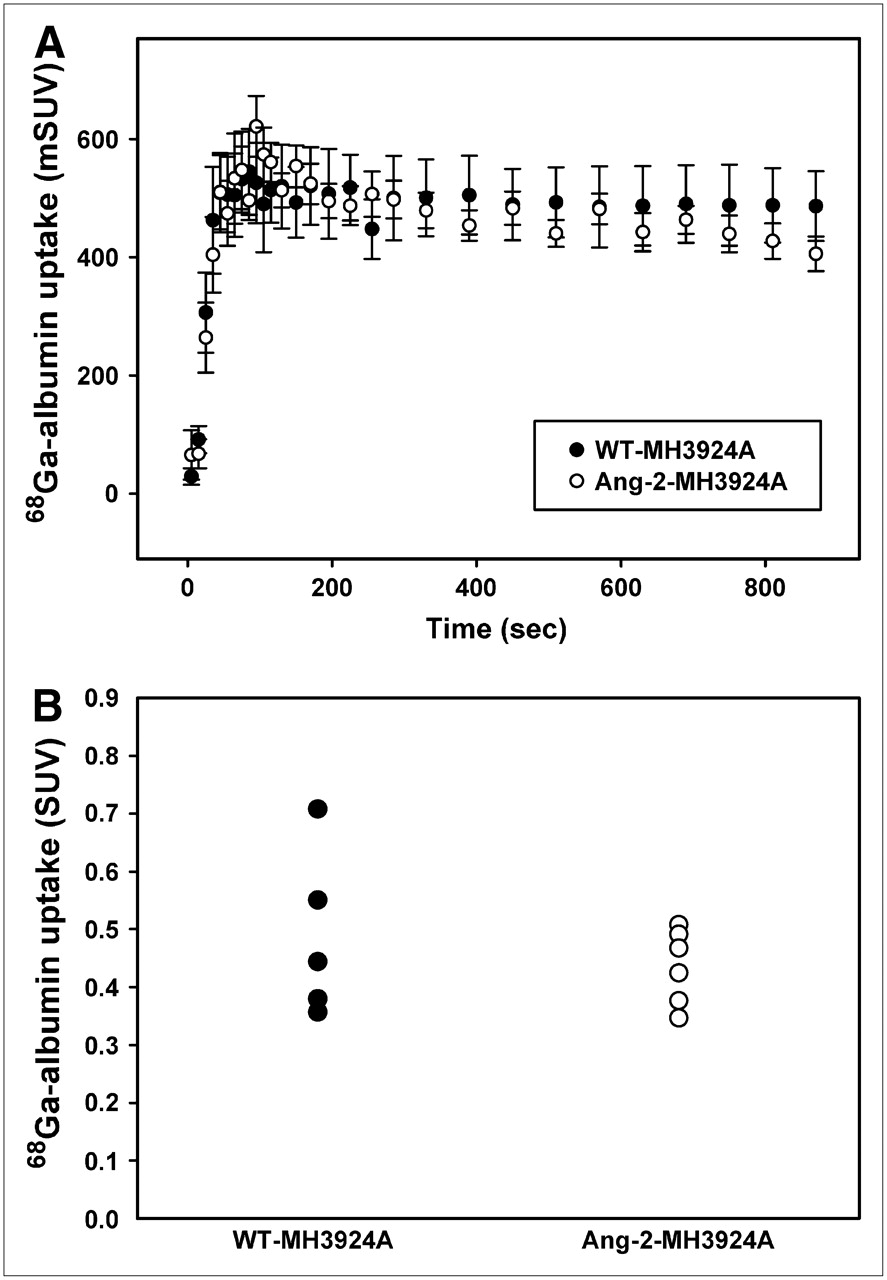
Time–activity curves for WT- and angiopoietin-2−expressing hepatomas (A). Blood pool within tumors, measured after 68Ga-DOTA-albumin application (representing permeability of vessels for albumin) showed no difference (B).
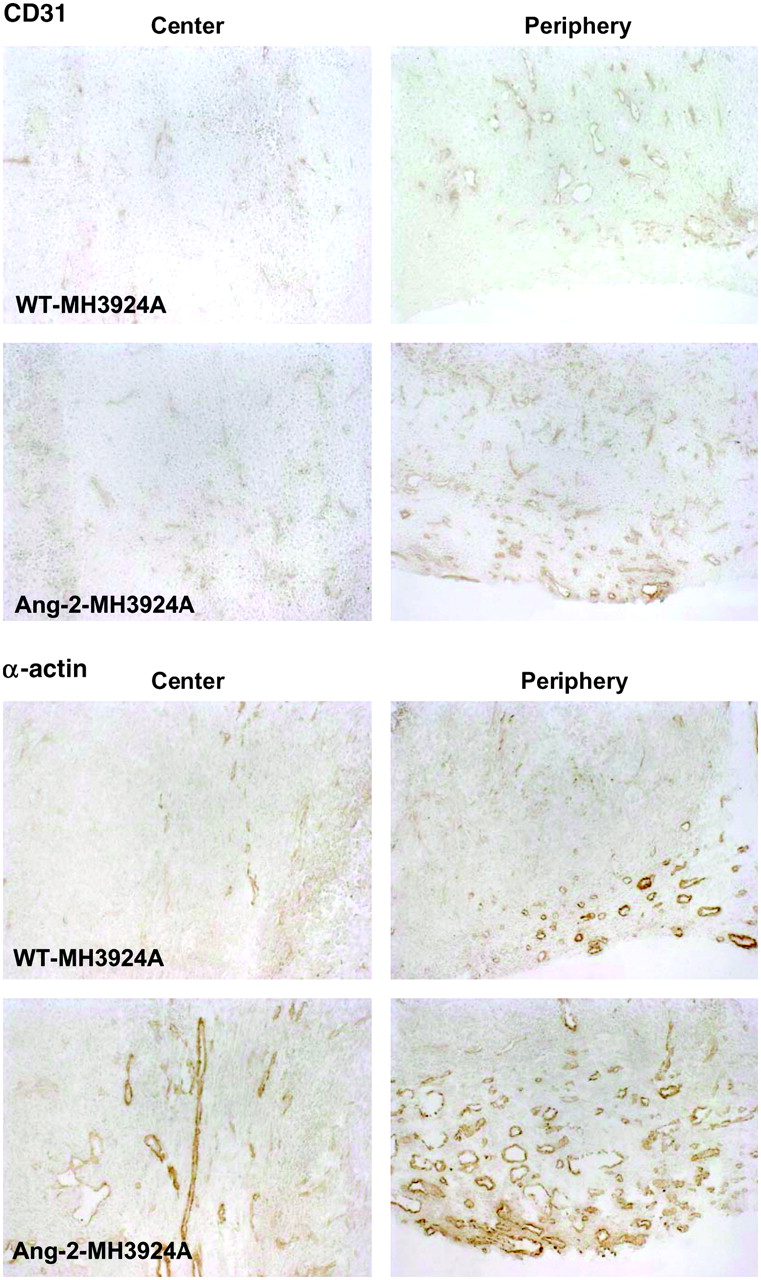
The evaluation of the anti-CD31 staining, a universal endothelial marker (arterioles, venules and capillaries), revealed a significant increase in vascularization in the periphery and the center of Ang-2-MH3924A tumors (Fig. 6 and Table 1; center: 2.3-fold, P = 0.003; periphery: 1.8-fold, P < 0.001). A commonly used vascular marker, α-actin, is expressed by mural cells of most arterioles and venules, but not in capillaries (10). In tumors of ACI rats, the macrovascularization, as indicated by the α-actin−immunoreactive area, was significantly increased in Ang-2−overexpressing tumors (Table 1; center: 2.5-fold, P = 0.006; periphery: 2.4-fold, P = 0.092).

Micro- and macrovascularization of Morris hepatoma identified by CD31 or α-actin staining. Representative pictures are shown. Note that Ang-2-MH3924A tumors revealed significant increase in microvessel density (as indicated by CD31 staining) compared with WT-MH3924A tumors, both in center and periphery of tumors as well as in macrovascularization (as indicated by α-actin staining). (×100)
Histochemical Analyses of Morris Hepatomas Transplanted into ACI Rats: Macrophages
Finally, we found no correlation between K1 and CD31, α-actin, or necrosis. In contrast, regression analyses revealed a strong positive correlation between the fractional volume of distribution (DV) and the percentage of CD31−immunoreactive area (r = 0.9), a weak correlation between DV and the percentage of α-actin−immunoreactive area (P = 0.64), as well as a weak negative correlation between DV and the area of necrosis (P = 0.65).
Ang-2 Overexpression: Effects on Proliferation, Apoptosis, Necrosis, and Markers of Oxidative Stress
Consistent with the higher vascularization and perfusion in the Ang-2 tumors, PCNA staining showed a significant increase of proliferating cells in the center (1.7-fold, P < 0.001) as well as in the periphery (3.5-fold, P < 0.001) of Ang-2-MH3924A tumors (Table 1). Besides the proliferative effect, Ang-2−overexpressing tumors also showed an increase of apoptotic cells in the periphery (1.7-fold, P = 0.035; Table 1), suggesting a high turnover in these tumors. In addition to the apoptotic effect, Ang-2 tumors revealed a strong reduction of necrotic area by approximately 90%, measured by hematoxylin and eosin staining and computer-assisted morphometry (WT: 20.67% ± 5.11%; Ang-2: 2.66% ± 0.71%, P = 0.003; Table 1).
The density of MIF-immunoreactive cells was significantly increased (85%) in the center of Ang-2-MH3924A tumors (P = 0.001; Table 1). Furthermore, the cluster area of CD11b−immunoreactive cells was significantly reduced by 71% (P = 0.027; Table 1).
Gene Transfer of Ang-2: Effects on Expression of Other Genes—Gene Arrays
To assess general effects of Ang-2 gene transfer on the tumor cells in vitro and in vivo, the gene expression pattern was studied using arrays. Because the transfer of selection markers such as the neomycin gene and the selection procedure may cause changes in the genetic program, we analyzed the expression pattern in the tumor cell lines as well in the tumors obtained after transplantation of these cell lines. In general, we found changes in the expression of genes related to cellular matrix, apoptosis, signal transduction and angiogenesis (Table 2).
Selected Genes Associated with Ang-2 Signaling: Ratio of Genetically Modified to WT Cells/Tumors
DISCUSSION
Ang-2−induced vessel formation is often described as leading to immature vessels sparsely coated by smooth muscle cells, pericytes, and CD34-positive endothelial precursor cells (5), suggesting leaky vessels rather than fully functional ones. Our results by dynamic PET measurements with H215O show that blood vessels, induced by overexpression of Ang-2, are of functional relevance, participating in tumor perfusion. Although the difference in K1 is not statistically significant and there is some overlap between Ang-2−expressing and WT tumors, the mean 1.5-fold increased perfusion (determined by H215O PET) in Ang-2-MH3924A in comparison with WT-MH3924A is in accordance with the increased vascularization (quantified by immunoreactive area for CD31 or α-actin) in Ang-2-MH3924A. The fractional volume of distribution (DV) significantly increased, possibly indicating changes in the membrane permeability. However, PET analysis conducted with 68Ga-DOTA-albumin, to determine the blood pool within the tumors, revealed no difference between the genetically modified and the WT-MH3924A tumors. The discrepancy between the values for DV and albumin accumulation may reflect the fact that the albumin data represent the volume defined by the intravascular space, whereas water evenly distributes in the total tumor volume.
Because of an increased perfusion and a significantly enhanced vascularization in Ang-2-MH3924A tumors, an increase of tumor growth might have been expected. However, between WT- and Ang-2-MH3924A, no difference in tumor volume was detectable. These results indicate that (a) increased perfusion/vascularization of tumors is not necessarily accompanied by enhanced growth and (b) plain anatomic imaging obviously does not reveal differences, which are induced by elevated angiogenic activity, whereas functional imaging seems to be more appropriate. Furthermore, our histomorphometric analyses revealed a significantly diminished necrotic area in Ang-2-MH3924A. The increase in proliferation rate as shown by PCNA staining may be compensated by an increase in apoptosis rate, leading to the observed similar growth of Ang-2−expressing and WT tumors. In addition, regression analyses revealed a positive correlation between DV and the percentage of CD31- (strong) or α-actin−immunoreactive area and a negative correlation between DV and the necrotic area. These data are best explained as a result of an improved functional vascularization and, consequently, an enhanced nutrition and oxygen supply of the tissue. Therefore, these alterations are more likely due to neovascularization than to direct effects on tumor cells.
Previous studies showed that Ang-2 is able to enhance proliferation of endothelial cells only in the presence of growth factors, but not in their absence, hypothesizing that growth factors modulate Ang-2 functions (5,11). In our experiments stimulation of HUVECs with a low concentration of bFGF (not being able to enhance the proliferation of HUVECs by itself) leads to a significant increase of proliferation in the presence of Ang-2. These data indicate that Ang-2 itself has the ability to modulate the effects of growth factors. Thus, Ang-2 might directly upregulate receptors of critical growth factors in angiogenesis, which may then affect downstream mechanisms.
Our CHIP data show that the VEGF receptors KDR and Flt-1, the FGF-R1, and the TNF-R are upregulated in Ang-2-MH3924A tumors. All of these receptors are involved in proliferation and migration of endothelial cells (12). KDR is known to be a key receptor of angiogenesis, by mediating proliferation, differentiation, and microvascular permeability (13). The assumption that the crucial step for the initiation of VEGF-induced angiogenesis is an upregulation of KDR, rather han an increase of VEGF itself (14), emphasizes the importance of this receptor in the initial phase of angiogenesis. It has been suggested by others that Ang-2 function is modulated by growth factors (11). However, our gene array data indicate that overexpression of Ang-2 induces an upregulation of growth factor receptors. In addition, the results of our in vitro proliferation assays with HUVECS are conclusive with the suggested mechanism of the interaction between Ang-2 and vascular growth factors. This possible interaction is also supported by data of several recent publications: Ang-2 has been shown to be no longer expressed in certain areas of the retina after day 7 after birth; in these areas, vessels were not sensitive to an application of VEGF, whereas VEGF application led to neovascularization in Ang-2−expressing areas (15). After induction of Ang-2 expression in these former VEGF insensitive areas, VEGF application resulted in neovascularization (15). These effects may be explained by the proposed mechanism of receptor induction by Ang-2 and show the ability of Ang-2 to sensitize blood vessels for growth factors. Furthermore, in a rat glioma model it has been reported that the initial neovascularization is regulated by a simultaneous upregulation of Ang-2 mRNA and KDR mRNA, whereas VEGF expression remained at a low transcription level, supporting the hypothesis that an increase of KDR rather than of VEGF itself is responsible for neovascularization (14).
MIF, a proinflammatory and carcinogenic cytokine, is related to the levels of biologic aggressiveness in hepatoma and colorectal tumors (16,17). Furthermore, MIF is suggested to act as an autocrine factor that stimulates angiogenesis and metastasis in hepatoma by promoting expression of angiogenic factors and migration of tumor cells (16). However, our data show that the percentage of MIF-positive cells is increased in the center of Ang-2-MH3924A tumors and the micro- or macrovascularization and apoptosis/proliferation rate are increased, whereas the necrotic area is diminished and the growth of these tumors is unchanged. These data indicate that immunohistochemically determined MIF may be linked to angiogenesis, perfusion/vascularization, and necrosis/apoptosis in hepatoma. Because MIF is a modulator of prooxidative stress-induced apoptosis (18) it may also function in this way in Ang-2-MH3924A.
CD11b(+) macrophages are suggested to be the major source of oxidative stress, and their cellular infiltration into the tissue was found to correspond to an enhanced tumor growth (19). However, our data show that a decrease of CD11b(+) macrophages in Ang-2-MH3924A is not necessarily linked with a reduction of tumor size, indicating that other factors play a major role in the regulation of tumor size. Because an increased growth of Ang-2−expressing tumors, due to an increased vasculature and blood flow, can be expected, the unchanged tumor growth might be due to the reduced percentage of CD11b(+) macrophages in these tumors.
Ang-2 promotes destabilization of vessels (2,20) by a loss of cell−cell and cell−matrix interactions as well as by modulating the basal lamina (11). As a consequence of the loss of these interactions, which mediate prosurvival signals on endothelial cells, Ang-2 leads to apoptosis in these cells; this apoptotic process occurs only if these prosurvival signals are not substituted by alternate prosurvival mediators such as VEGF or bFGF. Besides a significant increase in the percentage of apoptotic cells in Ang-2-MH3924A tumors, we found an upregulation in the expression of several proapoptotic factors such as Lice, caspase 6, tumor necrosis factor (TNF) receptor, and TNF-α converting enzyme. These results, in combination with the reduced necrotic area, indicate that proapoptotic/antiinflammatory mechanisms may prevail in Ang-2-MH3924A tumors. However, although proapoptotic/antiinflammatory processes are induced, growth is not inhibited in Ang-2-MH3924A tumors, probably due to the increased proliferation rate. Interestingly, besides their role in apoptotic processes, TNF-α and TNF receptor—which are upregulated in Ang-2 tumors (this study)—have been recently shown to be involved in angiogenesis (21). The proangiogenic effects of TNF-α and TNF receptor are described in interaction with Ang-2, being similar to the interaction between Ang-2 and growth factors: TNF's proangiogenic effect is enhanced in the presence of Ang-2—suitable to the observed upregulation of the TNF receptor (21).
Matrix-associated genes such as laminin, tropoelastin, SPARC, or tenascin are overexpressed in Ang-2-MH3924A. Several of these ECM-modulating and, in tumor invasion, involved proteins such as SPARC, SC1, tenascin, laminin, and osteopontin are reported to show similar effects on tumor invasiveness and metastasis as reported for Ang-2 (22–24). SPARC and SC1, a member of the SPARC-related family of matricellular proteins, are ECM-binding proteins, which induce tumor cell invasion in a glioma model (22). SPARC and Ang-2 have been suggested to act independently or synergistically in glioma progression. Osteopontin, a glycophosporylated protein interacting with integrin receptors such as αvβ3 and CD44, enhances the motility and invasive capacity of cancer cell lines (23) and reduces intercellular adhesion. Laminin and tenascins are major components of microvascular basement membranes, which facilitate tumor cell attachment to the ECM as well as migration and invasion of host tissue (24). Tenascins are found in invasive tumor sites, suggested to be involved in microvascular cell migration (25). Each of these ECM proteins is shown to be an important and potent mediator of tumor invasiveness and metastasis. Thus far, an association among these individual factors is not established, as well as their interactive mechanisms with Ang-2, which might be suggested as the connecting link for the induction of their expression.
It has been shown that the initiation of angiogenesis is highly dependent on the interaction of Ang-2 with endothelial growth factors (VEGF, b-FGF). Because the elementary signal for angiogenesis is a decrease in nutrition and oxygen supply, inducing VEGF via hypoxia-inducible factor 1α, VEGF, and familiar growth factors is the first response to this demand. Furthermore, VEGF directly upregulates Ang-2 production in endothelial cells via its receptor KDR (26), and TNF-α induces Ang-2- as well as Tie-2-receptor expression (27,28). Because only a few endothelial cells are KDR positive (14) in established vessels, the induction of Ang-2 probably results in only a weak response. The Ang−Tie2 system seems to have an autoregulatory feedback system, which regulates the overall activity of the Tie2 system (29). This auto-loop feedback mechanism may enhance the signal. Our results, showing that overexpression of Ang-2 coincides with the induced gene expression of the VEGF receptor KDR, as well as of other growth factor receptors such as bFGF-R1, have a logical and essential multiple role in this context: First, the postulated auto-loop feedback mechanism in endothelial cells could be explained; in a hypoxic environment, and therefore in the presence of hypoxia-regulated growth factors, the release of Ang-2 from the few KDR-positive endothelial cells may lead to an upregulation of KDR in the surrounding endothelial cells, resulting in an increased induction of Ang-2 via KDR on these formerly insensitive cells. Second, by inducing a loss of cell−cell and cell−matrix interaction, Ang-2 may diminish important survival signals in endothelial cells, mediated by these interactions. To prevent a shift toward apoptotic processes, it is a logical consequence that Ang-2 mediates an induction of substitutional prosurvival signals; by an upregulation of KDR, Flt-1, and bFGFR1, Ang-2 not only opens up alternative survival signals but also paves the way for proliferation and migration signals of the corresponding growth factors.
Although the clinical relevance of Ang-2 in antiangiogenic therapy still needs to be further elucidated, the existing results are very promising so far, showing that a blockade of Ang-2 leads to tumor regression and even a persistent disappearance after displacement of the treatment (30). The characteristics of Ang-2 make it an auspicious candidate for therapeutic issues: Ang-2 is only involved in few physiologic and pathologic situations, such as wound healing, ovarian cycle, tumor angiogenesis, or inflammatory processes. Therefore, an inhibition would lead to selective effects. By affecting multiple steps of angiogenesis, Ang-2 could disrupt the process on different levels. Focusing on its interaction with growth factors, a setting besides a blockade of Ang-2 would be an overexpression of Ang-2 in combination with a simultaneous blockade of growth factors, which is being studied currently by our group. Alternatively, a combination of Ang-2 overexpression with radiation therapy may increase the amount of oxygen radicals in the tumor, thereby potentiating the effects of radiation therapy.
CONCLUSION
Even though our results shed some light on potential cellular and molecular mechanisms of Ang-2, we are several steps away from understanding the complexity of its functions. It still remains unanswered whether Ang-2 mediates its effects through other pathways besides Tie-2. It seems unlikely that such diverse effects are mediated exclusively through an antagonism of Ang-1 at Tie-2. Because it has been recently shown that Ang-2 is capable of directly affecting glioma cells, another challenging issue remains—whether Ang-2 is able to directly influence other cell types involved in a tumorous environment. Because some postulated effects of Ang-2 are demonstrated to occur only in the presence of growth factors, the Ang-2−mediated induction of the receptor poses the question of whether these effects are actually direct effects of Ang-2 or, rather, effects mediated by the upregulated receptors. In this context, additional experiments must be done in the near future to answer this question.
Acknowledgments
The authors thank Uschi Schierbaum, Karin Leotta, Helmut Eskerski, and Ludwig Strauss for their help in performing the animal experiments and Silke Vorwald for expert technical assistance with immunohistochemistry. Financial support was from Wilhelm Sander Foundation (1999.085.1), Tumorzentrum Heidelberg/Mannheim, a bequest of Herbert Dauss, Foundation for Cancer and Scarlatina Research, and EMIL network.
Footnotes
-
↵* Contributed equally to this work.
-
COPYRIGHT © 2006 by the Society of Nuclear Medicine, Inc.
References
- Received for publication January 23, 2006.
- Accepted for publication May 8, 2006.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Ang-2-VEGF-A CrossMab, a Novel Bispecific Human IgG1 Antibody Blocking VEGF-A and Ang-2 Functions Simultaneously, Mediates Potent Antitumor, Antiangiogenic, and Antimetastatic Efficacy
- Targeting the Angiopoietin/Tie2 Pathway: Cutting Tumor Vessels With a Double-Edged Sword?
- Angiopoietin: A TIE(d) Balance in Tumor Angiogenesis
- Troponin I Overexpression Inhibits Tumor Growth, Perfusion, and Vascularization of Morris Hepatoma