


Abstract
Fatty acid oxidation (FAO) is the predominant energy-producing pathway in the healthy heart. Abnormalities in FAO are associated with many ischemic and nonischemic disease states. The aim of the present study was to further validate 16-[18F]-fluoro-4-thia-palmitate (18F-FTP) as a metabolically trapped FAO probe in the isolated perfused rat heart model by examining both the effects of hypoxia and the effects of changes in exogenous fatty acid availability. Methods: Hearts were excised from Sprague–Dawley rats and perfused in the Langendorff mode with Krebs–Henseleit solution under the following conditions: palmitate at 0.4 mmol/L with 95% oxygen, palmitate at 0.4 mmol/L with 35% oxygen, palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L with 95% oxygen, and palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L with 35% oxygen. Hearts were paced at 270 beats per minute, and the rate of left ventricular pressure change (LV dP/dt) was monitored. 18F-FTP in the perfusion medium was administered for 20 min, and this step was followed by a 20-min washout period without tracer in the perfusion medium. 18F kinetics in the whole heart were monitored externally, and the time–activity curves were analyzed to determine the fractional trapping rate for 18F-FTP (FTRFTP). A “lumped constant” (LC) was defined as the ratio of FTRFTP to the fractional rate of oxidation of fatty acid in the perfusion medium. Results: The kinetic data for 18F-FTP demonstrated metabolic trapping of 18F radioactivity that was insensitive to changes in the mixture of fatty acids in the perfusion medium but that was sensitive to the inhibition of mitochondrial FAO by hypoxia. LV dP/dt was reduced 47%–67% in hypoxic hearts relative to hearts with normal oxygenation (controls). FAO rates for palmitate and oleate were similar in group 3 (palmitate alone) and group 4 (palmitate and oleate). FAO was decreased 70%–76% with hypoxia, whereas FTRFTP was reduced 86%–88%, demonstrating hypersensitivity of a change in 18F-FTP retention to FAO inhibition by oxygen deprivation. The 18F-FTP LC was approximately 2 in myocardium with normal oxygenation and fell to 1.0–1.2 in hypoxic myocardium. Conclusion: The results confirm 18F-FTP to be a metabolically trapped palmitate analog that is capable of indicating rates of myocardial oxidation of exogenous long-chain fatty acids. The heterogeneous nature of fatty acids in plasma does not alter the quantitative analysis of 18F-FTP kinetics. However, the decreased LC value in hypoxic myocardium suggests the need to develop an understanding of the relationship of 18F-FTP processing to natural fatty acids at key limiting transport and metabolism processes, analogous to previous studies examining the LC values for radiolabeled deoxyglucose tracers used to estimate the glucose metabolic rate.
Although glucose, free fatty acids, lactate, pyruvate, and ketone bodies are consumed by the myocardium, the healthy heart derives 60%–80% of its energy requirements from fatty acid oxidation (FAO). There is a growing recognition of the importance of myocardial substrate metabolism—of FAO in particular—in the natural history of heart diseases, although many of the molecular mechanisms are not well understood. Marked abnormalities in FAO have been found to be associated with several cardiovascular diseases, including ischemic heart disease (1), heart failure (2), inherited disorders of FAO enzymes (3), and diabetes (4). Noninvasive assessment of myocardial FAO with PET and the development of an appropriate long-chain fatty acid tracer may provide a research and diagnostic tool for evaluating FAO in patients with cardiovascular diseases. Furthermore, an imaging method also would be of great value for monitoring disease progression and the effects of a growing number of drug therapies aimed at inhibiting FAO and promoting glucose uptake and oxidation (5–7).
11C-Labeled palmitate has been in research use for over 20 y as a PET tracer for fatty acid metabolism. FAO can be inferred from the early rapid clearance of radioactivity from the myocardium under aerobic conditions, but under ischemic conditions, the early phase is confounded by increased esterification and backdiffusion of nonoxidized radiotracer (8). Structurally modified fatty acid analogs with sulfur heteroatom substitutions have shown promise as FAO probes for addressing this issue (9,10). These probes do not suffer from the confounding effects of backdiffusion because FAO is inferred from the activity retained in the myocardium subsequent to metabolic trapping and because nonoxidative retention of radioactivity in lipid storage pools is very low.
Ebert et al. first developed 14-(R,S)-18F-fluoro-6-thia-heptadecanoic acid and showed that its uptake and retention in the myocardium track the changes expected with exercise and dipyridamole-induced hyperemia in healthy human subjects (11). The expected changes in 14-(R,S)-18F-fluoro-6-thia-heptadecanoic acid kinetics were seen with lactate infusion and hyperemia in extracorporeally perfused swine hearts (12). However, studies with rat and swine hearts showed 6-thia long-chain fatty acid probes to be insensitive to the inhibition of FAO by hypoxia (10,13). Subsequently, DeGrado et al. developed 16-[18F]fluoro-4-thia-palmitate (18F-FTP) and showed that moving the sulfur heteroatom to the carbon at position 4 allowed retention of the metabolic trapping character of the tracer while establishing proportionality of the tracer trapping rate and FAO under normal oxygenation and hypoxic conditions (10).
The present study was designed to further validate 18F-FTP kinetics in isolated rat hearts in response to hypoxia and changes in exogenous fatty acid composition. In addition, the relationship of 18F-FTP trapping in the myocardium to palmitate oxidation in the heart also was characterized. In this work, the 18F-FTP “lumped constant” (LC) is introduced by analogy to the LC used for estimation of the glucose metabolic rate with deoxyglucose analog radiotracers (14,15). An 18F-FTP LC is required for the quantitative estimation of regional rates of FAO from the PET-measured kinetics of 18F-FTP by relating the steady-state rate of oxidation of 18F-FTP to the rate of oxidation of exogenous nonesterified long-chain fatty acids. To provide meaningful conversions of the PET-measured rates of trapping of 18F-FTP to the FAO rates, the 18F-FTP LC should be predictable and consistent with respect to dependence on the metabolic rate, fatty acid substrate availability, and the influence of endogenous metabolic regulation (i.e., competing metabolic substrates). The presence of multiple fatty acid species in mammalian plasma makes the situation more complicated than that for 18F-FDG, which traces only glucose. 18F-FTP is presumed to be a palmitate analog that tracks changes in the palmitate oxidation rate. If there are significantly different oxidation rates for 18F-FTP and fatty acids other than palmitate, then changes in the composition of fatty acids in plasma could lead to erroneous estimates of FAO rates by use of a single LC based on the palmitate oxidation rate alone. To examine the importance of this potential problem, we used perfusion medium containing palmitate alone or an equimolar mixture of palmitate and oleate. These 2 fatty acids are the most prevalent nonesterified long-chain fatty acids in the plasma of rats and humans. 18F-FTP trapping rates were measured in relation to palmitate and oleate oxidation rates.
MATERIALS AND METHODS
Synthesis of 18F-FTP
18F-FTP was synthesized from cyclotron-produced no-carrier-added 18F by a previously described method (10). The radiochemical purity was greater than 99%, and the specific activity was greater than 7.4 × 1010 Bq/μmol. 18F-FTP was initially formulated in 1% albumin in isotonic saline, sterilely filtered, and then added directly to the perfusion medium for infusion into perfused rat hearts.
Isolated Rat Heart Perfusions
Sprague–Dawley (225–275 g) rats that had fasted overnight were anesthetized with pentobarbital (60 mg/kg) intraperitoneally. The hearts were excised and placed in ice-cold saline. After cannulation of the aorta, the hearts were perfused in retrograde fashion at a constant flow of 7 mL/min. The perfusion medium was Krebs–Henseleit bicarbonate solution with the following composition (mmol/L): Na+, 146; K+, 6.29; Ca2+, 2.40; Mg2+, 1.0; Cl−, 137; SO42, 1.18; H2PO4−, 1.36; HCO3−, 22.6; sodium acetate, 0.06; sodium pyruvate, 0.05; sodium lactate, 1.0; and glucose, 5. The medium also contained insulin at 4 mU/L and 2.67% albumin. The perfusate was filtered with 5-μm in-line filters (Millipore Corp.). The temperature was kept constant at 37°C. The hearts were paced at 270 beats per minute. A latex balloon was inserted into the left ventricle through the left atrium to obtain left ventricular (LV) pressure. LV pressure was recorded every 10 min with an analog-to-digital converter and computer logging software. The pressure traces were analyzed to determine the rate of left ventricular pressure change (LV dP/dt).
A total of 24 rats were randomly assigned to 1 of 4 experimental groups with 6 equally sized rats in each group (Table 1). To assess tracer kinetics under different oxygenation conditions, the solution was gassed with either 95% O2:5% CO2 for aerobic conditions or 60% N2:35% O2:5% CO2 for hypoxic conditions. To investigate the effects of differences in fatty acid composition in the perfusion medium, the hearts were perfused with either palmitate alone or a 1:1 molar mixture of palmitate and oleate. In all cases, the total fatty acid concentration in the perfusion medium was maintained at 0.4 mmol/L. [9,10-3H]Palmitate was infused to determine the palmitate oxidation rate from the release of tritiated water (2). For hearts perfused with oleate, [1-14C]oleate was present in the perfusion medium for the determination of oleate oxidation by measurement of released 14C-CO2.
Experimental Groups of Isolated Rat Hearts
After a 15-min stabilization period, the perfusion medium was changed to perfusate containing 18F-FTP at 3.5–5.0 GBq/L and 3H-palmitate at 1.9 MBq/L for a tracer “washin” period of 20 min. For hearts perfused with oleate, 14C-oleate at 1.9 MBq/L also was added. Effluent samples were collected at 10, 15, and 20 min. A 1-mL sample of the perfusate used during the 18F-FTP washin period was placed in a preweighed tube and weighed, and 18F radioactivity was determined with a well counter. After the tracer washin period, the perfusion medium was switched to medium that did not contain radiotracers for the study of 18F-FTP washout kinetics. At the end of perfusion, the hearts were weighed, and 18F radioactivity was determined with the same well counter.
Monitoring of 18F Radioactivity in Perfused Rat Hearts and Data Analysis
Two lead-collimated ∼5 × 5 cm (2 × 2 in.) NaI(Tl) scintillation probes were configured electronically for γ-coincidence detection of 18F radioactivity over the entire heart as previously described (10). The true coincidence counting rate in the heart (cps) was calculated as the difference between the prompt coincidence rate and the random coincidence rate, the latter of which was estimated by the delayed-signal technique. 18F radioactivity counts were recorded every second by computer and corrected for radioactive decay. Normalization of time–activity curves was performed by measurement of the apparent distribution volume (ADV) of 18F radioactivity in the heart at the end of perfusion with a well counter: Eq. 1The normalization factor that rescaled the entire time–activity curve was calculated as the quotient of the ADV from Equation 1 and the coincidence counting rate at the end of perfusion.
Eq. 1The normalization factor that rescaled the entire time–activity curve was calculated as the quotient of the ADV from Equation 1 and the coincidence counting rate at the end of perfusion.
The fractional trapping rate for 18F-FTP (FTRFTP) was determined from the slope of the ADV curves over the 10- to 20-min interval (Fig. 1) and rescaled to a per-gram-of-dry-weight basis: The linearity of the uptake curves after 10 min and the constancy of the washout curves after 10 min of washout (Fig. 1) support the assumptions that radioactivity in kinetically reversible pools (metabolic intermediates) reach steady state within 10 min of washin and that the clearance of metabolically trapped radioactivity is negligible over the 10- to 20-min uptake period. Because 18F-FTP is present in the washin perfusate at a constant concentration, the ADV curve during the washin period is equivalent to a Patlak–Gjedde graphic plot (16) that estimates the irreversible trapping rate for a tracer in tissue from the linear portion of the curve. A more complex analysis of the uptake data to determine individual kinetic rate constants for 18F-FTP transport and metabolism was deemed unnecessary in the present study.
The linearity of the uptake curves after 10 min and the constancy of the washout curves after 10 min of washout (Fig. 1) support the assumptions that radioactivity in kinetically reversible pools (metabolic intermediates) reach steady state within 10 min of washin and that the clearance of metabolically trapped radioactivity is negligible over the 10- to 20-min uptake period. Because 18F-FTP is present in the washin perfusate at a constant concentration, the ADV curve during the washin period is equivalent to a Patlak–Gjedde graphic plot (16) that estimates the irreversible trapping rate for a tracer in tissue from the linear portion of the curve. A more complex analysis of the uptake data to determine individual kinetic rate constants for 18F-FTP transport and metabolism was deemed unnecessary in the present study.

Representative time courses of 18F-FTP ADV in isolated rat hearts perfused under aerobic (95% O2) or hypoxic (35% O2) conditions. Perfusion medium contained palmitate at 0.4 mmol/L, glucose at 5 mmol/L, pyruvate at 0.05 mmol/L, lactate at 1.0 mmol/L, and insulin at 4 mU/L. Tracer was perfused at constant concentration over interval of 0−20 min. FTRFTP (mL/min/g) was determined from linear fit of data over interval of 10−20 min.
Measurement of Palmitate and Oleate Oxidation
In the effluent samples, tritiated water was separated from nonmetabolized 3H-palmitate by an organic solvent extraction procedure described by Saddik and Lopaschuk (17). The fractional palmitate oxidation rate (FORp [mL/min/g of dry sample]) was calculated as the product of the mass-specific perfusate flow (mL/min/g of dry sample) and the ratio of the effluent 3H2O concentration to the perfusate 3H-palmitate concentration. The palmitate oxidation rate (nmol/min/g dry) was calculated as the product of FORp and the palmitate concentration in the perfusate (nmol/mL).
To measure [1-14C]oleate oxidation, 14CO2 in the effluent samples was measured by acidification with 1N HCl within a sealed vial. The radioactive gas was trapped in a hanging bucket containing 1N NaOH in the same vial, and counts were determined. The recovery coefficient for 14C-CO2 was determined to be greater than 95% from measurements of mock samples treated with 14C-CO2. The fractional oleate oxidation rate (FORo [mL/min/g dry]) was calculated as the product of the mass-specific perfusate flow (mL/min/g dry) and the ratio of the effluent 14CO2 concentration to the perfusate 14C-oleate concentration. The oleate oxidation rate (nmol/min/g dry) was calculated as the product of FORo and the oleate concentration in the perfusate (nmol/mL).
The 18F-FTP LC values were calculated as the ratio of FTRFTP to FORp for hearts perfused with palmitate alone (groups 1 and 3) and FTRFTP to 0.5(FORp + FORo) for hearts perfused with equimolar concentrations of palmitate and oleate (groups 2 and 4).
Statistical Analyses
Data are expressed as mean ± SD. Weights, the FTRFTP, and the LC were analyzed with 2-way ANOVA models to investigate whether they were affected by 2 fixed effects, oxygenation of perfusion medium and fatty acid concentrations, and their interaction. Tukey post hoc tests were used to compare means if any of the effects were significantly different from the model. Mixed-effect models were used to investigate whether oxidation rates and LV were affected by 3 fixed effects, oxygenation of perfusion medium, fatty acid concentrations, and time, and their interactions; the subject identity was treated as a random effect. Means were compared by use of Tukey post hoc tests if the corresponding main or interaction effect was significant. Statistical analysis was performed with the SAS System for Windows, release 9.1/2003 (SAS Institute). P values of less than 0.05 were considered to be statistically significant in all tests.
RESULTS
Twenty-four rat hearts from 4 groups were studied according to oxygenation of the perfusion medium (95% = aerobic and 35% = hypoxic) and fatty acid concentrations (palmitate at 0.4 mmol/L or palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L) (Table 1). The wet and dry weights of the hearts were 0.99 ± 0.02 g and 0.14 ± 0.02 g, respectively. No difference in weights was found among the 4 groups (Table 2).
Wet and Dry Weights, FTRFTP, and LC in Isolated Rat Hearts*
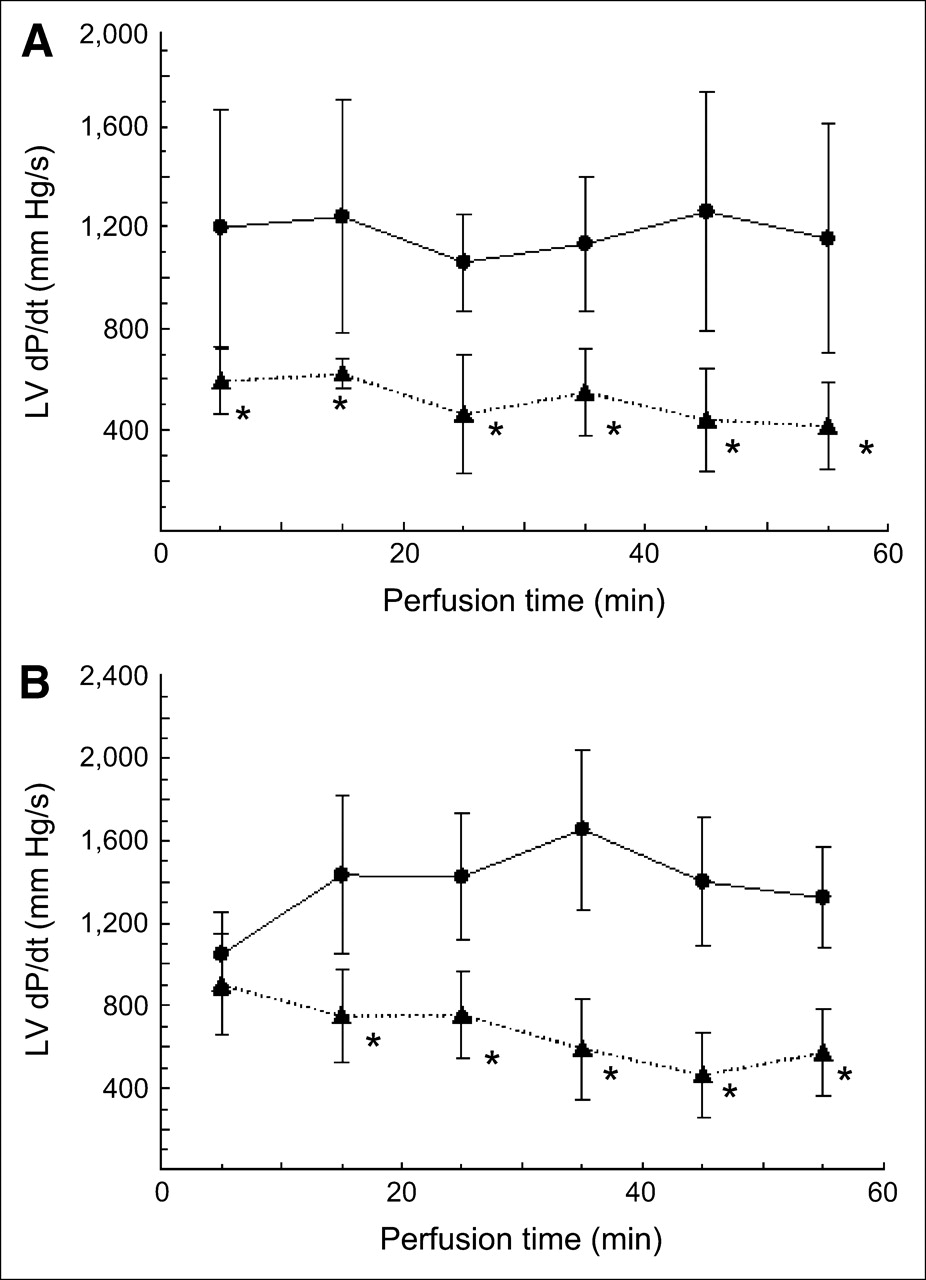
Figure 2 shows the effects of hypoxia on LV dP/dt. There was no statistically significant trend over time in the 4 groups. The LV dP/dt in hypoxic hearts was always lower than that in aerobic hearts at each time point from 5 to 55 min after perfusion of palmitate at 0.4 mmol/L, with an average decrease of 56%. A similar pattern was found with the perfusion of palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L, except that no difference was found at the 5-min time point and the average decrease (in LV dP/dt from aerobic to hypoxic) was 51%. Perfusion with palmitate at 0.4 mmol/L resulted in a lower LV dP/dt between 25 and 35 min than perfusion with palmitate at 0.2 mmol/L + oleate at 0.2 mmol/L under aerobic conditions. No other difference was found between the 2 concentrations at any other time point. No difference was found between the 2 concentrations under hypoxic conditions at any time point.

LV dP/dt in isolated rat hearts perfused with palmitate at 0.4 mmol/L (A) or palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L (B). Oxygenation of perfusion medium was aerobic (95% O2) (•) or hypoxic (35% O2) (▴). *P < 0.05 for hypoxic group versus aerobic group.
Rates of oxidation of exogenous fatty acids are shown in Table 3. Under all conditions, FAO rates were constant over the 10- to 20-min tracer infusion period. For aerobic hearts perfused with palmitate at 0.4 mmol/L, palmitate oxidation was 206.5–225.9 nmol/min/g dry. Perfusion under hypoxic conditions (35% O2) resulted in 68%–75% decreases in exogenous palmitate oxidation rates over the same interval (P < 0.001). For aerobic hearts perfused with equimolar concentrations of palmitate and oleate, the 2 fatty acids were oxidized at similar rates that were approximately one half the rate observed for hearts perfused with palmitate alone. Under anaerobic conditions, there was a trend toward higher rates of oleate oxidation than of palmitate oxidation (P = 0.11 at 10 min). The difference reached statistical significance at the 20-min time point.
Oxidation Rates for Exogenous Palmitate and Oleate in Isolated Rat Hearts as Function of Fatty Acid Composition and Oxygenation of Perfusate*
Figure 1 shows representative time–activity curves of 18F-FTP for hearts perfused with palmitate at 0.4 mmol/L under normal oxygenation and hypoxic conditions. Linearity of uptake was achieved under both conditions within 5 min of perfusion with the tracer at a constant concentration in the perfusion medium. This process allowed the estimation of FTRFTP to be derived by a linear fit of the time–activity curves over the interval of 10–20 min. Similar linearity was found for hearts perfused with a mixture of fatty acids (data not shown). When the perfusion medium was switched to a medium devoid of 18F-FTP tracer, there was an initial rapid clearance of radioactivity from the heart, likely representing washout of radioactivity from vascular and interstitial spaces, followed by 1 or 2 slower tissue clearance processes. The apparent half-lives of the slowest clearance processes were on the order of several hours in all experimental groups, suggesting that 18F-FTP is effectively metabolically retained in the myocardium. The amount of radioactivity retained in hypoxic myocardium at the end of perfusion was reduced markedly, consistent with a lower level of metabolic trapping of 18F-FTP during the constant-infusion period.
FTRFTP and LC data are shown in Table 2. As expected, hypoxia caused a dramatic reduction in FTRFTP. The mean FTRFTP values were only 14% and 12% the aerobic values with perfusion of palmitate at 0.4 mmol/L and perfusion of palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L, respectively (P < 0.05). FTRFTP was not affected by fatty acid composition in the perfusion medium under hypoxic conditions. However, the mean FTRFTP value with palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L was higher than that with palmitate at 0.4 mmol/L under aerobic conditions (Table 2). The LC was 2.09 ± 0.96 for aerobic hearts perfused with palmitate at 0.4 mmol/L. The LC was 2.15 ± 0.97 for aerobic hearts perfused with palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L. The LC values under aerobic conditions were approximately 2-fold higher than those for hypoxic hearts, consistent with the finding that FTRFTP was reduced more than the FAO rate in hypoxic myocardium. However, the LC was not affected by fatty acid composition under either oxygenation condition (Table 2).
DISCUSSION
In the healthy heart, FAO is the primary source of oxidized substrates for adenosine triphosphate production. However, FAO is altered in almost all myocardial disease states (1–4), and the use of nonlipid substrates is often not sufficient to maintain the adenosine triphosphate levels needed to support normal contractile function. Decreased FAO also can be responsible for increased levels of deleterious fatty acyl intermediates that may cause structural and functional abnormalities within myocytes (18–20). Consequently, FAO is an important biochemical pathway that has been targeted for both diagnostic and therapeutic applications in the management of patients with cardiovascular diseases. For this reason, medical imaging methods with many different radiolabeled fatty acid analogs have been explored with the goal of the noninvasive assessment of FAO in patients.
Because radioisotopic imaging methods such as PET cannot differentiate the signals of 2 or more metabolic entities in tissues, the methodology must properly differentiate oxidative metabolism and nonoxidative metabolism of fatty acid tracers by other means. One approach to this problem has been to use a direct analog tracer, such as [1-11C]palmitate, and to exploit tracer kinetic information in the tissue time courses to differentiate between the characteristically more rapid oxidation processes that release radiolabeled catabolites and the slower turnover of radioactivity in lipid storage pools. This approach was applied quantitatively by Bergmann et al. (21) through the use of a compartmental model to estimate oxidative flux and nonoxidative flux of 11C-palmitate. However, this approach cannot be applied to ischemic myocardium because the early rapid clearance phase is confounded by increased backdiffusion of nonoxidized tracer from the myocardium (8). If the tracer is able to be incorporated into endogenous lipid storage pools, then the tissue signal will reflect rates of uptake of exogenous fatty acids by the heart and esterification to form acyl coenzyme A (acyl-CoA) but will not be useful for indicating the flux of acyl-CoA toward β-oxidation. An example of such a tracer is 123I-β-methyl-p-iodophenylpentadecanoic acid (BMIPP), which can be used in conjunction with single-photon imaging techniques to evaluate fatty acid uptake by the myocardium. BMIPP has been studied extensively in patients with ischemic heart disease (22), cardiomyopathy (23), and heart failure (24). However, BMIPP kinetics cannot be used to evaluate FAO; experimental inhibition of carnitine palmitoyltransferase 1 was found to have no effect on BMIPP uptake in isolated rat hearts (25).
Results from various laboratories (9,10,26) have found that sulfur heteroatom substitution of long-chain fatty acid analogs renders thia fatty acid analogs poor substrates for lipid incorporation, although they are still capable of initial transport and oxidative metabolism in the mitochondrial FAO pathway. Substitution of sulfur at the carbon at position 3 results in poorly retained radiolabeled metabolites, whereas substitution at the carbon at position 4 or 6 results in radiolabeled metabolites that are well retained by the myocardium (9,10,26,27). These results are in keeping with the known biochemical processing of odd-carbon– and even-carbon–substituted thia fatty acids. Even-carbon–substituted fatty acids are β-oxidized to their respective 4-thia-3-hydroxy-CoA esters in the mitochondrion, which spontaneously fragments to form malonyl-CoA semialdehyde and a long-chain thiol (27). Long-chain thiols are highly reactive with cellular proteins and are retained in myocytes. The trapping of a metabolic fragment in the myocardium forms the basis of the retention mechanism for terminally labeled even-carbon–substituted long-chain thia fatty acid analogs. However, to be useful for the quantitative assessment of FAO in living subjects, the accumulation of radioactivity in the myocardium should have a well-defined relationship to FAO rates over a wide range of FAO rates and under various clinically relevant conditions.
An important and potentially problematic issue that must be addressed in the quantitative analysis of FAO with 18F-FTP PET is the influence of the heterogeneity of available plasma nonesterified fatty acids on 18F-FTP kinetics and estimates of FAO. Plasma contains multiple fatty acid species that differ in chain length and the degree of unsaturation. Oleate (18:1) and palmitate (16:0) are the most common fatty acids in human plasma and together account for approximately 70% of the total nonesterified fatty acids (28,29). The predominance of oleate and palmitate also is seen in the plasma of dogs (28) and rats (30). On the basis of chain length and degree of saturation, 18F-FTP is closest in structure to palmitate. The ability of 18F-FTP to trace the oxidation of total nonesterified fatty acids in plasma depends on the close relationship between the 18F-FTP trapping rate and the palmitate oxidation rate and, in turn, between the palmitate oxidation rate and the rates of oxidation of oleate and other fatty acids in plasma. In the present study, we showed that a change in the composition of the fatty acids in the perfusion medium from palmitate at 0.4 mmol/L to palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L resulted in no significant change in the total FAO of exogenous fatty acids. In the groups perfused with both fatty acids, there was no consistent preference for the oxidation of one fatty acid over the other, but a trend was observed for higher relative rates of oxidation of palmitate under aerobic conditions, reversing to higher relative rates of oxidation of oleate under hypoxic conditions. However, only under hypoxic conditions at the 20-min time point did this trend become statistically significant. It is interesting that the trapping rate for the palmitate analog 18F-FTP followed this trend in that it was found to be slightly (23%) higher in the group perfused with palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L than in the group perfused with palmitate at 0.4 mmol/L, but only under aerobic perfusion conditions. A possible explanation for this discrepancy is that 18F-FTP trapping requires only the first 2 metabolic steps of β-oxidation to occur, whereas the measurement of oxidation for [9,10-3H]palmitate and [1-14C]oleate requires more complete β-oxidation and the release of radiolabeled catabolites from tissues. Thus, differences in the metabolic processing of 18F-FTP through upstream processes may be accentuated in the FTRFTP metric relative to the signals obtained for the oxidation of [9,10-3H]palmitate and [1-14C]oleate. These differences may be realized only under aerobic conditions, in which upstream processes (i.e., transport) are more rate limiting to the overall FAO rate. Notwithstanding these observations, our results suggest that at least in rats, the heterogeneity of plasma fatty acids does not significantly complicate the relationship between tracer use and the total FAO rate.
These data are corroborated by in vivo studies in humans by Wisneski et al. (31), who showed that individual FAO rates for palmitate and oleate normalized to their concentrations in plasma were similar to each other in healthy subjects. In patients with coronary artery disease (CAD), the levels of palmitate, oleate, and total nonesterified free fatty acids in plasma were increased. FAO rates relative to plasma concentrations were decreased in patients with CAD, but the decreases were similar for both palmitate and oleate (31). However, somewhat higher myocardial extraction fractions of oleate relative to palmitate have been reported for humans (28) and dogs (21,28), but FAO rates were not measured in these studies. Bergmann et al. (21), using an experimental dog model, found that the amount of palmitate as a percentage of total nonesterified fatty acids in arterial plasma was constant and that the steady-state extraction fractions of individual fatty acids were similar in control animals and in several interventional groups, including groups with glucose infusion, dobutamine stress, and FAO inhibition. Overall, these data suggest that a palmitate tracer, if found useful for estimating palmitate oxidation rates, also may serve as a prototype fatty acid for the quantitative assessment of the total FAO rate in the myocardium.
A second potential pitfall in developing a quantitative 18F-FTP PET technique pertains to the accuracy of the LC value that is used to convert myocardial tissue trapping rates to estimates of regional FAO rates. Ideally, the LC should be stable over a wide range of FAO rates and should be independent of substrate availability. With respect to their relationship to FAO, we found the LC values to be approximately 2.1 in normally oxygenated perfused hearts and 1–1.2 in hypoxic perfused hearts. The LC was insensitive to the change in the fatty acid composition in the perfusate from palmitate at 0.4 mmol/L to palmitate at 0.2 mmol/L plus oleate at 0.2 mmol/L. Underlying the 50% decrease in the LC values with hypoxia is the fact that the decreases in 18F-FTP trapping rates in hypoxic myocardium are nearly 2-fold larger than the decreases in FAO rates. There are several possible reasons for the reduced LC values seen under hypoxic conditions. Further experiments with a more extensive set of conditions are needed to provide further clarification of this parameter. Nevertheless, a plausible explanation is that 18F-FTP and palmitate (or oleate) have different affinities for transporters and enzymes along the FAO pathway and that the inhibition of FAO by hypoxia enhances the strength of control of downstream processes over early transport processes. Under normal oxygenation conditions, FAO likely is limited to a greater extent by upstream transport processes. Under hypoxic conditions, however, the mitochondrial β-oxidation rate becomes rate limiting, and transport and early metabolic steps will exert less control. This scenario is plausible only if the relative flux of 18F-FTP through the early transport processes is greater than the flux of palmitate (or oleate), hence leading to an LC value of greater than 1, whereas the relative affinity of 18F-FTP through the oxidation-dependent steps is substantially lower than that of natural fatty acids. Such tracer behavior is analogous to that of 18F-FDG with respect to its relationship to glucose phosphorylation rates. Under conditions that favor glucose transport control (for example, low plasma glucose concentration plus insulin) of glucose phosphorylation, the LC value for 18F-FDG is greater than 1 in isolated rat hearts, consistent with the known higher enzyme efficiency for 18F-FDG than for glucose at the myocardial glucose transporter (15,32). However, under conditions that favor hexokinase activity as the rate-determining step (for example, high plasma glucose concentration plus insulin), the 18F-FDG LC value is decreased substantially, consistent with the increased influence of substantially poorer enzyme efficiency on the phosphorylation of FDG relative to glucose by hexokinase (15,32).
On the basis of the work of Kuwabara et al. (33), Botker et al. (32) demonstrated that the value of the myocardial 18F-FDG LC may be estimated in each experimental subject from regressed 18F-FDG transport and phosphorylation rate constants. Such an approach also could be pursued for estimation of the 18F-FTP LC, although the larger number of metabolic steps for fatty acids may prevent the creation of a comprehensive mathematic model. To provide a stronger basis for understanding the quantitative aspects of 18F-FTP trapping, future work should define the important limiting transport and metabolic steps, the relative affinity of 18F-FTP at these steps, and the influence of physiologic changes and pathologies on these steps.
A limitation of the present study was that hypoxia was studied without a change in perfusion. Ischemia may influence 18F-FTP metabolism or retention kinetics through differences in FAO intermediate levels and reduced clearance of radiolabeled intermediates or catabolites. Ischemia also will reduce the delivery of tracer to the myocardium. A second limitation of the present study was the use of a single level of hypoxia. It will be of interest to study 18F-FTP kinetics over a wide range of FAO levels and different oxygenation levels.
An 18F-labeled probe for the assessment of myocardial FAO is anticipated to have relevance for the clinical assessment of patients with various diseases, including ischemic heart disease, diabetic cardiomyopathy, and heart failure. Advances in PET/CT scanners in recent years have opened new possibilities for PET/CT to become a one-step technique for the simultaneous assessment of coronary anatomy and regional biochemical function (34,35). There is promise that such studies could greatly reduce the number and length of diagnostic imaging procedures that a typical cardiac patient experiences.
The biochemical and tracer kinetic properties of 18F-FTP make it an ideal candidate for PET/CT, particularly for the evaluation of ischemic heart disease. The present observations of 18F-FTP kinetics in isolated rat hearts further validate 18F-FTP as a metabolic PET probe that is capable of indicating FAO in the heart. The insensitivity of 18F-FTP trapping rates to changes in palmitate and oleate mixtures in the perfusion medium (at a constant total fatty acid concentration) suggests that myocardial trapping of 18F-FTP may serve as a prototype fatty acid tracer for indicating rates of oxidation of total plasma fatty acids available to the myocardium. Because FAO is indicated by the retention of the metabolically trapped tracer, the backdiffusion of 18F-FTP does not confound the interpretation of images, but sufficient time is necessary to distinguish the metabolically trapped fraction. In rat hearts, 10–15 min is adequate for the washout of untrapped 18F-FTP. Previous studies with bolus injections of 18F-FTP in swine confirmed the stable myocardial kinetics at 15 min after injection (10). The stable myocardial kinetics should allow for the acquisition of PET data with cardiac gating for the mitigation of cardiac motion artifacts, evaluation of the ejection fraction, and the potential assessment of LV wall motion abnormalities.
On the basis of previous experience with 18F-FDG and other 18F-substituted analog tracers that exhibit different relative affinities at different steps of transport or metabolism, it is not surprising that the 18F-FTP LC values are not constant over a range of FAO rates. The finding of an LC value for 18F-FTP of approximately 2 under aerobic conditions suggests that 18F-FTP trapping will provide an excellent signal for the evaluation of FAO by the heart. Indeed, preliminary imaging studies with swine (10) and humans (R.E. Coleman and T.R. DeGrado, unpublished data, June 2000 to December 2001) have demonstrated that 18F-FTP provides high ratios of myocardium activity to blood and lung background activities. In the present study, the inhibition of mitochondrial β-oxidation by oxygen deprivation led to a substantial decrease in the 18F-FTP LC values. If the 18F-FTP LC values behave in a similar manner in ischemic myocardium, then 18F-FTP PET images of ischemic myocardium may be hypersensitive to decreases in FAO, and careful attention should be given to interpreting quantitative assessments of tracer retention, particularly in potentially ischemic regions of the heart. In the context of qualitative PET image interpretation, however, the hypersensitivity of 18F-FTP to mitochondrial FAO inhibition may allow the identification of moderately ischemic myocardium as regions of 18F-FTP retention deficits. Further work is needed to characterize 18F-FTP retention at different levels of ischemia, including both perfusion-limited oxygen deprivation and demand-induced ischemia.
An additional concern for patients with CAD is that plasma fatty acid levels commonly are elevated and could affect the 18F-FTP LC. The present study suggests that the LC values may increase under such conditions because the strength of control of the transport step is expected to increase as the FAO rate limitation is shifted toward fatty acid transport.
CONCLUSION
The results of the present study confirm that 18F-FTP is a metabolically trapped palmitate analog that is capable of indicating rates of myocardial oxidation of exogenous long-chain fatty acids. A formalism with an LC conversion was introduced to relate 18F-FTP trapping rates in the myocardium to FAO rates by direct analogy to the use of an LC for estimation of the glucose metabolic rate with deoxyglucose radiotracers. An 18F-FTP LC value of approximately 2 was found for the indication of palmitate (or oleate) oxidation in aerobically (95% oxygen) perfused isolated rat hearts. Under conditions of hypoxia (35% oxygen), the 18F-FTP LC value decreased to approximately 1. The 18F-FTP LC was not sensitive to changes in the mixture of palmitate and oleate at a constant total fatty acid concentration. It was concluded that 18F-FTP retention in the myocardium is a sensitive indicator of FAO. However, further work is needed to understand 18F-FTP processing in relation to that of natural fatty acids at key limiting steps of transport and metabolism and to develop appropriate models for the quantitative estimation of FAO from imaging data.
Acknowledgments
This work was supported by National Institutes of Health grants HL-63371 and CA-108620.
References
- Received for publication August 16, 2005.
- Accepted for publication October 18, 2005.





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Organ-Specific Fuel Rewiring in Acute and Chronic Hypoxia Redistributes Glucose and Fatty Acid Metabolism
- Adipose Tissue Lipolysis Promotes Exercise-induced Cardiac Hypertrophy Involving the Lipokine C16:1n7-Palmitoleate
- Synthesis and Preliminary Evaluation of 18-18F-Fluoro-4-Thia-Oleate as a PET Probe of Fatty Acid Oxidation
- Radionuclide Imaging of Myocardial Metabolism