


Abstract
Mesothelin is a glycoprotein that is overexpressed in several human tumors, including mesotheliomas and ovarian cancers, and has been identified as a potential target for therapy. We evaluated the biodistribution and tumor-targeting ability of an antimesothelin tetravalent single-chain Fv-streptavidin fusion protein (SS1scFvSA) in mice. Methods: SS1scFvSA was labeled with 125I or 111In for evaluation of internalization in vitro and for optimization of its biodistribution. The A431-K5 mesothelin transfected cell line was used as the target. We used a 3-step pretargeting approach consisting of injections of (i) SS1scFvSA, followed 20 h later by (ii) a synthetic clearing agent, and (iii) 4 h later, radiolabeled (111In, 88Y/90Y, or 177Lu) 1,4,7,10-tetraazacyclododecane-N,N′,N″,N‴-tetraacetic acid (DOTA)-biotin. To optimize the tumor uptake, the effect of the specific activity of 111In-DOTA-biotin was evaluated. Results: Approximately 60% of SS1sc FvSA internalized within 6 h. The optimal dose of SS1scFvSA for pretargeting was 600 μg. Decreasing the specific activity of DOTA-biotin by administering 0.1–5 μg of DOTA-biotin resulted in tumor uptake decreasing from 31.8 to 5.5 %ID/g (percentage injected dose per gram) at 2 h. Pretargeted therapy of A431-K5 tumor with 90Y doses of 11.1–32.4 MBq resulted in a dose-dependent tumor response. With 32.4 MBq, 86% of mice survived tumor free for 110 d. All nontreated mice died, with a median survival of 16 d. Conclusion: SS1scFvSA localized in the mesothelin-expressing tumor, resulting in a high accumulation of radiolabeled DOTA-biotin. The specific activity of DOTA-biotin had a significant effect on its tumor uptake. Therapeutic tumor doses were obtained without dose-limiting toxicity.
Radioimmunotherapy (RIT) of nonhematopoietic solid tumors has shown limited success compared with that of lymphomas (1) Various pretargeting, multistep approaches have been proposed to address the limitations of RIT of solid cancers (2–4), which have in common injection steps that separate the tumor-targeting step with antibody from the delivery of the radioactivity to the tumor. In general, these approaches have relied on avidin or streptavidin (SA) interaction with biotin or on bifunctional antibodies that recognize tumor antigen (Ag) with one binding site and a chelated isotope with the other binding site.
A 3-step pretargeting approach has been developed by NeoRx Corp. (4,5). This approach consists of (i) delivery of the SA-conjugated antibody to target the tumor; (ii) injection of a clearing agent that eliminates the remaining circulating antibody-SA conjugate from the blood into the liver, where it is no longer available for binding; and (iii) injection of radiolabeled biotin that distributes quickly into the tissues where it binds to the antibody-SA conjugate or the unbound fraction is cleared quickly into the urine. Several studies have demonstrated that this approach can result in rapid tumor targeting, high tumor-to-nontumor ratios, and successful RIT (5–10). These experimental studies have evaluated a variety of antibodies and a variety of radioisotopes. Furthermore, proof of principle has been shown in clinical trials (4,6,11).
Recently, mesothelin has emerged as a new target for an antibody-based therapy (12,13). Mesothelin is a 40-kDa glycoprotein differentiation Ag that is not expressed in most normal tissues, except the pleura, pericardium, and peritoneum, which are lined with mesothelial cells (13). Since mesothelin is overexpressed in a variety of malignancies, it is a good target for antimesothelin antibody–based therapies (13,14). SSscFv, a monoclonal antibody (mAb) fragment that binds to mesothelin, was generated by immunizing mice with an eukaryotic expression vector coding for mesothelin (15). Pseudomonas immunotoxins based on SSscFv have been generated and one, SS1(dsFv)PE38, has been used in a clinical trial (16,17).
In this study, we optimized the pretargeting approach using a tetravalent single-chain Fv-streptavidin (SS1scFvSA) fusion protein that recognizes mesothelin. In addition to biodistribution studies with various radionuclides, we conducted a successful RIT with 90Y-1,4,7,10-tetraazacyclododecane-N,N′,N″,N‴-tetraacetic acid (DOTA)-biotin after pretargeting with SS1scFvSA.
MATERIALS AND METHODS
Mesothelin Antibodies
The SS1scFvSA (molecular weight, 172,928) was expressed as a genetic fusion of the SS1 single-chain variable region (scFv) to the full-length genomic SA by NeoRx Corp. using methods previously described (6,18). SS1scFvSA has 4 Fv fragments per molecule and a single SA with its 4 binding sites for biotin. Anti-Tac scFvSA fusion protein that was created using the same technology (9) was used as a mesothelin nonspecific control in therapy studies. It recognizes the α-subunit of interleukin 2 receptor. To confirm the tetrameric composition of SS1scFvSA, it was analyzed on 4%–20% tris-glycine sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) with and without reducing. The gel stained with Coomassie Blue revealed a major band at ∼170 kDa without reducing and an ∼43-kDa band when reduced, indicating that the tetrameric fusion protein was divided to 4 fragments on reduction.
Radiolabeling
To determine the immunoreactivity and the biodistribution of the SS1scFvSA, we labeled SS1scFvSA with 125I (Perkin-Elmer Life Science-NEN) at a specific activity of 2.4–3.7 kBq/μg using the IODO-GEN (Pierce Chemical Co.) method (19) or with 111In (Perkin-Elmer Life Science-NEN) at a specific activity of 307–740 kBq/μg using the 2-(p-isothiocyanatobenzyl)cyclohexyl-diethylenetriaminepentaacetic acid (CHX-A″) chelate conjugated to the fusion protein (20). For pretargeting studies, biotinidase-resistant DOTA-biotin (21) was labeled with 111In, 88Y (Oak Ridge National Laboratory), or 177Lu (International Isotopes Inc.) at a specific activity of 3.7, 0.44, or 2.67 MBq/μg for biodistribution studies and with 90Y at a specific activity of 65.5 MBq/μg for RIT (22).
Cell Lines
A431-K5 cell, a cell line established by transfection of a gene encoding mesothelin (13) to human epidermoid cancer A431 cells (American Type Culture Collection), was used as target in vitro and in vivo. A431-K5 has a stable and uniform expression of mesothelin (33,000 mesothelin sites per cell; Raffit Hassan, MD, unpublished data), which is similar to that observed with tumor specimens obtained from patients (13). A431-K5 and A431 cells were grown as previously described (12).
Binding Assays
Biotin-binding capacity of SS1scFvSA was determined as follows. First, SS1scFvSA was incubated with a molar excess of 111In-DOTA-biotin for 30 min. Then the mixture was applied to a gel-filtration size-exclusion column (Amersham Biosciences) to separate the bound and unbound fractions. The average number of biotins associated with the fusion protein was calculated.
The immunoreactive fraction (IRF) of both 125I- and 111In-SS1scFvSA was determined by a cell-binding assay using A431-K5 cells based on the method of Lindmo et al. (23). In addition, the IRF of 125I-SS1scFvSA was further examined by high-performance liquid chromatography (HPLC) analysis as follows. 125I-SS1scFvSA (18.48 ng) was incubated with a 400× molar excess of soluble mesothelin (16 μg; obtained from Dr. Ira Pastan’s laboratory at the National Institutes of Health [NIH]) for 1 h at room temperature and analyzed by HPLC using a Superose12 10/300 GL column (Amersham Biosciences). The amount in complexes was determined by comparing to the HPLC profile of the 125I-SS1scFvSA parental material with that mixed with excess of mesothelin (Peak Fit; Systat Software Inc.).
Competition assays were performed using a tracer amount of 125I-SS1scFvSA and competing with either unlabeled SS1scFvSA or a recombinant antimesothelin immunotoxin, SS1(dsFv)PE38, which has 1 Fv identical to those of SS1scFvSA and is known to specifically bind to mesothelin. One million A431-K5 cells were incubated with 5 ng of 125I-SS1scFvSA and increasing concentrations of unlabeled SS1scFvSA or SS1(dsFv)PE38 for 1 h at 4°C. The dose required to inhibit 50% of the binding of radiolabeled SS1scFvSA to mesothelin-expressing cells (IC50) was determined (Prism 4.0; GraphPad Software, Inc.).
Mouse Model
Female, 5-wk-old, athymic nude mice were inoculated subcutaneously with 3.8 × 106 A431-K5 cells on the left flank and, for the biodistribution studies, A431 cells were also inoculated on the right flank as a control. Experiments were performed when xenograft tumors reached a diameter of about 5 mm. Animal experiments were performed under an NIH Animal Care and Use Committee–approved protocol. Mice were euthanized when the tumor size reached >2 cm in diameter, tumor was ulcerated, or excessive weight loss was noted, according to the protocol guidelines.
Internalization of SS1scFvSA
125I- and 111In-SS1scFvSA were used to determine the rate of internalization by A431-K5 cells. In brief, A431-K5 cells (1 × 106) were incubated in round-bottom tubes with 125I- or 111In-SS1scFvSA (5 ng) for 1 h at 4°C. The cells were then centrifuged and washed once with phosphate-buffered saline (PBS) and collected for γ-counting (time 0) or further cultured at 4°C or 37°C in fresh medium. At various time points, the supernatant and the cells were separated by centrifugation. To determine the internalized fraction, the cells were then washed with 1 mL of an acidic buffer (0.1 mol/L sodium acetic acid in 0.01 mol/L PBS, pH 2.85) for 5 min and separated by centrifugation to determine the cell-bound (acid resistant, internalized fraction) and the released radioactivity (acid-labile, membrane-bound fraction). The released radioactivity was further separated into the protein-bound and the free radioactivity using trichloroacetic acid precipitation method (24).
Biodistribution of SS1scFvSA
The effect of the SS1scFvSA protein mass on biodistribution was studied using 100, 400, or 600 μg of SS1scFvSA at various times (14–48 h) after injection. A mixture of unlabeled SS1scFvSA and tracer doses of 125I- (0.20 μg/48.1 kBq) and 111In- (0.08 μg/33.3 kBq) SS1scFvSA was injected to mice bearing A431-K5 and A431 tumors.
Pretargeting Study
Our pretargeting protocol had 3 steps as follows: (i) We first injected SS1scFvSA, 24 h before injection of radiolabeled DOTA-biotin (−24 h); (ii) then, 20 h later, we injected synthetic clearing agent (sCA, 100 μg), consisting of a bifunctional moiety with multiple N-acetyl-galactosamine residues linked to biotin (8,651 Da) (6), to clear the blood of SS1scFvSA (−4 h); and (iii) 4 h later, the radiolabeled DOTA-biotin was administered (0 h).
The efficacy of sCA to clear the circulating SS1scFvSA was examined in 3 nude mice that were injected intravenously with a mixture of 125I- and 111In-SS1scFvSA (11.1 kBq/400 μg) followed by an injection of 100 μg of sCA 1.5 h later. Serial blood samples were collected from the tail vein immediately before (time = 0) and 3 min to 2 h after the injection of sCA. The samples were counted in a γ-counter (Wallac Inc.), and the percentage injected dose per gram (% ID/g) in the blood was calculated.
Biodistribution of pretargeted 111In DOTA-biotin was determined at various times after injection. In addition, the effect of the mass (i.e., specific activity) of the DOTA-biotin on the tumor uptake was examined in a dose range of 0.1–5 μg DOTA-biotin.
For pretargeting studies, mice were fed a biotin-free diet starting 7 d before the SS1scFvSA injection to reduce the endogenous biotin level (biotin-deficient Purina diet). A regular diet was then started 2 d after injection of the radiolabeled DOTA-biotin.
Dosimetry
For dosimetry estimates, 88Y-DOTA-biotin was used in place of 90Y-DOTA-biotin. Together with 88Y-DOTA-biotin, 177Lu-DOTA-biotin was also evaluated by a dual-isotope method. In brief, a mixture of 88Y-DOTA-biotin (111 kBq/253 ng) and 177Lu-DOTA-biotin (148 kBq/55.5 ng), to which unlabeled DOTA-biotin was added to adjust the total DOTA-biotin dose to 0.5 μg, was injected to groups of mice that had received injections of SS1scFvSA (600 μg) and sCA following our pretargeting protocol. Biodistribution was determined after 2, 24, 48, and 120 h. The mean %ID/g of 88Y- DOTA-biotin in the A431-K5 tumor and tissues from 2 h to 5 d was determined. The integral of the radioactivity underneath the curve (AUC) in the tumor and selected organs was then extrapolated to that of 90Y accounting for the decay differences using trapezoidal integration up to 5 d and estimating the area beyond 5 d assuming it cleared with the estimated terminal half-life of the clearance curve. 90Y radiation dosimetry was then calculated using the AUC determined for 90Y and the medical internal radiation dose method adjusted for organ size using the “Nodule Module” of MIRDOSE3 (25). The dosimetry for 177Lu DOTA-biotin was estimated by determining the AUC for 177Lu and using the dose factors for 177Lu obtained online at www.doseinfo-radar.com derived using methodology described by Stabin and Siegel (26). The dose was estimated for 37 MBq DOTA-biotin of each isotope, and the dose factors used were adjusted to tumor or organ weight.
Therapy Study
Pretargeted RIT was performed in groups of mice bearing A431-K5 tumors. Mice received intravenous injections of 600 μg of SS1scFvSA (at −24 h) and sCA (at −4 h) and were then injected with 11.1, 20.4, or 32.4 MBq/0.5 μg of 90Y labeled DOTA-biotin. As control animals, we used one group of mice without any treatment and another treated with 32.4 MBq of 90Y DOTA-biotin after pretargeting with the nonspecific anti-Tac scFvSA. The sizes of A431-K5 tumors were measured once or twice a week after the therapy and the tumor volume was calculated using the formula (width2 × length)/2. Survival of the mice was followed daily. Toxicity was assessed by following serial complete blood counts, observation of mouse activity, and body weight.
Statistical Analysis
Statistical analysis was performed using a nonpaired t test for comparison of 2 groups, ANOVA for comparison of 3 or more groups, or Kaplan–Meier cumulative survival plots (StatView version 5.0, SAS Institute Inc.).
RESULTS
Characterization of SS1scFvSA
The SS1scFvSA molecule is a tetrameric fusion protein with 4 binding sites for biotin and 4 Fv Ag-binding sites. Each SS1scFvSA molecule was estimated to be capable of binding an average of 2.8 111In DOTA-biotin molecules. The IRFs of 125I- and 111In- SS1scFvSA based on cell binding were 50.3% ± 4.6% and 50.1% ± 3.5% (average ± SD), respectively. SS1scFvSA did not bind to the parental cell line, A431, which does not express mesothelin. To exclude the possibility of having damaged SS1scFvSA binding to biotin during the radiolabeling procedure, we examined the IRF of SS1scFvSA-111In-DOTA-biotin, by reacting SS1scFvSA with 111In-DOTA-biotin, followed by purification using a size-exclusion column. The IRF of SS1scFvSA-111In-DOTA-biotin was 47.6% and comparable to that of 125I- and 111In-SS1scFvSA. The IRF of 125I-SS1scFvSA determined by the HPLC assay with soluble mesothelin was 64.5%.
Competition assays were performed to examine the binding specificity and avidity of SS1scFvSA to mesothelin using unlabeled SS1scFvSA and a mesothelin-specific immunotoxin, SS1(dsFv)PE38. The IC50 determined by competition assay was 2 nmol/L for SS1scFvSA, whereas that of SS1(dsFv)PE38 was 23 nmol/L, indicating that SS1scFvSA was specifically bound to mesothelin expressed on the cells and that SS1scFvSA with 4 Fv sites has a >11-fold higher avidity to mesothelin compared with SS1(dsFv)PE38 with 1 Fv.
Internalization of SS1scFvSA
In the pretargeting approach, only the SS1scFvSA on the target cell surface contributes to the binding of the radiolabeled DOTA-biotin. We examined the rate of internalization of 125I- and 111In-SS1scFvSA from the cell surface using A431-K5 cells. As shown in Figure 1, approximately 40% of total radiolabeled SS1scFvSA bound to the cells after a 1-h preincubation at 4°C (time 0) and more than half of this initially bound 111In- or 125I-SS1scFvSA was internalized by 3 h at 37°C. the internalized fraction (acid resistant) of 111In-SS1scFvSA increased to >60% of initially bound SS1scFvSA at 18 h (Fig. 1A), whereas the acid-resistant fraction of 125I-SS1scFvSA decreased after 3 h and the free (nonprotein bound) 125I in the medium increased (Fig. 1B). This indicates that internalized 125I-SS1scFvSA underwent dehalogenation and the free iodine was released into the medium. Internalization of prebound 111In- and 125I-SS1scFvSA was blocked at 4°C (data not shown).
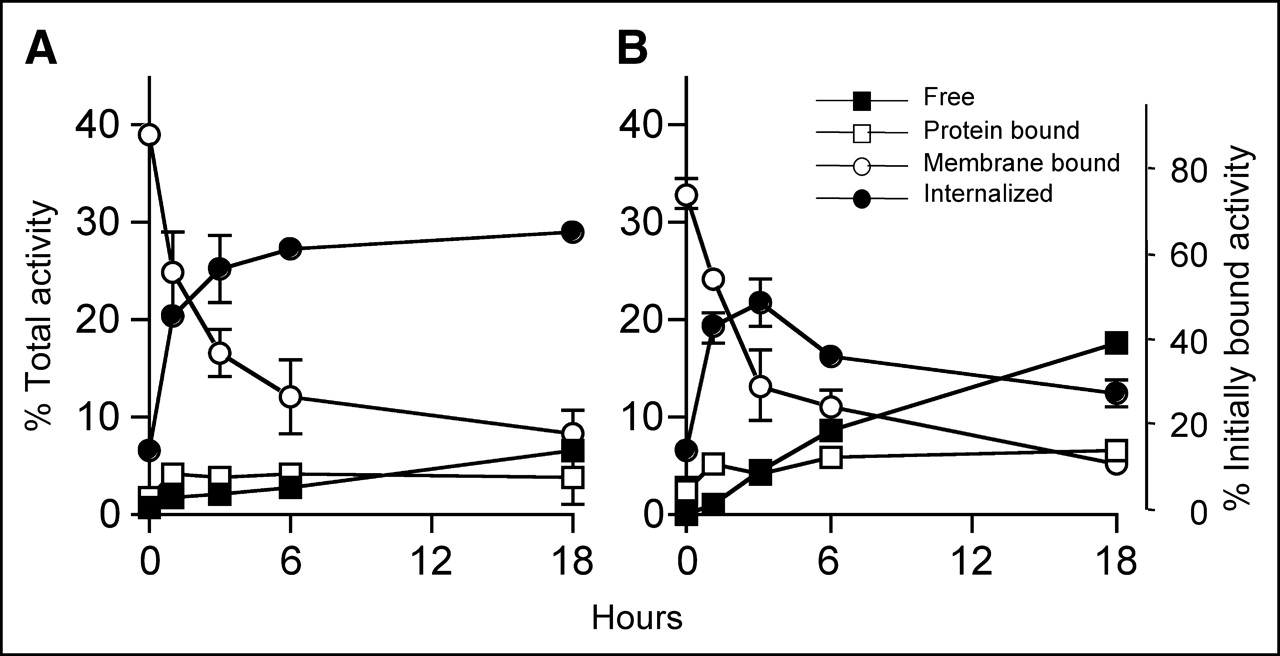
Rate of internalization of 111In- (A) and 125I- (B) SS1scFvSA bound to A431-K5 cells was evaluated. Cells were preincubated with radiolabeled fusion proteins for 1 h at 4°C and then were transferred into fresh medium and cultured at 37°C (A and B) or 4°C (data not shown). At various time points, radioactivity bound to cells was separated into the internalized (acid resistant) fraction and membrane-bound (acid labile) fraction by using an acid wash, and radioactivity in supernatant was separated into protein-bound and free (nonprotein bound) radioactivity using trichloroacetic acid. SS1scFvSA was highly internalized by A431-K5 cells at 37°C.
Clearance of Circulating SS1scFvSA by sCA
In our multistep RIT, sCA was used to remove the circulating fusion protein to avoid binding of the radiolabeled biotin the circulating fusion protein, which would lead to increased normal tissue irradiation. The sCA injection resulted in rapid removal of the 111In- and 125I-SS1scFvSA from the blood. The level of 111In- and 125I-SS1scFvSA in the blood before sCA was 31.7 and 43.4 %ID/g and decreased to 1.5 and 4.5 %ID/g at 2 h after sCA, with half-lives of 25 and 38 min, respectively. The half-life of SS1scFvSA without injection of sCA calculated from the biodistribution studies (vide infra) was 8.85 h.
Optimization of SS1scFvSA Dose
To optimize scheduling and dosing of SS1scFvSA in pretargeting, the biodistribution of 125I- and 111In-SS1scFvSA was examined at different time points and doses. 111In-SS1scFvSA (600 μg) showed the highest A431-K5 tumor uptake of 14.8 %ID/g at 24 h (Fig. 2A), whereas the 125I-SS1scFvSA accumulation in A431-K5 tumor peaked at 8.2 %ID/g at 14 h and decreased thereafter (P < 0.01 for 14 vs. 24 h and 24 vs. 48 h, P < 0.0001 for 14 vs. 48 h, Fig. 2B). The decrease of the 125I- SS1scFvSA tumor uptake and the increase in gastric uptake were consistent with dehalogenation after internalization (Fig. 2B). High liver and splenic uptakes were observed with 111In-SS1scFvSA (16.0 and 17.7 %ID/g at 24 h, respectively). Examination for the formation of aggregates in the 111In-SS1scFvSA sample by HPLC revealed no presence of aggregates, but revealed a major peak of 111In-SS1scFvSA (>90%) and a minor peak consistent with a dimer of the SS1scFvSA, which was also present in the unlabeled SS1scFvSA (data not shown). Thus, the reason for the high liver and spleen uptakes was unknown.
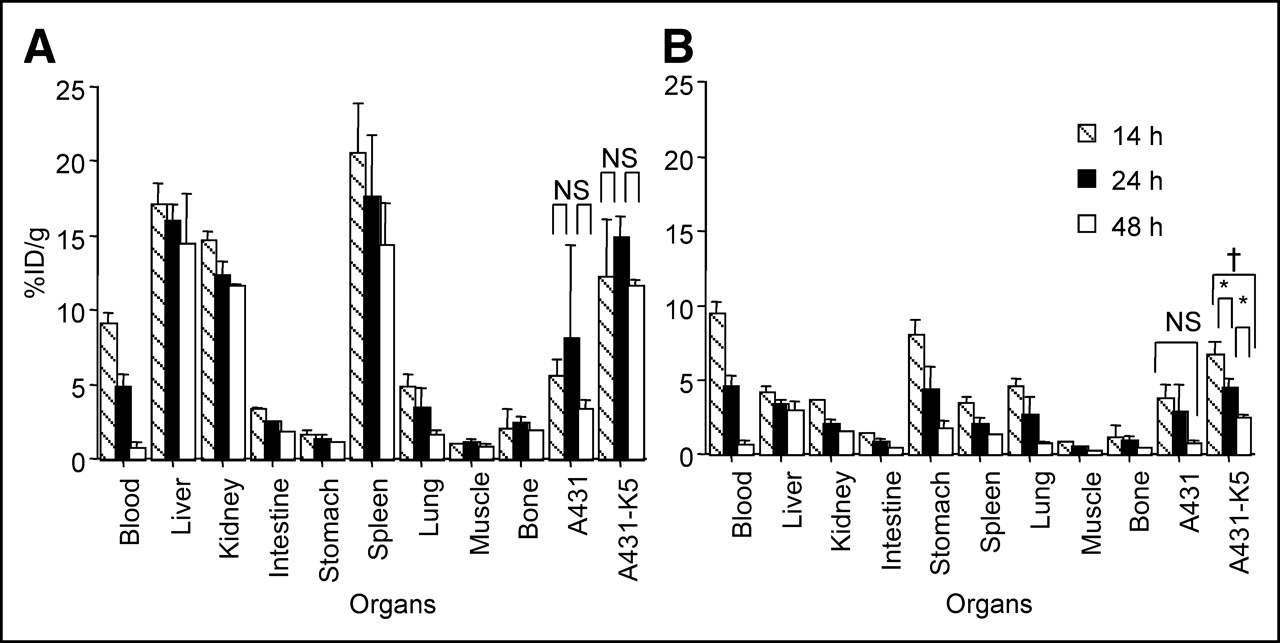
Biodistribution of 111In- (A) and 125I- (B) SS1scFvSA (600 μg) 14, 24, and 48 h after injection (n = 5; mean %ID/g ± SD, *P < 0.01, †P < 0.0001, NS: P > 0.05). NS = not significant.
Although increasing the dose of SS1scFvSA from 100 to 600 μg resulted in a lower %ID/g in A431-K5 cells (P < 0.05 for 100 vs. 600 μg), the total mass of SS1scFvSA taken by A431-K5 tumor increased (P < 0.0001 for 100 vs. 400 or 600 μg; Figs. 3A–3C). When the specific tumor uptake was calculated by subtracting the 111In-SS1scFvSA uptake in A431 tumor uptake (μg/g) from A431-K5 tumor uptake (μg/g), it appeared that the A431-K5 tumor was approaching saturation between 400 and 600 μg of SS1scFvSA (Fig. 3C) (P > 0.5 for 400 vs. 600 μg). We then evaluated the biodistribution of DOTA-biotin (1 μg) after pretargeting using 400 and 600 μg of SS1scFvSA. The 600 μg of SS1scFvSA gave a >2-fold higher A431-K5 tumor uptake than that achieved with 400 μg at 24 h (P < 0.01). The uptake of 111In-DOTA-biotin in A431 tumor and spleen was also increased (P < 0.05), but other normal organ uptake did not change by increasing the dose of SS1scFvSA.
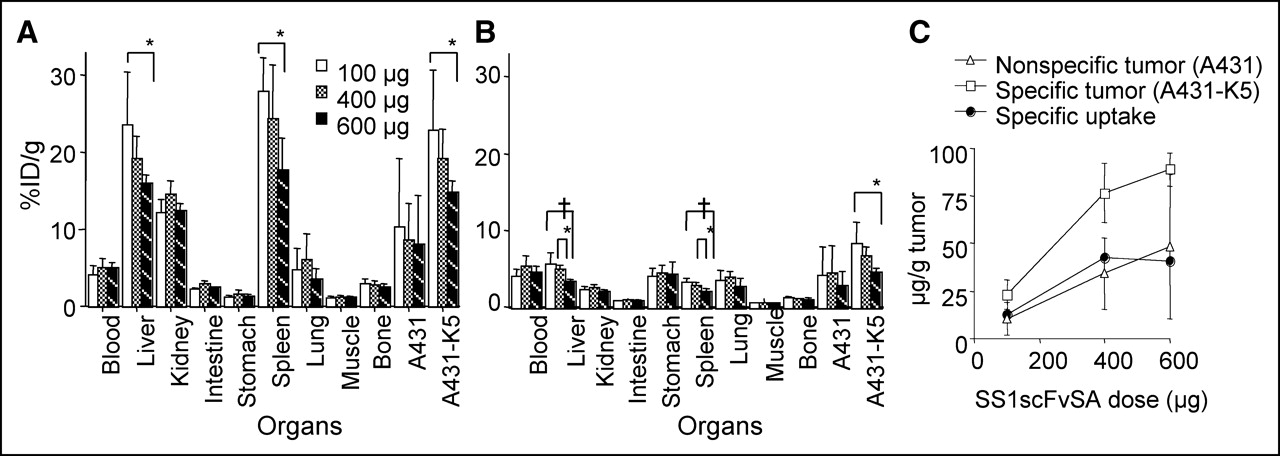
Biodistribution of 111In- (A) and 125I- (B) SS1scFvSA (100, 400, and 600 μg) 24 h after injection was determined (n = 5; mean %ID/g ± SD, *P < 0.05, †P < 0.01). The absolute amount of SS1scFvSA taken up by specific tumor was calculated from biodistribution data of 111In-SS1scFvSA; SS1scFvSA mass taken up per gram of nonspecific tumor was subtracted from A431-K5 tumor uptake (μg/g). A431-K5 tumor was approaching saturation between 400 and 600 μg of SS1scFvSA (C).
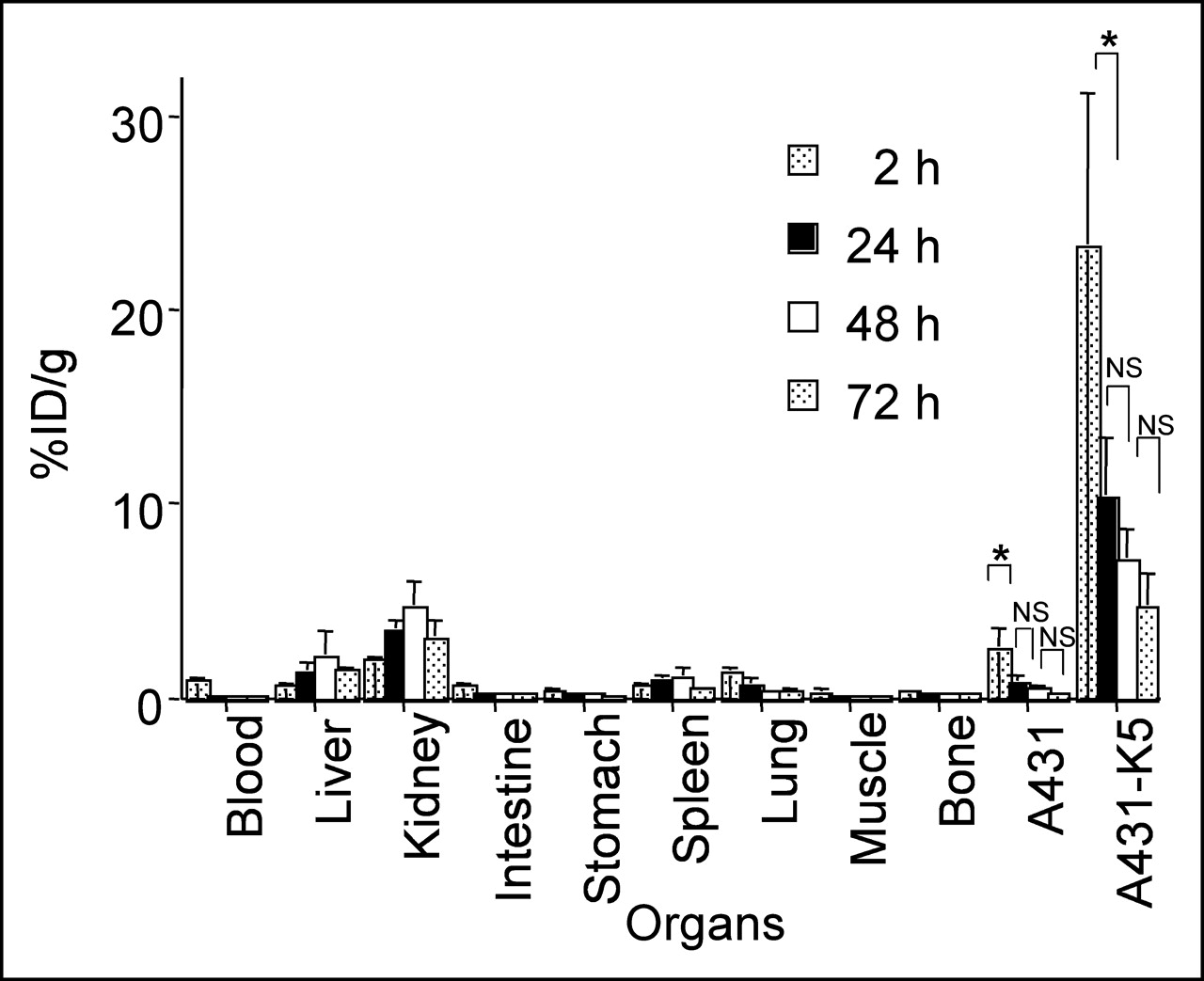
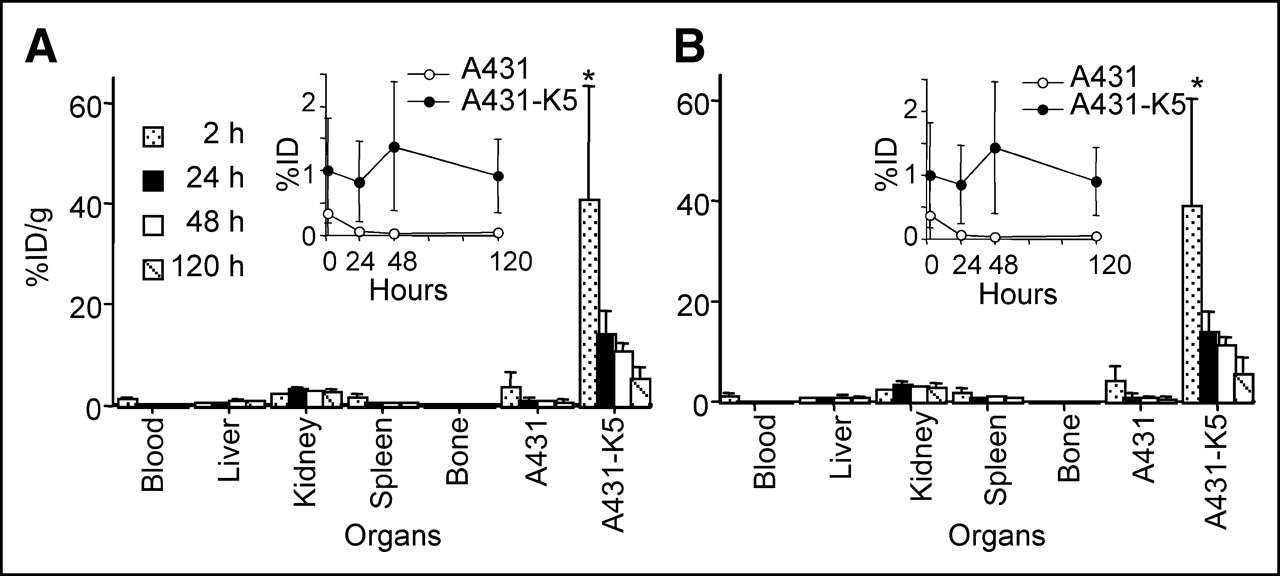
111In-DOTA-biotin (1 μg), injected after pretargeting with 600 μg of SS1scFvSA and sCA, rapidly accumulated in A431-K5 tumor with >20 %ID/g at 2 h, which decreased thereafter due to an increase in tumor volume. Normal organ uptake was <3.5 %ID/g (Fig. 4).

With 600 μg of SS1scFvSA, 111In-DOTA-biotin (1 μg) rapidly accumulated in A431-K5 tumor with >20 %ID/g at 2 h after pretargeting and sCA injection (n = 5–12). Tumor uptake decreased thereafter, which was predominantly accounted by tumor growth since there was no decrease in total uptake dose in tumor (mean %ID/g ± SD, *P < 0.0001, NS: P > 0.05). NS = not significant.
Optimization of DOTA-Biotin Dose
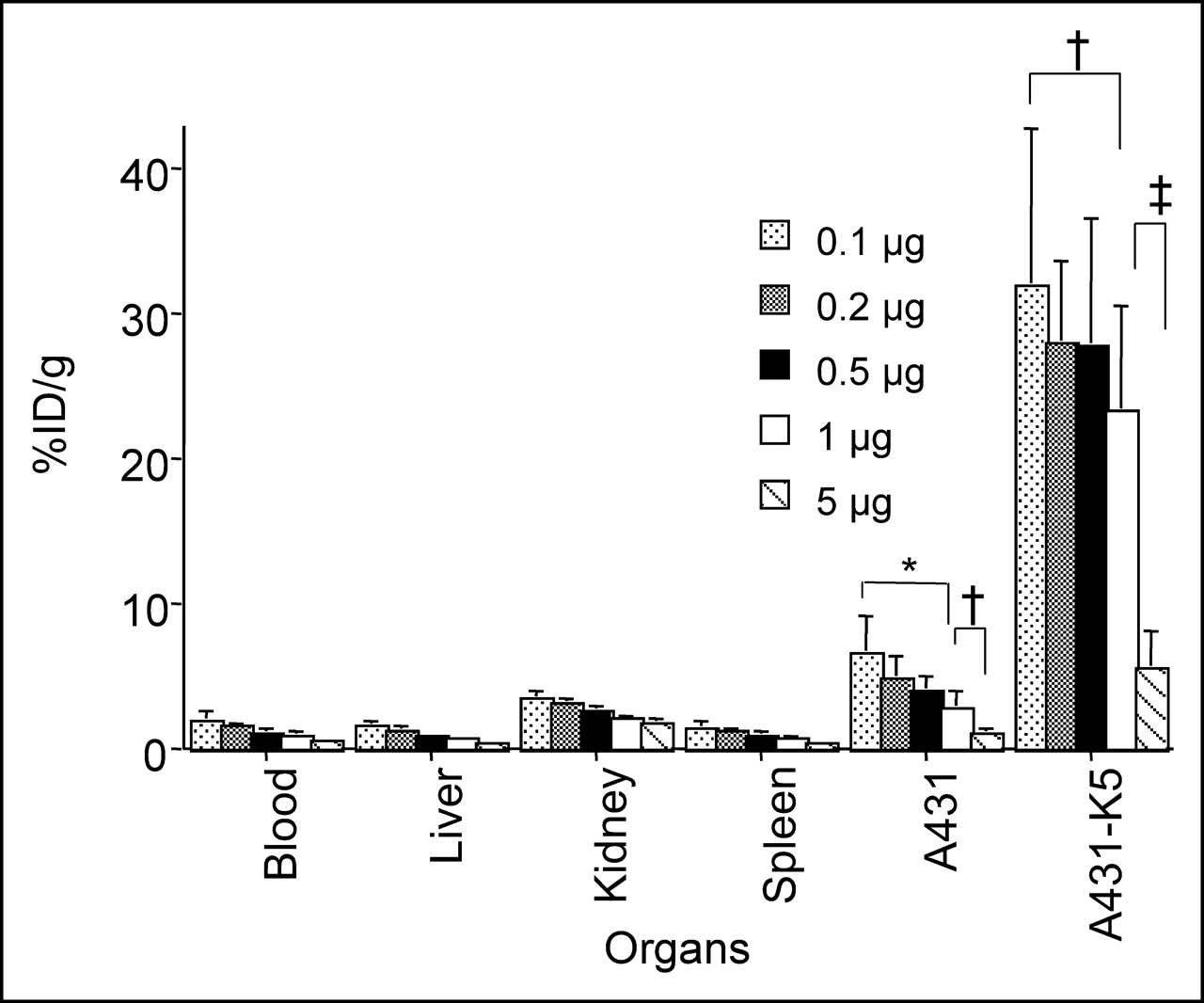
Various experiments were performed to determine the effect of the specific activity of radiolabeled DOTA-biotin on tumor uptake. The tumor uptake of radiolabeled DOTA-biotin at doses of 0.1–5 μg was evaluated 2 h after its administration following the initial 2 pretargeting steps. The A431-K5 tumor uptake decreased as the mass of DOTA-biotin increased, suggesting a saturable uptake (Fig. 5). With 0.5, 0.2, or 0.1 μg of radiolabeled DOTA-biotin, the tumor uptake was >27 %ID/g, whereas with 1 μg, the uptake decreased to 23.3 %ID/g. Five micrograms of DOTA-biotin resulted in only 5.5 %ID/g in A431-K5 tumor (P < 0.05 for 0.1 vs. 1 μg; P < 0.0001 for 0.1, 0.2, 0.5, or 1 μg vs. 5 μg). A comparison of the 111In-DOTA-biotin uptake in the nonspecific tumor and various normal tissues also showed a saturable effect.
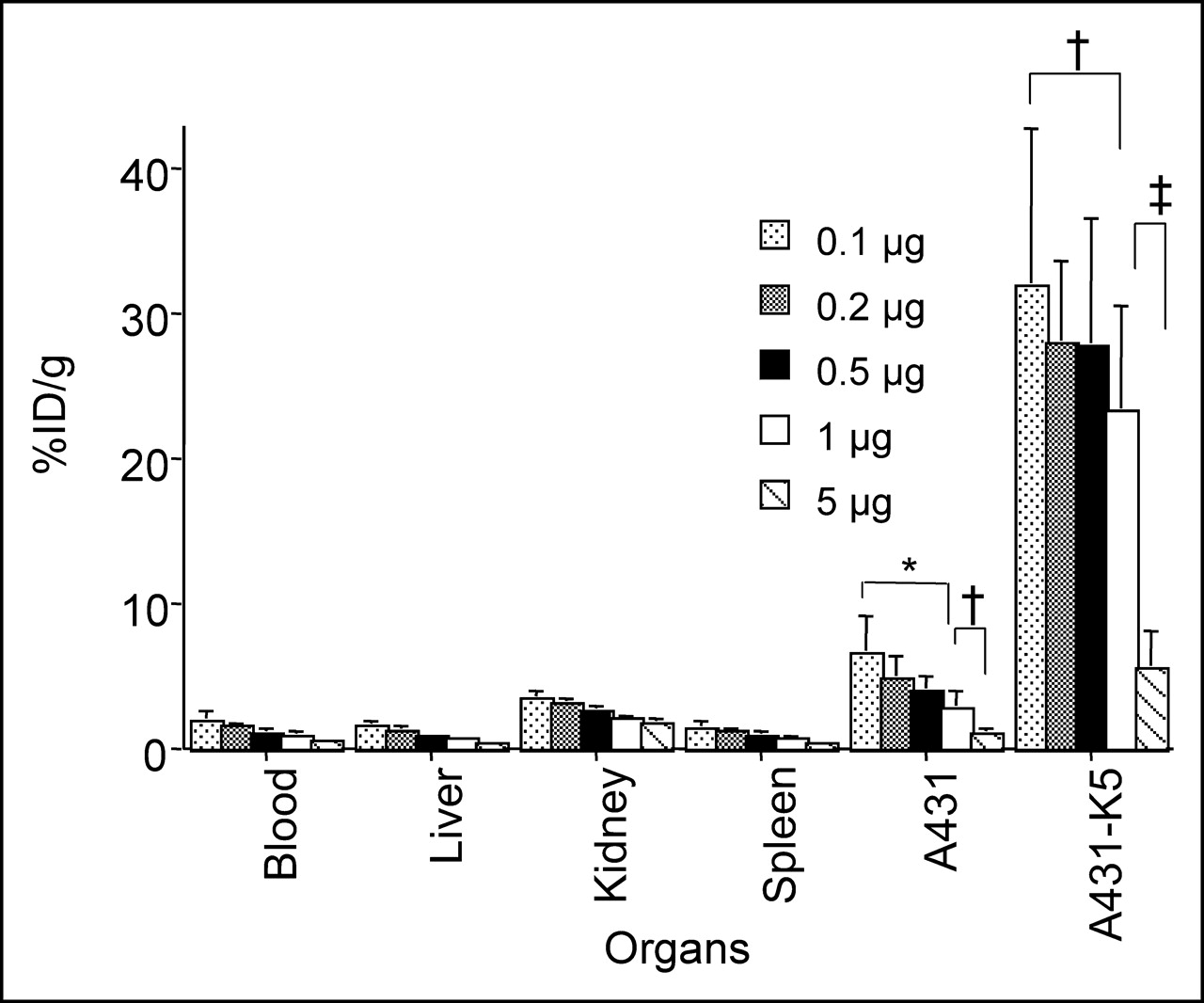
Effect of dose (0.1–5 μg) of 111In-DOTA-biotin on tumor uptake was evaluated. Biodistribution of 0.1–5 μg of 111In-DOTA-biotin after pretargeting with SS1scFvSA and sCA injection was examined at 2 h. A431-K5 tumor uptake decreased as mass of DOTA-biotin increased, suggesting a saturable uptake (n = 7–10; mean %ID/g ± SD, *P < 0.01, †P < 0.05, ‡P < 0.0001).
Therapy
The radiation dosimetry of 90Y-DOTA-biotin was calculated from the time–activity curves obtained from 88Y-DOTA-biotin (%ID/g). The biodistribution of pretargeted 177Lu-DOTA-biotin was evaluated as described for 111In and 88Y. The biodistributions of 88Y- and 177Lu-DOTA-biotin (Figs. 6A and 6B) were almost identical with no significant difference in A431-K5 or nonspecific tumor uptake observed at any time points (P > 0.05). The uptake to A431-K5 tumor at 2 h was comparable to that of 0.5 μg of 111In-DOTA-biotin shown in Figure 6 (P > 0.05). The decrease of %ID/g in A431-K5 tumor was mainly due to the growth of the tumor during the dosimetry study, because absolute A431-K5 tumor uptake (%ID ± SD) did not change (insets in Figs. 6A and 6B).
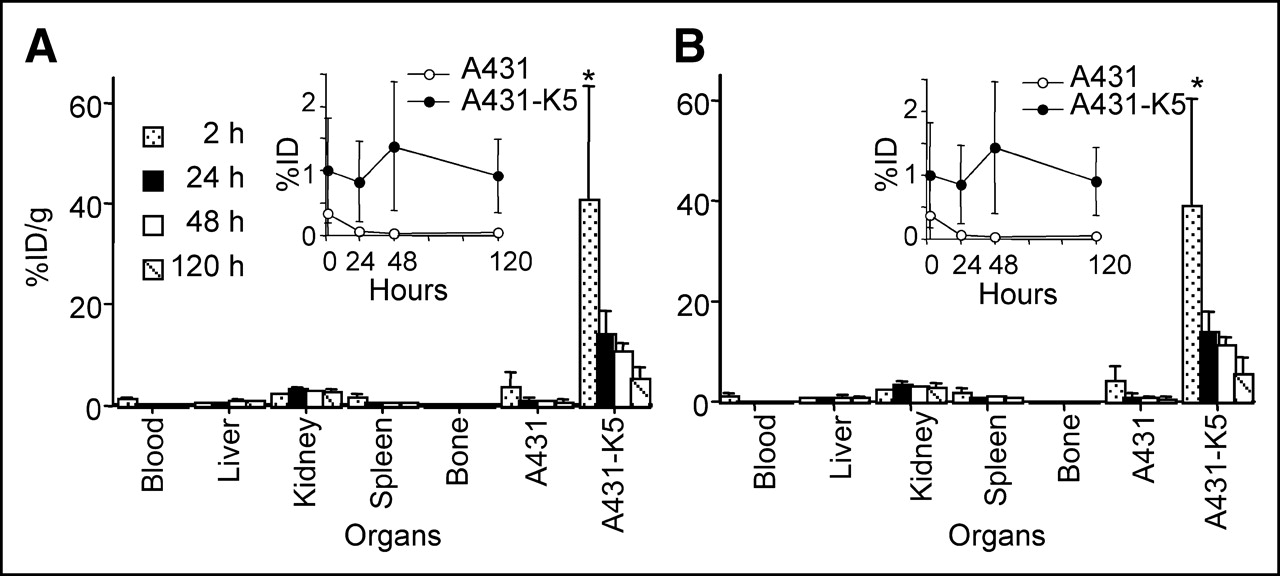
Biodistribution of 88Y- (A) and 177Lu- (B) DOTA-biotin after pretargeting with 600 μg of SS1scFvSA and administration of sCA. 88Y- and 177Lu-DOTA-biotin showed a similar biodistribution at 2–120 h after injection (n = 5; mean %ID/g ± SD, *P < 0.05). Absolute A431-K5 tumor uptake is shown as insets (mean %ID ± SD) As A431-K5 tumor grew, absolute tumor A431-K5 uptake was unchanged.
Pretargeted RIT with 90Y-DOTA biotin led to shrinkage of A431-K5 tumor and resulted in a dose-dependent survival rate of the mice (Fig. 7). With 11.1, 20.4, and 32.4 MBq of 90Y-DOTA-biotin, 57% (4/7), 71% (5/7), and 86% (6/7) of mice were alive 110 d after the RIT, respectively (P < 0.01 0 vs. 11.1, 20.4, or 32.4 MBq; P > 0.05 for 11.1 vs. 20.4 MBq and 20.4 vs. 32.4 MBq). Since, in 2 of 4 surviving mice treated with 11.1 MBq, the tumor regrew to 4–5 mm in diameter at day 110, the tumor-free survival rate of the 11.1-MBq group was 29%. All of the surviving mice treated with 20.4 or 32.4 MBq had no tumor recurrence up to 110 d. All of the nontreated mice died, with a median survival of 16 d. As a control, we used a nonspecific fusion protein, anti-Tac scFvSA, as a “pretargeting” molecule and used a 90Y-DOTA-biotin radioactivity dose of 32.4 MBq. With this high dose, the growth of the tumor was somehow retarded, with a median survival of 43 d (Fig. 7). Control experiments were repeated using groups of mice (n = 5) treated with 90Y-DOTA-biotin pretargeted by anti-Tac scFvSA or 90Y-DOTA-biotin alone, or with no treatment. The use of anti-Tac scFvSA as a pretargeting molecule resulted in similar findings. 90Y-DOTA-biotin alone without pretargeting had no effect in prolonging survival compared with the nontreated group.
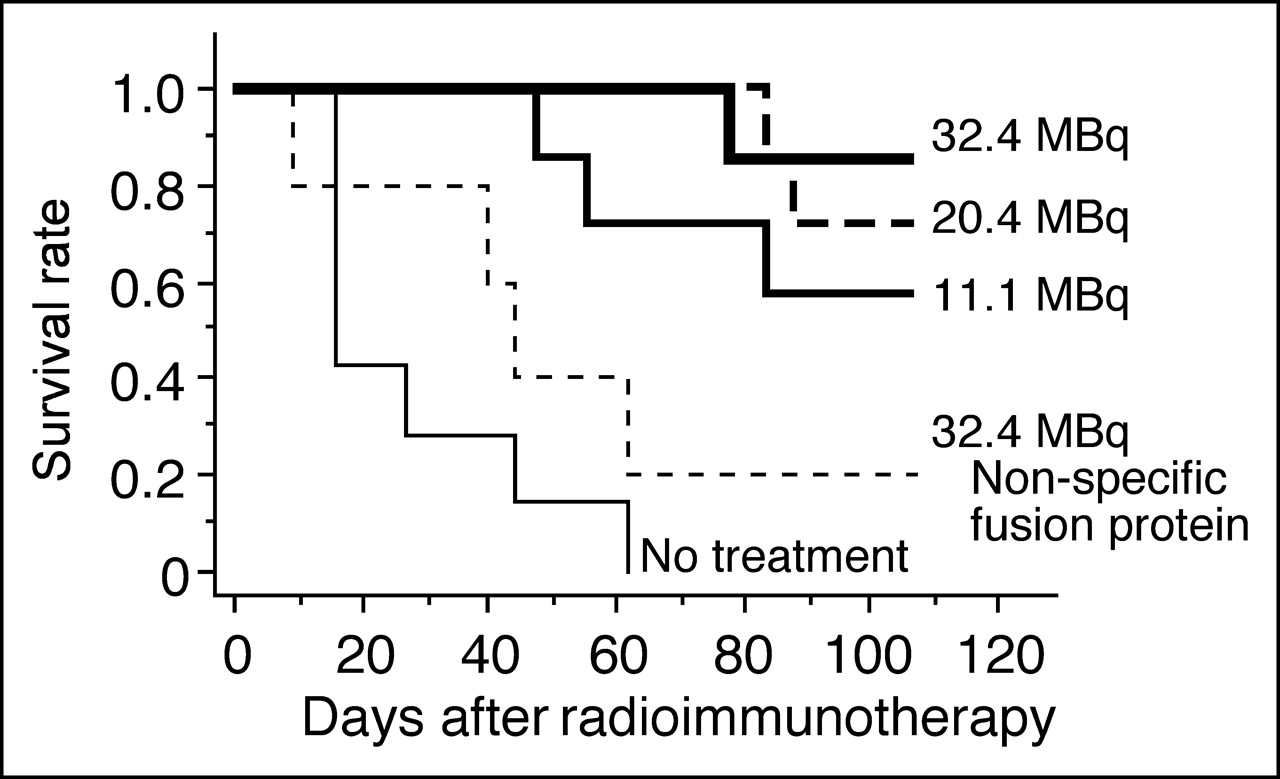
Kaplan–Meier survival plot. Survival of A431-K5 tumor-bearing mice after RIT was dose dependent. With 11.1, 20.4, and 32.4 MBq of 90Y-DOTA-biotin, 57% (4/7), 71% (5/7), and 86% (6/7) of mice were alive 110 d after RIT, respectively. A high dose of 90Y-DOTA-biotin (32.4 MBq) pretargeted with nonspecific anti-Tac scFvSA had some effect, but survival was not significantly prolonged compared with that of nontreated group.
The dosimetry estimates for the specific A431-K5 tumor and the other organs after injection of 37 MBq of 90Y- or 177Lu-DOTA-biotin are shown in Table 1. In our pretargeted RIT, 90Y doses of 11.1, 20.4, and 32.4 MBq gave estimated tumor irradiation doses of 19, 36, and 57 Gy, respectively. Normal organ and nonspecific tumor irradiation doses were estimated to be <10 Gy, except for the kidney, which received 29.7 Gy.
Estimated Dose Delivered to Organs and Tumors by Pretargeted 90Y- or 177Lu-DOTA-Biotin: Gy/37 MBq (1 mCi) Injection*
Toxicity
A transient decrease of white blood cell (WBC), red blood cell (RBC), and platelet counts was observed. The WBC counts at the nadir (day 14) in mice receiving 90Y doses of 11.1, 20.4, and 32.4 MBq were 26%, 17%, and 10% of that of nonirradiated mice, respectively. In the group treated with 32.4 MBq, it took >4 wk to recover the WBC counts to baseline. The RBC counts decreased to 88%, 73%, and 71% and the platelet counts decreased to 73%, 41%, and 39%, respectively, with a nadir between day 9 and day 14. Both RBC and platelet counts recovered by day 22. A decrease in body weight was not observed in any group after the therapy.
DISCUSSION
In this study, we investigated pretargeting RIT of mesothelin-expressing tumor using SS1scFvSA, a tetravalent fusion protein. Mesothelin is a differentiation Ag expressed on several human malignancies and normal mesothelial cells. Because of its limited expression in normal organs, mesothelin may be a good target for antibody-based therapies. As with other antibodies, with some crossreactivity with normal tissues, careful dose-escalation studies need to be performed to exclude normal tissue toxicity. Pretargeted RIT has an advantage over immunotoxins, which inhibit protein synthesis specifically limited to the cell that internalizes it, whereas the irradiation from radiolabeled biotin bound to the SS1scFvSA on the tumor cell surface can kill several tumor cells within its effective pathlength. One disadvantage of this approach is the high immunogenicity of SA, as with all pretargeting approaches using the SA-biotin system, which may allow only one injection of this reagent (11). Although some approaches are being explored to decrease the immune response to SA, other pretargeting approaches are expected to have less immunogenicity.
As shown in this article and by others, genetically engineered tetravalent fusion proteins have biochemical uniformity and, in addition, are easier to produce (6,18), which make them more desirable agents compared with chemically synthesized mAb-SA covalent conjugates. In addition, because they lack the Fc region, fusion proteins have shown a more rapid blood clearance than mAb-SA conjugates (5,6,18,27). At the same time, because of their greater molecular weights (>170 kDa), fusion proteins have longer half-lives in the vascular compartment than conventional antibody fragments, which are readily cleared via glomerular filtration.
When we compared the affinity of SS1scFvSA with its 4 Fv Ag-binding sites to that of the unlabeled SS1(dsFv)PE immunotoxin, which has only 1 Fv, there was an 11.5× lower IC50, consistent with a higher avidity. Similar findings have been described previously with other fusion proteins (6,18). The biotin-binding capacity of the SS1scFvSA molecule (∼2.8 biotins) was lower than the theoretic maximum of 4, but was in the range of other tetravalent fusion proteins (6,18). This lower value could be the result of some occupancy of the SA by biotin in the media during the generation of the fusion protein.
The biodistributions of SS1scFvSA radiolabeled with 111In and 125I were significantly different. These findings are likely related to dehalogenation and subsequent release of 125I, which is common with many internalizing antibodies (28). Similar findings have been observed with 111In- versus 125I-labeled humanized anti-Tac, which undergoes slow internalization (Meili Zhang, PhD, personal communication, 2002). It should be noted that internalization of mAb targeting the same Ag may be cell line dependent, as shown recently (7). 111In-SS1scFvSA showed high uptakes in the liver and spleen (Fig. 3A). Though this could be detrimental if we were using a directly labeled antibody, this was not problematic for pretargeting since the antibody in these organs was not available for binding of the radiolabeled DOTA-biotin (Fig. 5). Since we did not observe such a high uptake in the liver or spleen with 125I-SS1scFvSA, the uptake seen with 111In-SS1scFvSA could be due to the labeling procedure with 111In. Biodistribution of 111In-labeled K1 antibody against mesothelin showed a slightly higher A431-K5 tumor uptake (∼30 %ID/g) and lower splenic and liver uptakes (<10 %ID/g at 24 h) than those shown by 111In-SS1scFvSA (12). These differences in tumor accumulation may be related to the shorter retention in the blood for the fusion proteins that has been observed in other model systems (18). Similarly, slightly higher uptake has been seen in the liver, spleen, and kidney when 125I-labeled huNR-Lu-10 scFvSA was compared with the chemical conjugated counterpart (18). Although the directly labeled antibody conjugate showed localization in the liver, spleen, and kidney, this was not reflected in the biodistribution of pretargeted radiolabeled DOTA-biotin, which showed a favorable localization in tumor and high tumor-to-nontumor ratios.
This study showed that the mass of DOTA-biotin injected follows saturable ligand–receptor type interaction. By decreasing the mass (i.e., increasing the specific activity) of DOTA-biotin injected per mouse, we could successfully obtain a significantly higher A431-K5 tumor uptake of radiolabeled biotin. Many investigators, including us, used 1 μg of biotin in multistep tumor targeting of mice (10,22). In our study, 1 μg of DOTA-biotin gave >4 times higher tumor uptake than that shown with 5 μg. When we further lowered the injected DOTA-biotin amount, the tumor uptake continued to increase (Fig. 5). A similar finding has been reported in a separate system using NR-Lu-10 mAb-SA. The degree of saturation by DOTA-biotin will depend on the amount of mAb-SA available on the tumor surface. The issue of specific activity is significant for isotopes that result in labeling of DOTA-biotin at low specific activity due to their labeling kinetics, low specific activity, or contaminants, such as occurred for the 177Lu that was available at the time of our study. With this 177Lu, the highest specific activity we could obtain would allow us to inject only 1.33 MBq of radiolabeled DOTA-biotin on 0.5 μg. With technical improvements, higher specific activity 177Lu is now available.
In the pretargeted RIT, we decided to use 0.5 μg of DOTA-biotin per mouse, because it gave a high tumor uptake (>27 %ID/g) and also fulfilled the requirement to label with high enough specific activity that would allow us to inject as high as 32.4 MBq 90Y-DOTA-biotin per mouse. The high tumor-to-normal organ ratios resulted in cure of mesothelin-expressing tumor in 86% of mice when treated with an 90Y dose of 32.4 MBq. The survival of the mice was dose dependent and, even with 11.1 MBq, 29% of the mice were cured with a follow-up of 110 d. This high therapeutic effect of pretargeted RIT with SS1scFvSA is promising as a therapy for mesotheliomas, ovarian cancers, or other mesothelin-expressing malignancies. Although a side effect on mesothelin-positive normal tissue such as pleura or peritoneum may occur, this would have to be carefully monitored in the phase I trial setting. Other therapeutic successes have been seen in various mouse tumor models with a variety of antibody SA constructs and various isotopes (5,7,8,22,27).
The mice treated with 32.4 MBq of 90Y-DOTA-biotin without pretargeting did not show any treatment effect on their survival nor was there shrinkage of tumor when nonradiolabeled SS1scFvSA was administered (data not shown). A nonspecific therapeutic effect was seen on A431-K5 tumor when 32.4 MBq of 90Y-DOTA-biotin pretargeted with the nonspecific anti-Tac scFvSA. Similar nonspecific effects have been noted previously (22,29). This was likely secondary to the slightly longer circulation time of 90Y-DOTA-biotin when a pretargeting antibody is administered and to a certain amount of low-level nonspecific uptake. These preclinical studies with SS1scFvSA together with clinical studies performed previously with various SA chemical conjugates or fusion proteins (11,18,30) suggest that this is an approach that warrants further clinical investigation in mesothelin-expressing tumors. The dosimetry data (Table 1) showed that, among the normal organs, the kidneys received the highest dose; however, it was below the dose reported to cause toxicity (31). Behr et al. had reported that mice receiving a renal dose of <66 Gy had no evidence of renal toxicity and a renal dose of 72 Gy did not cause toxicity in our previous pretargeting studies using 90Y-DOTA-biotin (22,31). With our highest dose of 32.4 MBq, the dose delivered to the kidney was estimated to be 26 Gy.
In addition to demonstrating tumor targeting with yttrium-labeled DOTA-biotin after pretargeting with SS1scFvSA, to our knowledge, we are among the first to demonstrate that 177Lu- DOTA-biotin has a similar biodistribution as pretargeted 111In- and 88Y-DOTA-biotin. 177Lu has some features that may make it a therapeutic alternative to 90Y in certain situations, as it emits γ-rays that can be imaged, it emits a lower energy and, thus, shorter pathlength β-particle, and is easy to radiolabel using conventional chelates. We did not perform a therapy study using 177Lu, because of the unavailability of 177Lu with a high purity that could be labeled with DOTA-biotin at a high enough specific activity for therapy. Recently, higher specific activity 177Lu is commercially available, which would allow for higher doses for preclinical RIT studies.
CONCLUSION
SS1scFvSA, though showing some evidence of internalization, showed good pretargeting ability. Good tumor-to-background ratios were obtained when using 3 different radiolabeled (111In, 88Y, or 177Lu) DOTA-biotins. Furthermore, we demonstrated that the dose of DOTA-biotin is critical in optimizing pretargeted delivery. Using 90Y- DOTA-biotin, encouraging tumor responses were observed without reaching dose-limiting toxicity and, thus, this may be a promising approach for RIT of mesothelin-expressing malignancies.
Acknowledgments
We thank Drs. Thomas Waldmann and Meili Zhang for the anti-Tac scFvSA. research support: Fusion Protein, DOTA-biotin, and sCA were provided by NeoRx Corp.
Footnotes
Received Dec. 14, 2004; revision accepted Mar. 29, 2005.
For correspondence or reprints contact: Jorge A. Carrasquillo, MD, Nuclear Medicine Department, Warren G. Magnuson Clinical Center, 10 Center Dr., MSC-1180, Bethesda, MD 20892-1180.
E-mail: jcarrasquillo{at}cc.nih.gov
REFERENCES
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Pretargeting: A Path Forward for Radioimmunotherapy
- 124I-huA33 Antibody Uptake Is Driven by A33 Antigen Concentration in Tissues from Colorectal Cancer Patients Imaged by Immuno-PET
- A Fiber-Modified Mesothelin Promoter-Based Conditionally Replicating Adenovirus for Treatment of Ovarian Cancer
- Synergistic Antitumor Activity of Taxol and Immunotoxin SS1P in Tumor-Bearing Mice
- Comparison of a tetravalent single-chain antibody-streptavidin fusion protein and an antibody-streptavidin chemical conjugate for pretargeted anti-CD20 radioimmunotherapy of B-cell lymphomas
- Antibody Pretargeting Advances Cancer Radioimmunodetection and Radioimmunotherapy